Содержание
Введение
1 Факторы, определяющие реакцию растений на облучение
2 Действие внешнего ионизирующего излучения на организм
2.1 Варианты возможного радиационного воздействия
2.2 Влияние ионизирующей радиации на иммунитет
2.3 Сроки гибели животных после воздействия радиации в летальных дозах
2.4 Хозяйственно полезные качества животных, подвергшихся воздействию ионизирующей радиации
2.5 Воспроизводительные способности животных
Список литературы
ВВЕДЕНИЕ
Во время радиоактивного распада ядер испускаются α-, β- и γ- лучи, обладающие ионизационной способностью. Облучаемая среда частично ионизируется поглощаемыми лучами. Эти лучи взаимодействуют с атомами облучаемого вещества, что приводит к возбуждению атомов и вырыванию отдельных электронов из их электронных оболочек. В результате атом превращается в положительно заряженный ион (первичная ионизация). Выбитые электроны, в свою очередь, сами взаимодействуют со встречными атомами, вызывая вторичную ионизацию. Электроны, затратившие всю энергию, «прилипают» к нейтральным атомам, образуя отрицательно заряженные ионы. Число пар ионов, создаваемых в веществе ионизирующими лучами на единице длины пробега, называется удельной ионизацией, а расстояние, пройденное ионизирующей частицей от места ее образования до места потери энергии движения, называется длиной пробега.
Ионизирующая способность различных лучей неодинакова. Она наиболее высока у альфа-лучей. Бета-лучи вызывают меньшую ионизацию вещества. Самой низкой ионизационной способностью обладают гамма-лучи. Проникающая же способность наивысшая у гамма-лучей, а наинизшая - у альфа-лучей.
Не все вещества одинаково поглощают лучи. Высокой поглощающей способностью обладают свинец, бетон и вода, которые чаще всего и используют для защиты от ионизирующих излучений.
1 Факторы, определяющие реакцию растений на облучение
Степень поражения тканей и растительного организма в целом зависит от множества факторов, которые можно разделить на три основные группы: генетические, физиологические и условия внешней среды. К генетическим факторам относятся видовые и сортовые особенности растительного организма, которые в основной определяются цитогенетическими показателями (размером ядра, хромосом и количеством ДНК). Цитогенетические характеристики — размеры ядер, число и строение хромосом — определяют радиоустойчивость растений, которая находится в тесной зависимости от объема клеточных ядер. К физиологическим факторам относят фазы и стадии развития растений в момент начала облучения, скорость роста и обмен веществ растительного организма. К факторам внешней среды относят погодно-климатические условия в период облучения, условия минерального питания растений и т. д.
Объем клеточного ядра отражает содержание в нем ДНК, существует связь между чувствительностью растений к облучению и количеством ДНК в ядрах их клеток. Поскольку число ионизации внутри ядра пропорционально его объему, то чем больше объем ядра, тем больше повреждений хромосом будет приходиться на единицу дозы. Однако обратной пропорциональной зависимости между величиной летальной дозы и объемом ядра не наблюдается. Это обусловлено тем, что число и строение хромосом в клетках растений различных видов неодинаково. Поэтому более верным показателем радиочувствительности служит величина объема ядра в расчете на одну хромосому, т. е. отношение объема ядра в интерфазе к числу хромосом в соматических клетках (кратко называют объемом хромосом). В логарифмическом масштабе эта зависимость выражается прямой с тангенсом угла наклона, равным 1, т. е. между указанными характеристиками существует линейная связь (рис.).
Радиочувствительность различных растений при хроническом облучении (по А. Спэрроу)
| Растение | Продолжительность облучения, нед | Мощность дозы, Р/сут | ||
| винимое повреждение отсутствует | среднее повреждение | сильное повреждение | ||
| Сосна | 10 сезонов (120 недель) | 0,75 | 1 | 3,5 |
| Лилия | 8 | 10 | 20 | 40 |
| Традесканция болотная | 12 | 15 | 20 | 40 |
| Бобы конские | 15 | 30 | 60 | 90 |
| Хлопчатник | 15 | 25 | 112 | 250 |
| Махорка | 15 | 50 | 10.0 | 400 |
| Хризантема | 12 | 215 | 430 | 870 |
| Гладиолус | 12 | 800 | 1500 | 5000 |
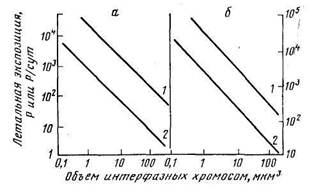
Зависимость радиочувствительности древесных (а) и травянистых (б) растений от объема интерфазных хромосом (по Спэрроу, 1965): 1—острое облучение (экспозиция в Р); 2 — хроническое об» лучение (экспозиция в Р/сут)
Из этого следует, что произведение двух величин — дозы (или мощности дозы) и объема хромосомы при данной степени лучевого повреждения — величина постоянная, т. е. при постоянном среднем числе ионизации в каждой хромосоме появляется одинаковая вероятность повреждения генетического материала клетки. Это означает, что для лучевого поражения клеток растений существенна не столько величина удельной поглощенной дозы (например, на 1 г ткани), сколько величина энергии излучения, поглощенной ядерным аппаратом. Обратная пропорциональность изоэффективных доз размерам хромосомного аппарата означает, что среднее количество энергии, адсорбированной хромосомами при экспозициях, необходимых для вызывания данного эффекта, примерно постоянно в пределах каждой растительной группы, т. е. для деревьев и трав. Изоэффективная доза — доза, оказывающая такой же (подобный) эффект.
На устойчивость растений к облучению влияет и степень плоидности растительных организмов. Более чувствительны диплоидные виды. Дозы, повреждающие полиплоидные виды, выше. Полиплоидные виды устойчивы к радиационному поражению и к действию других неблагоприятных факторов, поскольку обладают избытком ДНК.
Из физиологических факторов на радиочувствительность растений влияет скорость роста, т. е. скорость клеточного деления. При остром облучении зависимость радиочувствительности от скорости деления подчиняется закону Бергонье — Трибондо: растения обладают большей радиочувствительностью в стадии наиболее интенсивного роста, медленно растущие растения или их отдельные ткани устойчивее к действию облучения, чем растения или ткани с ускоренным ростом. При хроническом облучении проявляется обратная зависимость: чем выше скорость роста, тем меньше угнетаются растения. Это обусловлено интенсивностью деления клеток. Быстро делящиеся клетки накапливают за время одного акта клеточного цикла меньшую дозу и, следовательно, повреждаются слабее. Такие клетки имеют больше возможностей перенести облучение без существенного нарушения функций. Поэтому при облучении в сублетальных дозах любой фактор, увеличивающий продолжительность митоза или мейоза, должен усиливать радиационное повреждение, вызывая увеличение частоты наведенных излучением хромосомных перестроек и более сильное торможение скорости роста.
Критерий действия ионизирующих излучений на растения. Поскольку радиочувствительность — явление сложное, комплексное, определяющееся многими факторами, следует остановиться на тех методах оценки и критериях, по которым судят о степени радиочувствительности растений. Обычно в качестве таких критериев используют: подавление митотической активности при клеточном делении, процент поврежденных клеток в первом митозе, число хромосомных аберраций на одну клетку, процент всхожести семян, депрессии в росте и развитии растений, радиоморфозы, процент хлорофильных мутаций, выживаемость растений и в конечном результате урожай семян. Для практической оценки снижения продуктивности растений от воздействия радиации обычно используют два последних критерия: выживаемость растений и их урожай.
Количественная оценка радиочувствительности растений по критерию выживаемости устанавливается по показателю ЛД50 (или ЛД50, ЛД100). Это величина дозы, при которой погибает 50 % (или 70, 100 %) из числа всех облученных особей. Показатель ЛД50 может быть использован также и при оценке потерь урожая в результате радиационного поражения растений. В этом случае он показывает, при какой дозе облучения растений их урожай снижается на 50 %.
Радиочувствительность растений в разные периоды их развития. В процессе роста и развития радиочувствительность растений существенно изменяется. Это обусловлено тем, что в различные периоды онтогенеза растения отличаются не только морфологическим строением, но и разнокачественностью клеток, тканей, а также характерными для каждого периода физиолого-био-химическими процессами.
При остром облучении растений в различные периоды онтогенеза они реагируют по-разному в зависимости от этапа органогенеза в момент начала облучения (рис.). Радиация вызывает у растений поражение тех органов и смещение тех процессов, которые формируются и протекают в период воздействия. В зависимости от величины дозы облучения эти изменения могут носить либо стимулирующий, либо повреждающий характер.
Радиационное поражение растений в той или иной степени затрагивает все органы и все функциональные системы организма. Наиболее чувствительными «критическими органами», повреждение которых определяет развитие и результат радиационного поражения растений, являются меристематические и эмбриональные ткани. Качественный характер реакции растений на их облучение зависит от биологической специфичности морфофизиологического состояния растений в период накопления основной дозы облучения.
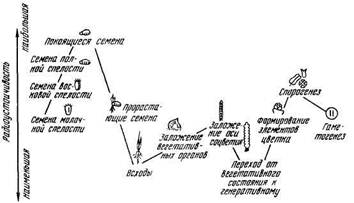
Колебания радиоустойчивости растений в онтогенезе (Батыгин, Потапова, 1969)
По поражению основного побега все культуры проявляют наибольшую чувствительность к действию радиации в первый период вегетации (I и III этапы органогенеза). Облучение растений в эти периоды тормозит ростовые процессы и нарушает взаимосогласованность физиологических функций, определяющих формообразовательные процессы. При дозах облучения, превышающих их критические значения для определенной культуры (ЛД70), во всех случаях наблюдается гибель основного побега злаковых растений.
Если растения подвергаются облучению на ранних этапах органогенеза (I и V), образуются дополнительные побеги, которые при благоприятных условиях сезона успевают дойти до созревания и дать урожай, компенсирующий в той или иной мере потери, связанные с гибелью главного побега. Облучение растений на VI этапе органогенеза — в период формирования материнских клеток пыльцы (мейоз) — может привести к значительной стерильности и потере урожая зерна. Критическая доза облучения (например, 3 кР для пшеницы, ячменя и гороха) в этот период вызывает полную стерильность соцветий основных побегов. Дополнительные побеги кущения или ветвления, развивающиеся у этих растений в сравнительно позднее время, не успевают завершить свой цикл развития и не могут компенсировать потери урожая с основных побегов.
При облучении растений на том же VI этапе органогенеза в период формирования одноядерных пыльцевых зерен устойчивость к действию ионизирующей радиации у растений значительно повышается. Например, при облучении пшеницы дозой 3 кР в период мейоза урожай зерна практически равен нулю, тогда как при облучении растений в период формирования одноядерной пыльцы наблюдается снижение урожая на 50%. На последующих этапах органогенеза устойчивость растений к действию радиации возрастает еще сильнее. Облучение растений в период цветения, эмбриогенеза и налива зерна при одних и тех же дозах не вызывает заметного снижения их продуктивности. Следовательно, к наиболее чувствительным периодам относятся прорастание семян и переход растений от вегетативного состояния к генеративному, когда закладываются органы плодоношения. Эти периоды характеризуются повышенной метаболической активностью и высокой интенсивностью клеточного деления. Наиболее устойчивы растения к радиации в период созревания и в период физиологического покоя семян (табл.). Злаковые культуры более радиочувствительны в фазы выхода в трубку, кущения и колошения.
Выживание озимых культур при их облучении в осенне-зимне-весенний период заметно повышается при посеве озимых культур в наиболее ранние из установленных сроков. Это объясняется, очевидно, тем, что облученные растения, уходя под зиму более окрепшими, в состоянии полного кущения, оказываются более устойчивыми к последствиям действия радиации.
Аналогичная закономерность снижения урожая зерна при облучении растений в разные фазы развития получена и на других культурах. Зерновые бобовые культуры обладают наибольшей радиочувствительностью в период бутонизации. Самое резкое снижение урожая овощных культур (капуста, свекла, морковь) и картофеля наблюдается при воздействии ионизирующего облучения в период всходов.
Все зерновые культуры обладают максимальной радиочувствительностью в фазе выхода в трубку. В зависимости от биологических особенностей растений наблюдается некоторое различие. Так, овес проявляет максимальную радиочувствительность в конце фазы выхода в трубку и в период выметывания метелки.
Снижение урожая зерна озимых зерновых культур (пшеница, рожь, ячмень) в зависимости от облучения растений γ-лучами в разные фазы развития растений, % к необлученному контролю
| Фаза развития | Доза облучения, Р | ||
| 1000 | 2000 | 3000 | |
| Кущение | 5 | 25 | 55 |
| Выход в трубку | 25 | 55 | 80 |
| Колошение | 15 | 20 | 28 |
| Цветение | 8 | 13 | 21 |
| Молочная спелость | 5 | 7 | 9 |
| Полная спелость | 0 | 0 | 0 |
Отрицательное действие внешнего γ-облучения меньше сказывается на продуктивности зерновых культур при их облучении в фазе кущения. При частичном повреждении растений происходит усиленное кущение и в целом снижение урожая компенсируется за счет формирования вторичных побегов кущения. Облучение зерновых культур в период молочной спелости не вызывает заметного повышения стерильности колосьев.
2 Действие внешнего ионизирующего излучения на организм 2.1 Варианты возможного радиационного воздействия
Источники ионизирующего излучения (радионуклиды) могут находиться вне организма и (или) внутри его. Если животные подвергаются воздействию излучения извне, то говорят о внешнем облучении, а воздействие ионизирующих излучений на органы и ткани от инкорпорированных радионуклидов называют внутренним облучением. В реальных условиях чаще всего возможны различные варианты и внешнего, и внутреннего облучения. Такие варианты воздействия называются сочетанными радиационными поражениями.
Доза внешнего облучения формируется главным образом за счет воздействия γ-излучения; α- и β-излучения не вносят существенного вклада в общее внешнее облучение животных, так как они в основном поглощаются воздухом или эпидермисом кожи. Радиационное поражение кожных покровов β-частицами возможно в основном при содержании скота на открытой местности в момент выпадения радиоактивных продуктов ядерного взрыва или других радиоактивных осадков.
Характер внешнего облучения животных во времени может быть различным. Возможны разные варианты однократного облучения, когда животные подвергаются радиационному воздействию в течение короткого промежутка времени. В радиобиологии принято считать однократным облучением воздействие радиации на протяжении не более 4 сут. Во всех случаях, когда животные подвергаются внешнему облучению с перерывами (они могут быть различными по продолжительности), имеет место фракционированное (прерывистое) облучение. При непрерывном длительном воздействии ионизирующего излучения на организм животных говорят о пролонгированном облучении.
Выделяют общее (тотальное) облучение, при котором радиационному воздействию подвергается все тело. Этот вид облучения имеет место, например, при обитании животных на территории, загрязненной радиоактивными веществами. Кроме того, в условиях специальных радиобиологических исследований может осуществляться местное облучение, когда радиационному воздействию подвергается та или иная часть тела! При одной и той же дозе облучения наиболее тяжелые последствия наблюдаются при общем облучении. Например, при облучении всего тела животных в дозе 1500 Р отмечается практически 100%-ная их гибель, тогда как облучение ограниченного участка тела (головы, конечностей, щитовидной железы и т. д.) каких-либо серьезных последствий не вызывает. В дальнейшем рассматриваются последствия только общего внешнего облучения животных.
2.2 Влияние ионизирующей радиации на иммунитетМалые дозы радиации, по-видимому, не оказывают заметного влияния на иммунитет. При облучении животных сублетальными и летальными дозами происходит резкое снижение резистентности организма к инфекции, что обусловлено рядом факторов, среди которых важнейшую роль играют: резкое повышение проницаемости биологических барьеров (кожи, дыхательных путей, желудочно-кишечного тракта и др.), угнетение бактерицидных свойств кожи, сыворотки крови и тканей, снижение концентрации лизоцима в слюне и крови, резкое уменьшение числа лейкоцитов в кровеносном русле, угнетение фагоцитарной системы, неблагоприятные изменения биологических свойств микробов, постоянно обитающих в организме, — увеличение их биохимической активности, усиление патогенных свойств, повышение резистентности и др.
Облучение животных в сублетальных и летальных дозах приводит к тому, что из крупных микробных резервуаров (кишечник, дыхательные пути, кожа) в кровь и ткани поступает огромное количество бактерий.! При этом условно выделяют период стерильности (его продолжительность одни сутки), в течение которого микробов в тканях практически не обнаруживается; период обсемененности регионарных лимфатических узлов (обычно совпадает с латентным периодом); бактериемический период (длительность его 4—7 дней), который характеризуется появлением микробов в крови и тканях, и, наконец, период декомпенсации защитных механизмов, в течение которого отмечается резкое возрастание количества микробов в органах, тканях и крови (этот период наступает за несколько дней до гибели животных).
Под действием больших доз радиации, вызывающих частичную или полную гибель всех облученных животных, организм оказывается безоружным как к эндогенной (сапрофитной) микрофлоре, так и к экзогенным инфекциям. Считают, что в период разгара острой лучевой болезни и естественный, и искусственный иммунитет сильно ослаблен. Однако имеются данные, указывающие на более благоприятный исход течения острой лучевой болезни у животных, подвергшихся иммунизации до воздействия ионизирующего излучения. Вместе с тем экспериментально установлено, что вакцинация облученных животных отягощает течение острой лучевой болезни, и по этой причине она противопоказана до разрешения болезни. Напротив, через несколько недель после облучения в сублетальных дозах выработка антител постепенно восстанавливается, и поэтому уже через 1—2 мес после радиационного воздействия вакцинация вполне допустима.
2.3 Сроки гибели животных после воздействия радиации в летальных дозахПри однократном облучении сельскохозяйственных животных в дозах, вызывающих крайне тяжелую степень острой лучевой болезни (более 1000 Р), обычно они погибают в течение первой недели после радиационного воздействия. Во всех других случаях летальные исходы острой лучевой болезни наблюдаются чаще всего на протяжении 30 дней после облучения.1! Причем после однократного облучения большая часть животных погибает между 15-м и 28-м днями (рис.); при фракционированном облучении летальными дозами гибель животных происходит в течение двух месяцев после радиационного воздействия (рис.).
Как правило, молодняк погибает в более ранние сроки после облучения в летальных дозах: смертность животных обычно отмечается на 13—18-й день. Для всех возрастных групп животных, облученных в летальных дозах, характерна более ранняя гибель при наиболее высоких дозах радиационного воздействия (рис.). Однако это явление можно расценивать скорее как тенденцию, чем закономерность, так как имеется достаточно много экспериментальных данных о ранних сроках гибели животных при облучении их сравнительно невысокими дозами радиации.
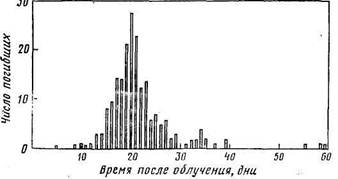
Смертность овец после внешнего γ-облучения летальными дозами (Пейч в др., 1968)
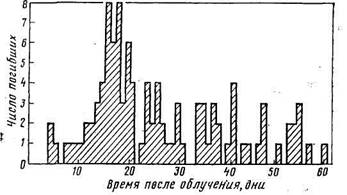
Смертность коз, подвергшихся фракционированному рентгеновскому облучению (Тайлор и др., 1971)
Следует иметь в виду, что при фракционированном облучении сроки гибели животных зависят прежде всего от мощности дозы. Так, при ежедневном облучении ослов в дозе 400 Р все животные погибали между 5-м и 10-м днями. В экспериментах, где доза ежедневного облучения составляла 50 и 25 Р, средняя продолжительность жизни после начала радиационного воздействия составляла соответственно 30 и 63 дня. Кроме того, на продолжительность жизни сильно влияют видовые особенности животных. При фракционированном ежедневном облучении свиней в дозе 50 Р средняя продолжительность жизни у них оказалась равной 205 дням, что в 6,4 раза превышало среднюю продолжительность жизни ослов при тех же условиях радиационного воздействия.

Смертность коров в различные сроки после γ-облучения (Броун и др., 1961)
2.4 Хозяйственно полезные качества животных, подвергшихся воздействию ионизирующей радиацииВ принципе все сельскохозяйственные животные, подвергшиеся действию ионизирующих излучений, могут быть разделены на две категории. К первой категории относятся животные, получившие летальные дозы радиации. Срок их жизни от момента облучения сравнительно невелик, но в некоторых ситуациях продуктивность смертельно пораженных животных может представлять известный интерес.
Молочная продуктивность коров в первые 10— 12 дней после радиационного воздействия изменяется незначительно, а затем резко падает, и уже за 2 дня до гибели животных лактация полностью прекращается. Мясная продуктивность животных, которая обычно характеризуется динамикой живой массы, также изменяется незначительно: снижение массы тела у смертельно пораженных животных (если оно имеет место), как правило, не превышает 5—10%. Яйцекладка у кур-несушек, подвергшихся воздействию летальных доз радиации, прекращается в течение ближайших 5—7 дней. О шерстной продуктивности летально пораженных овец говорить не приходится, так как у них через 7—10 дней после радиационного воздействия наблюдается интенсивная эпиляция.
У животных, выживших после облучения в летальных или сублетальных дозах (вторая категория), продуктивность снижается ненадолго. Например, при облучении коров за 60 дней до отела в дозе 400 Р их молочная продуктивность на протяжении первых 10— 12 нед была ниже контроля на 5—10%. После повторного облучения в дозе 350 Р через 18 нед после начала лактации удой в течение первой недели после облучения снизился на 16%, к 5-й неделе —на 8%, а на 6-й неделе молочная  продуктивность облученных коров вернулась к норме. Ориентировочно можно считать, что облучение коров в дозах, которые могут вызвать частичную гибель дойного стада, приводит к снижению удоя в целом за лактацию в среднем на 5—8 %.
продуктивность облученных коров вернулась к норме. Ориентировочно можно считать, что облучение коров в дозах, которые могут вызвать частичную гибель дойного стада, приводит к снижению удоя в целом за лактацию в среднем на 5—8 %.
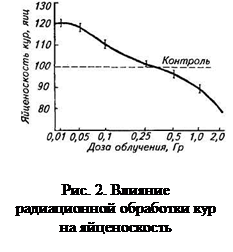
У выживших животных, подвергшихся воздействию радиации в полулетальных дозах (или близких к ним), отмечены также другие неблагоприятные последствия. Так, после двукратного облучения свиней (480 рад + 460 рад через 4 мес) отмечено снижение прироста массы: спустя 2 года после радиационного воздействия облученные животные имели массу тела на 45 кг ниже, чем контрольные свиньи. Продолжительность жизни свиней сокращается в среднем на 3 % на каждые 100 рад внешнего облучения животных (рис.). При облучении кур породы белый леггорн в дозе 800 Р (смертность кур составляла в среднем 20%) наблюдается заметное снижение яйцекладки (рис.).
Дозы облучения, вызывающие острую лучевую болезнь легкой или средней степени тяжести, обычно не отражаются заметным образом на продуктивности сельскохозяйственных животных. Например, после внешнего γ-облучения в дозе 240 Р в течение последующих 40 нед бычки имели прирост массы тела 131 кг (в контрольной группе 118 кг). Свиньи, подвергавшиеся хроническому облучению в дозах 360—610 Р (мощность дозы 1,4 Р/ч), в течение всего времени облучения и последующие 90 дней опыта имели достаточно высокий среднесуточный прирост (500—540 г) и по этому показателю не отличались от контрольных групп (примерно 470 г). Аналогичная картина наблюдалась и при фракционированном облучении свиней в дозе 50 Р/сут. Не было обнаружено снижения яйцекладки у кур после облучения их в дозе 400 Р, а при дозе 600 Р яйцекладка снижалась примерно на 20 % только в первой декаде после воздействия.
Таким образом, при облучении сельскохозяйственных животных в сублетальном диапазоне доз существенных изменений в их продуктивных качествах не отмечается (если, конечно, животным созданы нормальные условия содержания и они обеспечены соответствующими рационами). При облучении животных абсолютно летальными дозами продуктивность снижается, но качество животноводческой продукции остается достаточно высоким. При длительном скармливании животным продукции, полученной от смертельно пораженных радиацией овец и коров, не наблюдается каких-либо патологических изменений как у потребляющих эту продукцию, так и у их потомства. Однако при использовании для питания продукции от радиационно пораженных животных рекомендуется проводить особо тщательно бактериологические исследования и соответствующую кулинарную обработку.
2.5 Воспроизводительные способности животныхПоловые железы животных отличаются высокой чувствительностью к действию ионизирующих излучений. При облучении самцов сублетальными дозами происходит лучевое поражение семяродного эпителия в семенных канальцах, а также сперматогониев и сперматоцитов; созревшие и сформированные сперматозоиды считаются радиорезистентными. Высокие дозы радиации вызывают почти полное уничтожение семяродного эпителия и последующее затухание спермопродукции, тогда как облучение самцов средними и низкими дозами вначале приводит к снижению сперматогенеза, а затем отмечается постепенное его восстановление (рис.). Весьма характерны уменьшение объема эякулята, снижение концентрации и подвижности спермиев в эякуляте, появление в большом количестве уродливых сперматозоидов, падение биологической полноценности спермы и ее оплодотворяющей способности. Кроме того, уменьшается масса семенников: при γ-облучении хряков в дозе 400 Р масса тестикулов снизилась на 30%, а при облучении петушков в дозе 500 Р она уменьшилась в 3 раза по сравнению с массой семенников у контрольных петушков.
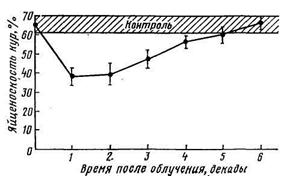
Влияние внешнего γ-облучения кур в дозе 800 Р на яйценоскость выживших кур (Малоний, Мрац, 1969)
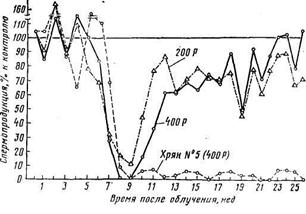
Спермопродукция хряков, подвергшихся воздействию внешнего γ-облучения в сублетальных дозах (Паке в др., 1962).
Облучение в дозе 400 Р у отдельных хряков вызывает длительное бесплодие (хряк № 5)
Если дозы облучения не слишком велики, то с течением времени наблюдается частичное или полное восстановление воспроизводительной функции у самцов. В опытах на баранах, например, было установлено, что при облучении в дозе 100 Р качество спермы восстанавливается уже через 4 мес, в дозе 430 Р — лишь через 12 мес. Заметим, что аналогичное восстановление качества спермы у облученных хряков и быков наблюдалось уже через 5—б мес, т. е. примерно вдвое быстрее, чем у баранов.
Ионизирующая радиация влияет и на репродуктивную функцию самок. У облученных животных повреждаются и частично гибнут все виды клеток функционирующего яичника (в особенности первичные и вторичные фолликулы, зрелые яйцеклетки), нарушаются астральные циклы. Следует, однако, иметь в виду, что вскоре после облучения (даже среднелетальными дозами) воспроизводительная функция у самок восстанавливается и они могут приносить жизнеспособное потомство. Например, не было отмечено снижения плодовитости у взрослых коров, подвергшихся двукратному (с перерывом в 2 года) радиационному воздействию в дозах 400 Р.
Наиболее тяжелые последствия наблюдаются при воздействии ионизирующей радиации на животных в период их внутриутробного развития. Большая часть зародышей погибает в предимплантационный период, т. е. в период, когда еще не произошло внедрения развивающегося оплодотворенного яйца в толщу слизистой оболочки матки (у овец и свиней — в первые 13, у коров — в первые 15 дней после оплодотворения), или подвергается резорбции (рассасыванию) сразу же после имплантации. При облучении беременных животных в период основного органогенеза (у овец — на 17— 19-й, у свиней — на 15—18-й, у коров — на 22—27-й день) даже при сравнительно невысоких дозах радиационного воздействия (200—300 Р) во многих случаях возможна резорбция эмбриона, а у выживших эмбрионов наблюдаются отставание в росте, появление пороков развития, увеличение смертности новорожденных, сокращение продолжительности жизни. Например, при облучении сукрольных самок на 12—14-й день беременности в дозе 400 Р наблюдали случаи сросшихся пальцев передних и задних конечностей у потомства. При облучении животных на более поздних стадиях беременности радиочувствительность плодов несколько снижается.
При исследовании последствий действия ионизирующих излучений на организм в период внутриутробного развития была обнаружена исключительно высокая чувствительность воспроизводительной системы плода к действию радиации. При хроническом облучении свиноматок в течение 108 дней беременности (дозы γ-облучения от 1 до 20 рад/сут, длительность ежесуточного облучения 22 ч) беременность у животных протекала нормально, общее состояние свиноматок, число живых поросят в помете и их послеродовая жизнеспособность не отличались от тех же показателей в контрольных группах животных. Вместе с тем даже при облучении супоросных свиноматок в дозе 1 рад/сут у новорожденных поросят обнаруживается существенное снижение общего числа зародышевых клеток (у животных обоих полов). Так, у боровков количество гоноцитов (первичных предшественников половых клеток) составляло всего 3 % контроля, а у самок число выживших ооцитов было равным 7 % ооцитов контрольных свинок. Облучение в утробный период развития было причиной снижения спермопродукции (на 83 %), увеличения числа дефектных сперматозоидов с 2,8 % (контроль) до 11,4 °/о, что повлекло за собой бесплодие у 4 из 10 хряков. Несмотря на существенное снижение количества первичных и растущих фолликулов у облученных свинок, их воспроизводительные способности в первом помете были такими же, как у контрольных животных, но при повторном спаривании у 4 из 23 свиноматок было установлено бесплодие. Облучение супоросных свиноматок в дозе 0,25 рад/сут практически не влияет на воспроизводительную функцию у потомства.
Список литературы
1. Анненков Б.Н., Юдиннева Е.В. Основы сельскохозяйственной радиологии.— М.: Агропромиздат, 1991. — 287 с: ил.
2. Старков В.Д., Мигунов В.И. Радиационная экология. Тюмень: ФГУ ИПП «Тюмень», 2003, 304 с.
Похожие работы
... действии ионизирующих излучений на животный организм можно судить по ускорению или повышению под влиянием облучения таких факторов, как рост, развитие, продуктивность. Первые попытки практического использования ионизирующих излучений для повышения продуктивности сельскохозяйственных животных были сделаны в птицеводстве путем облучения яиц до и после инкубации, а также облучения цыплят и кур в ...
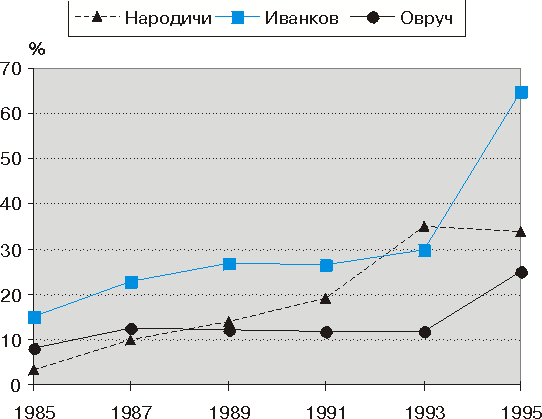
... зависит будущее нации. На пострадавших территориях Украины, где плотность радиоактивного загрязнения по 137Cs составила от 5 до 40 Ku/км2, возникли условия длительного воздействия малых доз ионизирующего излучения, влияние которого на организм беременной и плода до Чернобыльской катастрофы фактически не изучалось. С первых дней аварии велось тщательное наблюдение за состоянием здоровья ...

... частиц (α-, β-частиц, протонов, электронов), кинетическая энергия которых достаточна для ионизации атомов при столкновении, относится к классу непосредственно ионизирующего излучения. Нейтроны и другие элементарные частицы непосредственно не производят ионизацию, но в процессе взаимодействия со средой высвобождают заряженные частицы (электроны, протоны), способные ионизировать атомы и ...
... , добавленным к хлороформу. По плотности окраски судят о дозе излучения (поглощенной энергии). На этом принципе основано устройство химических дозиметров ДП-70 и ДП-70М. Сцинтилляционныи метод измерения ионизирующих излучений основан на том, что некоторые вещества (сульфит цинка, иодид натрия) светятся при воздействии на них ионизирующих излучений. Количество световых вспышек пропорционально ...
0 комментариев