Міністерство освіти і науки України
Чернігівський державний педагогічний університет імені Т.Г. Шевченка
Кафедра ботаніки, зоології та охорони природи
КУРСОВА РОБОТА
Поняття „Консорції” в сучасній синекології та її значення у структуруванні біоценозів
Чернігів – 2007
Зміст
Вступ
Розділ 1. Поняття про консорцію та особливості взаємодії в ній організмів
Розділ 2. Особливості біотичних взаємодій між організмами в консорції
2.1 Паразитизм
2.2 Алелопатія, або антибіоз
2.3 Позитивна взаємодія: коменсалізм, протокооперація, мутуалізм
Розділ 3. Середовищні зв`язки
Висновки
Список використаної літератури
Вступ
Актуальність. Біоценоз це свого роду сукупність різних живих організмів, які заселяють певну ділянку простору та характеризуються певними стосунками між собою та пристосованістю до середовища. Вивчення трофічних зв'язків свідчить, що в природі жодний живий організм не може існувати ізольовано. Більше того, живі організми в межах біогеоценозів або ж біогеоценотичних комплексів створюють спеціалізовані об'єднання — консорції.
До кожної екологічної структури (об`єднання) входить багато різних видів, які належать до продуцентів, консументів чи редуцентів. В кожному угрупованні існують харчові зв`язки. Харчові зв`язки між цими видами утворюють трофічну мережу, або мережу живлення. Суть їх полягає в тому, що в меню будь-якого виду, за винятком продуцентів, входить не один, а кілька або багато інших видів. Кожен з видів, своєю чергою, може бути їжею для кількох інших. У результаті безліч ланцюгів живлення у біогеоценозі складно переплітаються між собою.
Мета роботи: проаналізувати поняття консорції та її значення в структурі біоценозів.
Завданнями роботи є наступне:
1) розглянути поняття консорції;
2) охарактеризувати особливості взаємодії між організмами в консорції;
3) проаналізувати особливості біотичних взаємодій у консорції;
4) розглянути середовищні взаємодії в консорції.
Розділ 1. Поняття про консорцію та особливості взаємодії в ній організмів
Консорція, (від лат. consortіum - співучасть, співтовариство) у биогеоценологи і, сукупність популяцій організмів, життєдіяльність яких зв'язана з центральним видом фітоценозу - автотрофною рослиною. У ролі центрального виду консорції звичайно виступає едифікатор - основний вид, що визначає особливості фітоценозу. Іноді інші види, у тому числі гетеротрофні організми (паразити, сапрофіти), зв'язані з центральним видом односторонньо - споживання, затінення. Це ядро консорції - джерело існування 2-го, 3-го і наступних його кіл, куди входять організми, що руйнують органічні речовини, створені центральним і ін. автотрофними видами, і використовують укладену в них енергію. Терміном "Консорція" у 70-х р. 19 ст. позначали симбіоз деяких рослин (наприклад, водорості і гриба, що утворять лишайник) [8].
Характерною особливістю цих об'єднань є те, що вони звичайно формуються на базі популяцій автотрофних рослин (сосна, смерека, береза, пшеничне чи картопляне поле тощо). Такі популяції називають детермінантами, а види, які об'єднуються довкола них, — консортами. На рис. 1 зображено консортне об'єднання, сформоване довкола ядра, яким стала трав'яниста рослина, даючи поживу численним фітофагам. Звичайно консорцію зображають у вигляді концентричної фігури (рис. 3) з рослиною-ядром (детермінантом) та оточуючими його консортами, які, в свою чергу, утворюють консорти першого, другого, третього порядку і т.ін.
Всі види, які створюють біоценоз, функционально пов`язані с едифікаторами. Мінімальною своєрідною структурною одиницею біоценозу, яка відображає цей зв`язок являеться консорція. Найбільш детально вчення про консорціях розробили В.Н. Беклемишев и Л.Г. Раменський в 50-х р.
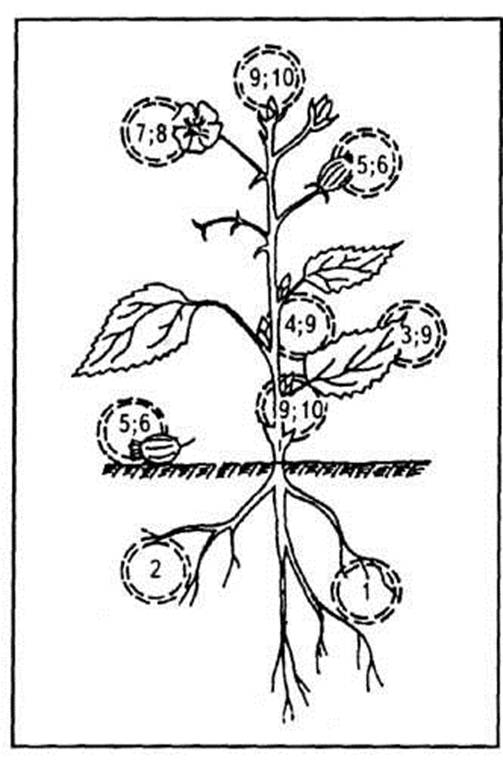
Рис. 1. Екологічні ніші серед фітофагів: 1 — коренеїди; 2 — ті, що поїдають кореневі виділення; 3 — листоїди; 4 — стовбуроїди; 5 — плодоїди; 6 — насіннєїди; 7 — квіткоїди; 8 — пилкоїди; 9 — сокоїди; 10 — брунькоїди
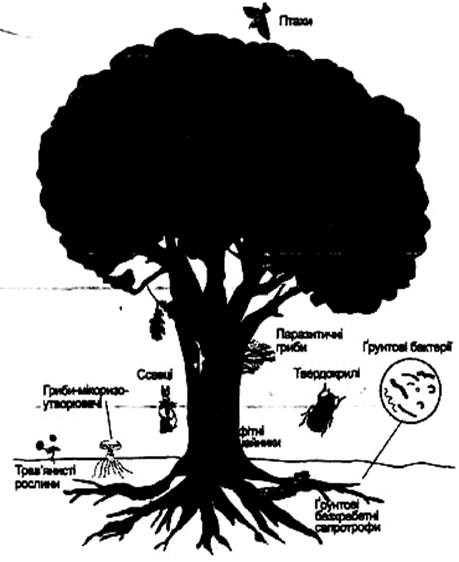
Рис. 2. Консорція
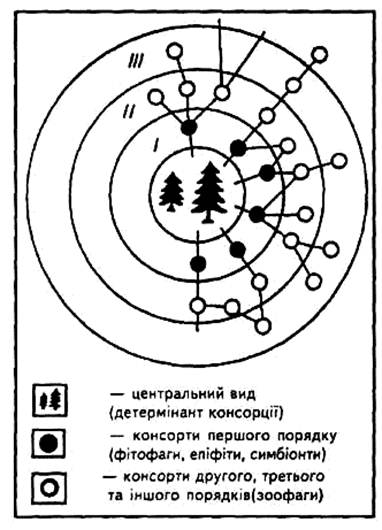
Рис. 3. Схематична будова консорції: І, ІІ, ІІІ — концептори
Едифікатори – види, які визначають структуру біоценозу і специфічні умови існування в ньому (ступінь затінення, умови росту, характер процесів кругообігу).Найчастіше едифікаторами виступають рослини верхніх ярусів. Едифікатори завжди з числа домінантів, але не завжди домінанти є едифікатором [8].
Асектатори (види супутники), які завжди присутні в біоценозі, але ніколи не бувають домінантними.
У складі фітоценозів виділяють групи видів, які мають різне значення в житті рослинного угруповання, їх називають ценотипами. Російські геоботаніки Г.М. Висоцький та І.К. Висоцький виділили дві групи таких видів:
1) основні, постійні види, які визначають властивості у фітоценозів;
2) тимчасові, непостійні види, які з’являються у фітоценозі періодично (інградієнти).
Топічні зв'язки виникають тоді, коли види не пов'язані прямими трофічними зв'язками, але внаслідок життєдіяльності одних видів змінюються умови існування інших. Наприклад, часто одні види мешкають на поверхні тіла інших (вусоногі раки — на молюсках, китах тощо, орхідеї — на стовбурах дерев і т.п.). Ярусність рослинних угруповань — яскравий приклад топічних зв'язків.
Завдяки топічним зв'язкам одні організми можуть створювати сприятливі умови для існування інших (наприклад, лишайники забезпечують передумови для заселення вищими рослинами субстратів, де їх раніше не було) або, навпаки, робити його неможливим. Так, багато рослин виділяє спеціальні речовини (фітонциди, антибіотики тощо), які пригнічують життєдіяльність інших видів рослин, грибів, бактерій [12].
В результаті топічних зв’язків відбувається “кондиціонування” середовища, тобто деякі організми створюють своєрідні, сприятливі або несприятливі фізичні або хімічні умови для поселення інших організмів. Наприклад, створення сприятливого середовища одним видом для поселення другого (температура, кількість опадів, освітленість). Усе це може сприяти поселенню одних організмів і не сприяє іншим.
У лісі створюється свій мікроклімат, який характеризується підвищеною вологістю, малою контрастністю температур, меншою освітленістю. Це сприяє росту рослин і розміщенню їх за ярусами.
Завдяки трофічним і топічним, які часто переплітаються між собою, організми об’єднуються в досить стабільні угруповання різних рівнів, що входить до біоценозу. Внаслідок чого в біоценозі встановлюються досить складні міжвидові взаємозв’язки, які становлять основу їх існування.
Між організмами біоценозу існують також форичні та фабричні відносини. Форичні відносини це такі відносини при яких одні організми сприяють переміщенню інших. Наприклад, великі ссавці – олені, лосі тощо спричиняють поширенню плодів і насіння рослин, населення паразитів на тілі цих тварин або птахів. Фабричні відносини – це зв’язки при яких один вид тварин використовує частини іншого виду рослин чи тварин для будівництва жител (гнізд). Наприклад бобри використовують гілки дерев для будівництва житла, птахи гілки листя тощо [5].
У фітоценозі розрізняють три основні форми впливу рослин між собою: контактні, трансабіотичні, і трансбіотичні. Контактні відношення – це механічний вплив рослини на рослину, паразитизм, симбіоз тощо; трансабіотичні – вплив одних рослин на інші шляхом зміни навколишнього середовища (боротьба за світло, вологу, поживні речовини та виділення рослинами продуктів життєдіяльності в навколишнє середовище тощо); трансабіотичні – впив рослин на рослину через інші організми. Звичайно у фітоценозі різні форми взаємовпливу проявляються одночасно.
Структура біоценозу – це закономірні зв’язки і визначений розподіл різних елементів системи. Розрізняють видову, просторову (або хорологічну) і трофічну структури.
Вивчення біоценозу розпочинають з видового складу даного угруповання. Визначення видового складу — це надзвичайно складна і копітка робота, яка вимагає спеціальних знань і великих трудових затрат. Складність її полягає в тому, що угруповання значно різняться між собою видовим різноманіттям, яке є одним з основних показників структури біоценозу [8].
З одного боку, охарактеризувати видовий склад угруповання нескладно: перелічити види, які в ньому виявлені, скласти їх список і, нарешті, дати оцінку видового багатства. З іншого боку, трудність полягає в тому, що не всі ділянки обстежуваного нами угруповання мають однакові кількісні показники видового складу (рис. 4). Як свідчать результати досліджень, плато досягається не завжди. Тому видове багатство різних угруповань можна досліджувати, користуючись вибірками однакових розмірів (наприклад, ділянки площею 50 чи 100 м2) і сумарної кількості організмів або модулів на всіх пробних площах.
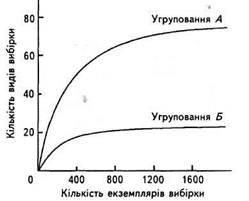
Рис. 4. Залежність видового багатства чисельності окремих організмів у двох гіпотетичних угрупованнях. В угрупованні А видове багатство значно вище, ніж в угрупованні Б.
Однак у згаданій бучині ростуть не лише дерева, але й кущі і кущики, трави, її заселяють співочі птахи та білки (а вони нас можуть цікавити з точки зору рекреаційного використання лісу). Як мовою математики показати це співвідношення, що має не лише теоретичне, а й суто прикладне значення (частка присутності рослин-медоносів чи рослин і тварин, які занесені до Червоної книги)?
Найпростішим для визначення є індекс різноманітності (Уіттекер, 1980) [8], де головним виміром є кількість видів5, описаних на площі стандартних розмірів; S — це ще одиниця виміру видового багатства, на відміну від щільності популяції.
Зі збільшенням кількості видів в угрупованні S максимальні значення обох показників, особливо показника Шеннона, збільшуються. Тому для одержання незалежного від кількості видів S характеристик звертаються до нормування шляхом ділення на максимальне значення того чи іншого показника. Цей показник набуває значень в інтервалі від 0 до 1.
До біоценозів з низьким видовим різноманіттям слід віднести екосистеми, які перебувають у стадії деградації: рекреаційні ліси та пасовища з високим рівнем витоптування, урбанізовані біоценози, а також культурні біоценози (лісові культури, посіви та сади, газони). Біоценозів зі середнім різноманіттям (порядку 0,5) порівняно небагато. Групи біоценозів з високою видовою різноманітністю охоплюють природні екосистеми, які неодержують із зовні концентрованої енергії та біогенних матеріалів і живуть лише за рахунок розсіяної енергії сонячного випромінювання (ліси і степи, озера, збагачені стоками водозбірних басейнів). Сюди слід віднести стабільні екосистеми вологих тропіків та дно океану.
Видова різноманітність угруповань змінюється в часі й є наслідком складних процесів імміграції й еміграції видів, які відбуваються в кожній екосистемі з більшою чи меншою інтенсивністю. Нова наука палеоекологія дає можливість, вивчаючи викопні рештки тваринних і рослинних організмів, встановити, який був видовий склад угруповань минулого, простежити процес їхньої еволюції. Наприклад, якщо викопні знахідки свідчать про те, що 10 тис. років тому, тобто після останнього льодовикового періоду, на місці сучасних дібров росли смерекові ліси, то в нас є всі підстави вважати, що 10 тис. років тому клімат був холоднішим, оскільки сучасні види смереки адаптовані до холоднішого клімату більше, ніж види дуба.
Усі біоценози суші і вод характеризуються нерівномірністю просторового розміщення. Виняток становлять штучні, або культурні, біоценози, зокрема, поля, плантації, лісові культури, сади, лісосмуги, газони тощо.
Просторова неоднорідність біоценозів зумовлена неоднорідністю чи мозаїчністю місцезростань, які колонізували організми. Заселенням біотопу організмами починається цикл змін у середовищі, спричинений активністю біологічних компонентів.
Просторову структуру біоценозу можна розглядати в декількох різних аспектах: поверховість, чи ярусність, біоценозів; розмежування біоценозів, що має два типи структури (перша є характерною для водних екосистем з їх шарами більшої і меншої освітлюваності водної товщі; друга більше відповідає біоценозам суші, між якими утворюється перехідна зона, названа екотоном).
Неоднорідність середовища і біоценозів, яка трапляється у вигляді окремих біоценотичних зон, розглядається вченими як явище мозаїчності і становить предмет окремих досліджень структури (рис.5).
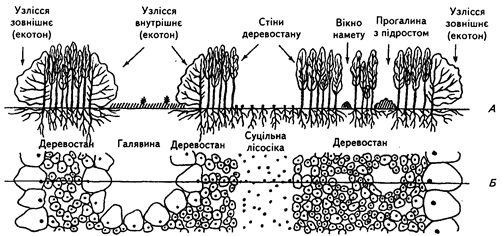
Рис. 5. Вертикальна (А) і горизонтальна (Б) структури лісового біоценозу.
Розділ 2. Особливості біотичних взаємодій між організмами в консорції 2.1 Паразитизм
Враховуючи особливості функціональної дії паразитів, розглянемо їх окремо. Паразит — організм, який живе на поверхні або в органах і тканинах інших організмів (живителів), живиться за їх рахунок, одержуючи від них готові речовини, та завдає їм певної шкоди. Розрізняють ендопаразитів, що селяться всередині живителя, і ектопаразитів, що існують на поверхні організму живителя. Виділяють також облігатні та факультативні форми паразитизму [12].
Паразити, подібно до хижаків з пасовищним типом живлення, поїдають лише частини своєї жертви (господаря). З того часу, — підкреслює Р.Уіттекер, — як паразит став постійно залежати від свого господаря як джерела живлення, йому стало невигідно убивати його. На відміну від справжніх хижаків і хижаків із пасовищним типом живлення, які вбивають велику кількість жертв, паразит тісно пов'язаний зі своєю жертвою-господарем. Типовим прикладом паразита є туберкульозна паличка, яка "роз'їдає" легені ослабленого людського організму. Стосовно дерева, наприклад берези, паразитом є гриб — березовий трутовик, який своїми гіфами висмоктує з неї поживні соки.
Паразитоїдами називають групу комах, головним чином перетинчастокрилих, а також багатьох двокрилих (Оіріега),в яких дорослі самки і їхні личинки вирізняються подібністю поведінки. Комахи-паразитоїди відкладають яйця в тіло іншої комахи або ж на його поверхню, тоді як комахи-господарі, як правило, ще не досягли дорослого віку. На їх тілі (або на їх поверхні, або поряд) із вилупленого яйця з'являється личинка, яка починає інтенсивно поїдати свого молодого господаря. Таким чином, господар гине, не досягши зрілого стану — в стадії лялечки або й раніше (яскравим прикладом такої поведінки є оса, яка паразитує на бджолах).
Паразитоїди тісно пов'язані з певним організмом господаря (подібно до паразитів), вони не призводять до швидкої загибелі господаря (подібно до паразитів і хижаків із пасовищним типом живлення), однак з часом господар неминуче загине (цим паразитоїди нагадують справжніх хижаків). Виділення паразитоїдів у окрему групу є цілком виправданим: за деякими даними до них належить близько 25% усіх видів, які живуть сьогодні на нашій планеті. Оскільки один вид комах має як мінімум одного паразитоїда, а останній — свого власного паразитоїда, то це свідчить про величезну кількість паразитоїдів.
Екологи добре вивчили природу цієї групи комах, використовують їх для боротьби зі шкідниками. Ю. Одум (1986) [12] наводить приклад використання одного з видів хижого кліща, який ефективно контролює чисельність рослиноїдного кліща на посадках суниць у Каліфорнії. Водночас автор звертає увагу на неефективність використання неспеціалізованих хижаків, а часто й на непередбачене лихо, яке можуть спричинити некваліфіковані заходи біологічного захисту. Наприклад, мангусти, яких завезли на острови Карибського басейну для боротьби зі щурами на плантаціях цукрової тростини, почали масово поїдати птахів, які гніздилися на землі. Чисельність цих птахів зменшилась більше, ніж чисельність щурів.
2.2 Алелопатія або антибіозАлелопатія — один із прикладів аменсалізму. Це явище взаємного антагонізму серед рослин, яке виявляється у виділенні хімічних речовин, отруйних для іншого виду і не шкідливих для виду, що їх виділяє. Хімічні виділення є продуктами метаболізму — обміну речовин: ефірні олії, фітонциди, глюкозиди, які часто називають колінами. Найчастіше алелопатія проявляється у конкурентному витісненні одного виду іншим. Ще в давнину помітили, що під горіхом, як правило, не росте трав'яна рослинність. Подібну властивість проявляє дуб звичайний і особливо дуб північний. Пирій та інші бур'яни витісняють культурні рослини. Листя каштанів, яке розкладається, виділяє токсичні речовини, здатні пригнічувати ріст сіянців багатьох рослин.
Алелопатія суттєво впливає на швидкість рослинних сукцесій і на видовий склад стабільних угруповань. Хімічні виділення можуть впливати двояко: в одних випадках вони зменшують видову різноманітність, в інших (за рахунок адаптивних пристосувань, які складаються в умовах диференціації ніш) підтримують високу видову різноманітність [8].
Не завжди рослина виділяє токсини за допомогою коріння. Вважають малоймовірним, наприклад, що шавлія витісняє із угруповання інший вид саме цим способом. Дослідники спостерігали, що довкола групи рослин шавлії утворюється гола ділянка, яка відокремлює ці рослини від сусідніх ділянок, вкритих травою. Коріння, як виявилося, доходить лише до оголеної ділянки і його не можна вважати агентом негативного впливу на цю територію. Виявляється, що листя шавлії продукує леткі терпени (клас органічних сполук, до яких належить камфора), які, на думку Р.Ріклефса, впливають на рослини, що ростуть поблизу, безпосередньо через атмосферу. Сильні дощі виводять ці токсини із атмосфери в ґрунт, знижуючи антибіотичну дію шавлії на інші види. Цим зумовлюється відсутність безпосередньої хімічної конкуренції в регіонах з вологим кліматом. Водночас терпени, які виробляє шавлія і які подавляють ріст інших рослин, у процесі еволюції вироблялися не як засіб конкуренції, а як аттракторний (приваблюючий) засіб для його опилювачів, зокрема бджіл.
Алелопатія відома не лише в світі вищих рослин, але й серед мікроорганізмів. Наприклад, гриб мікроміцет Ріпісіііит пригнічує розвиток багатьох бактерій своїми хімічними інгібіторними виділеннями.
2.3 Позитивна взаємодія: коменсалізм, протокооперація, мутуалізм
Життя організмів характеризується не лише боротьбою, але й і багатьма формами взаємовигідного існування, яке склалося в процесі еволюції. Це, як зазначено вище, коменсалізм, протокооперація і мутуалізм.
Позитивні взаємодії популяцій розглянемо в тій послідовності, в якій вони формувалися в процесі еволюції: коменсалізм (перевагу чи користь має одна популяція), протокооперація (користь одержують обидві популяції) і мутуалізм (користь одержують обидві популяції, причому вони повністю залежать одна від одної) [3].
Коменсалізм полягає передусім в односторонньому наданні притулку одним видом іншому. Характерним є те, що вид, який одержав для себе певну користь, не завдає помітної шкоди своєму доброзичливому господарю. Так поводить себе, наприклад, мальок ставриди під куполом медузи чи в мантійній порожнині каракатиці. Чимало коменсалів призвичаїлись жити в чужих норах, гніздах, дуплах, а також у мурашниках і термітниках, використовуючи середовище тварин-партнерів зі стабільним і сприятливим мікрокліматом.
Значно поширене прикріплення одних рослин до поверхні інших, причому перші використовують їх як опору (рослини-епіфіти). До епіфітів, зокрема, належать лишайники, які прикріпляються до кори дерев. У водяних угрупованнях рослини і тварини покриті плівкою з мікроорганізмів. Окремі види водяних тварин народжуються на поверхні інших, наприклад, на клешнях раків живуть п'явкоподібні черви, рибки-прилипки влаштовуються на тілі акул; деякі креветки живуть в сплетіннях отруйних щупалець актиній.
Протокооперація (взаємокорисне об'єднання двох популяцій, зв'язки яких необлігатні (необов'язкові)). Питанням протокооперації багато уваги приділяв В.Оллі, який вважав, що початкові стадії кооперації трапляються в природі повсюди. Класичний приклад протокооперації — взаємодія крабів і кишковопорожнинних, які, сидячи на спині краба, маскують його, оберігаючи таким чином від ворогів. Яка ж їм вигода від крабів? Передусім вони використовують крабів як транспортний засіб, а головне — їм потрапляє чимало об'їдків від столу партнера. Однак нікраб, ні кишковопорожнинні не перебувають у повній залежності один від одного.
Вважають, що на останньому етапі еволюції, який триває і сьогодні, розвивається залежність однієї популяції від іншої. Таку категорію взаємодії називають мутуалізмом. У цих умовах зв'язок популяцій сприятливий для росту і виживання обох популяцій, причому в природі жодна з них не може існувати без іншої (рис. 6). Мутуалізм ще називають облігатним (обов'язковим) симбіозом. Типові приклади мутуалізму трапляються лише у тому випадку, коли якийсь гетеротроф стає повністю залежним від автотрофа стосовно корму, а існування останнього автотрофа залежить від мінерального обміну, захисту й інших важливих функцій гетеротрофа. Такими є симбіотичні стосунки бобових рослин з азотофіксуючими бактеріями, які знаходяться в бульбочках, прикріплених до коріння рослин. Рослини постачають бактерії "збалансованою їжею", а бактерії живлять рослини азотом.

Рис. 6. Мутуалізм між Lactobacillus arabinosus 17-5 і Streptococcus faecalis R, що вирощують на синтетичному середовищі, позбавленому фенілаланіну і фолієвої кислоти:
1 — S. faecalis (штам, що потребує фолієвої кислоти);
2 — L. arabinosus (потребує фенілаланіну);
3 — обидва види при спільному культивуванні.
Поширеними є також мікоризні зв'язки, побудовані на взаємних стосунках вищої рослини і гриба. Гіфи гриба утворюють не лише чохли на дрібних всмоктуючих корінчиках рослини, але й глибоко проникають у тканину і навіть клітини коріння. Частина гіфів розгалужується в ґрунті чи мертвому відпаді. Все це сплетіння гіфів гриба є складною системою, яка поглинає неорганічні поживні речовини, щоби передати їх рослині, яка, в свою чергу, віддає певну частину поживи грибам.
Розділ 3. Середовищні зв`язки
Будь-яке угруповання можна представити у вигляді кормової мережі, яка являє собою схему всіх трофічних зв'язків між видами, що входять до складу цього угруповання. Кормова мережа звичайно складається із декількох ланцюгів живлення, кожний з яких є окремим її каналом (рис.7).
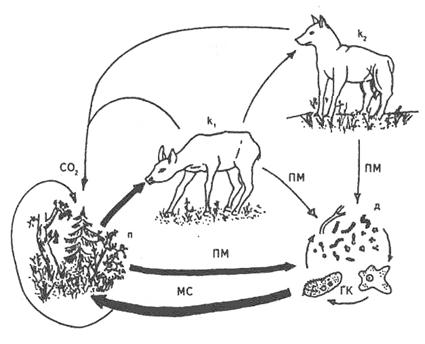
Рис. 7. Взаємозв'язок продуцентів (п) з консументами 1-го порядку (к,), консументи 2-го порядку (Ц) та деструкторами (д) і ґрунтовими консументами (ГК); ПМ — продукти метаболізму, мертвий відпад; МС — мінеральні солі
Завдяки кормовим взаємостосункам у біоценозах здійснюється трансформація біогенних речовин, акумуляція енергії і розподіл її між видами (популяціями). Чим багатший видовий склад біоценозу, тим різноманітніші напрями і швидкість потоку речовин і енергії. Ланцюги живлення, або канали живлення, якими постійно перебігає енергія, прямо чи опосередковано об'єднують всі організми в єдиний комплекс.
Перший трофічний рівень представлений первинними продуцентами, або автотрофами. До них належать зелені рослини, які здатні використовувати сонячне проміння для утворення хімічних сполук, багатих на енергію. Первинні продуценти — це найважливіша частина біоценозу, тому що практично решта організмів, що входять до його складу, прямо чи опосередковано залежать від постачання енергією, якою запаслися рослини.
Крім первинних продуцентів до складу біоценозу входять гетеротрофи (від грецьк. гетеро — інший, трофе — корм) — організми, які використовують для споживання (корму) готові органічні речовини, представлені консументами і деструкторами. Перша група утворює ланцюги поїдання, а друга — ланцюги розкладу [8].
Другий трофічний рівень утворюють травоїдні тварини, яких називають первинними консументами. М'ясоїдних, які живляться травоїдними, називають вторинними консументами, або первинними хижаками; вони перебувають на третьому трофічному рівні.
Хижаки, які живляться первинними хижаками, в свою чергу, становлять четвертий трофічний рівень і називаються третинними консументами. Тварин, що споживають вторинних хижаків, називають третинними хижаками, або ж четвертинними консументами, і т.д.
Оскільки чимало тварин всеїдні (живляться як рослинами, так і тваринами), тобто одночасно одержують енергію з декількох різних трофічних рівнів, їх неможливо віднести до відповідного рівня. Звичайно вважають, що такі організми входять відразу до декількох трофічних рівнів, а їхня участь в кожному рівні пропорційна складу вживаної ними їжі.
Кінцеву ланку кормового ланцюга утворюють так звані деструктори, або біоредуктори — організми, які розкладають органічні речовини. Це переважно мікроорганізми (бактерії, дріжджі, гриби-сапрофіти), які селяться на трупах і екскрементах і поступово їх руйнують. Завдяки їхній діяльності відбувається повернення в мінеральне царство елементів, які містяться в органічних речовинах [13].
Гриби, наприклад, в основному беруть участь у розкладі клітковини рослин, а бактерії розкладають трупи тварин. Мікроорганізми-деструктори виконують і інші функції: вони продукують інгібітори (наприклад, антибіотики) або, навпаки, речовини-стимулятори (наприклад, деякі вітаміни), екологічне значення яких дуже важливе, однак ще мало вивчене.
Серед кормових ланцюгів, які починаються з автотрофних рослин, можна передусім виділити ланцюги хижаків і ланцюги паразитів.
Кормові ланцюги хижаків ідуть від продуцентів до травоїдних, які поїдаються дрібними м'ясоїдними, останні, в свою чергу, поїдаються крупними хижаками і т.д. В міру пересування цим ланцюгом хижаків тварини в більшості випадків збільшуються розмірами і зменшуються чисельно.
Наприклад, відносно простий і короткий ланцюг має такий вигляд (рис.8.):
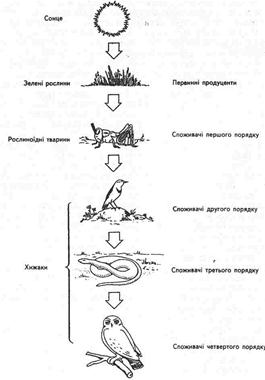
Рис. 8. Спрощений наземний кормовий ланцюг, який показує послідовність трофічних рівнів
трава → коник → просянка → вуж → сова
продуцент І консумент II консумент III консумент IV консумент; а більш довгий і складний ланцюг, який включає консументи п'ятого порядку, виглядає таким чином:
сосна звичайна → попелиця → сонечко → павуки → комахоїдні птахи → хижі птахи.
Кормові ланцюги паразитів на відміну від попередніх ведуть до організмів, які зменшуються в розмірах і збільшуються чисельно:
трава → травоїдні ссавці → блохи → Lерtотопаs.
У наведеному прикладі ссавці є місцем існування численних бліх, які живляться їх кров'ю, одночасно в організмі бліх розвиваються тисячі джгутикових одноклітинних організмів Lерtотопаs.
Кормові ланцюги можуть починатися не лише з автотрофних організмів (автотрофний тип), але і з неживих органічних речовин (детритний тип). Це трапляється тоді, коли в ролі консументів виступають детритоїдні організми, які належать до різних системних груп (безхребетні, які живуть в ґрунті, харчуючись опалим листям, або ж бактерії чи гриби, які розкладають органічні речовини) [12]. Тут спостерігається чітка узгодженість: тварини створюють умови мікроорганізмам, розділяючи трупи загиблих тварин і рослин (рис. 9).
Кормові ланцюги, які починаються з рослиноїдних і детритоїдних організмів, найчистіше представлені в екосистемах одночасно, але майже завжди один з них домінує. Однак у підземній сфері, де існування хлорофільних організмів неможливе, оскільки туди не доходить сонячне світло, існують лише ланцюги, які починаються з детритоїдних форм.
Вивчення трофічних зв'язків свідчить, що в природі жодний живий організм не може існувати ізольовано. Більше того, живі організми в межах біогеоценозів або ж біогеоценотичних комплексів створюють спеціалізовані об'єднання — консорції. Характерною особливістю цих об'єднань є те, що вони звичайно формуються на базі популяцій автотрофних рослин (сосна, смерека, береза, пшеничне чи картопляне поле тощо). Такі популяції називають детермінантами, а види, які об'єднуються довкола них, — консортами.
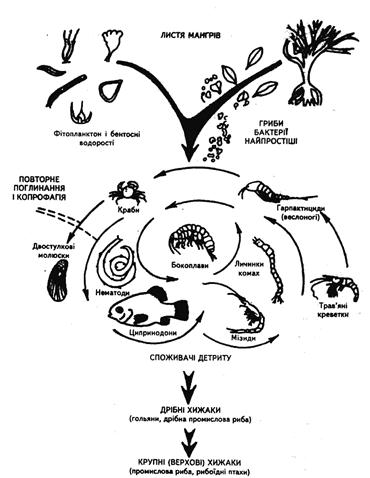
Рис. 9. Детритний харчовий ланцюг, заснований на листках мангрових дерев, що падають у мілководний естуарій на півдні Флориди
Фрагменти листя, обробленого сапротрофами і заселеного водоростями, з'їдаються одними тваринами і після проходження через їх кишковик знову з'їдаються іншими тваринами (копрофагія), які належать до ключової групи дрібних споживачів детриту: ці останні, в свою чергу, служать основною їжею промислової риби.
Слід звернути увагу на трофічні рівні кормових раціонів. Організми вважають такими, що належать до одного трофічного рівня в тому випадку, коли в кормовому ланцюгу вони віддалені від зелених продуцентів однаковим числом трофічних рівнів. Трофічний рівень і кормовий ланцюг не завжди мають струнку послідовну схему. Всеїдні тварини годуються одночасно рослинним і тваринним кормом, а деякі хижаки мають широкий набір жертв. Оскільки дана рослина чи тварина може служити кормом для різних як живоїдних, так і травоїдних, rормові ланцюги часто перетинаються один з одним, складаючи кормову мережу, або сітку (рис.10, 11). Наприклад, трава може бути спочатку спожита не кроликами, а іншими травоїдними; кролика замість лисиці може з'їсти орел і т.д.
Щоби встановити існування кормових мереж і трофічних рівнів, слід знати харчовий раціон різних видів. Для цього існують різні способи [9].
По-перше — це безпосереднє спостереження з використанням астрономічних телескопів, прихованих відеокамер тощо.
По-друге, це аналіз вмісту шлунків. Особливо цей метод використовують щодо птахів. Французький еколог Р.Дажо наводить приклад з вивченням вмісту шлунка сороки. В ньому, зокрема, виявилося 92 представники різних видів комах. У цілому в шлунку було 494 комахи, в тому числі жуків — 302, метеликів — 39, прямокрилих — 16, двокрилих — 14, коників — 10. Крім того, виявлено 112 павукоподібних, 35 ракоподібних, 4 молюски, 3 хребетні. Слід відзначити, що понад 80% — це жертви довжиною 0,6-1,5 см. Вибір проявляється також щодо біотопу: сорока в основному живилася на лужках.
А ось так виглядає раціон однієї пари яструба, кормове угіддя якої влітку досягає 3700 га, а взимку — 500,0 га (поблизу Гамбурга, Німеччина): хижаки (домашні коти, горностаї, ласки, дрібні хижі птахи) — 1,02; воронові (ворона, сорока, сойка, галка) — 7,04; дрібна дичина (заєць, кролик, куріпка, фазан, голуб) — 43,04; водоплавні птахи (мандаринки, качки, лисухи) — 1,01; домашні птахи (кури, голуби) — 20,25; різні птахи — 27,20; дрібні ссавці — 0,24. Як бачимо, склад жертв яструба різноманітний: 65 видів ссавців і птахів, причому переважають голуби, куріпки, граки, воронові і кролики. Метод аналізу вмісту шлунка має свої недоліки, оскільки у шлунках трапляються окремі м'які частинки тіла, походження яких важко встановити.
У вивченні кормових раціонів широко використовують радіоізотопний метод. Рослини , які поїдаються певною групою тварин, поливають розчином ізотопного фосфору (Р32). Розчин швидко поглинається тваринами. Через рівні відрізки часу відловлюють на досліджуваній ділянці тварин і вимірюють їх радіоактивність. Виявляється, що тварини, які активно живляться цими рослинами, тобто своїм основним раціоном, швидше набувають радіоактивності. Тварини, які живляться в основному іншими рослинами, стають радіоактивними значно пізніше — через 2-5 тижнів. У павуків вона з'являється через п'ять тижнів, а у равликів і жужелиць майже не спостерігається, тобто вони майже не вживають досліджуваних рослин.

Рис. 10. Трофічні зв'язки в простих харчових ланцюгах
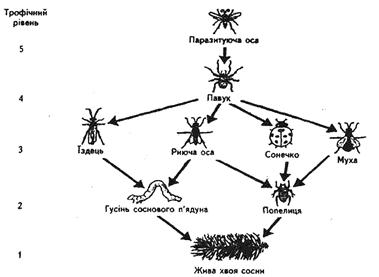
Рис. 11. Проста харчова мережа трофічних стосунків
Висновки
1. Консорція - це сукупність організмів, життєдіяльність яких у межах біогеоценозу трофічно або топічно зв'язана з центральним видом - автотрофною рослиною, у ролі якого виступає едифікатор. Популяції інших видів консорції утворять її "ядро", за рахунок якого існують види, що руйнують органічну речовину, створювана автотрофами. Інші члени консорції можуть створювати більш дрібні консорції і т.д., тобто можна виділити консорції першого, другого, третього і т.д. порядку. Звідси ясно, що біоценоз - це система зв'язаних між собою консорцій. Прикладом консорції може служити рослина з усіма зв'язаними з ним організмами (епіфіти, паразити, шкідники, мутуалісти).
2. Для консорції характерні біотичні взаємодії між організмами: паразитизм, алелопатія (або антибіоз), позитивна взаємодія (коменсалізм, протокооперація, мутуалізм).
3. В консорції відбуваються трофічні взаємодії між організмами, характерні трофічні зв`язки. Оскільки певна рослина чи тварина може служити кормом для різних як живоїдних, так і травоїдних, кормові ланцюги часто перетинаються один з одним, складаючи кормову мережу, або сітку.
Список використаної літератури
1. Білявський Г.О. та інші. Основи екологічних знань: Навч. посібник. – К.: Либідь, 2002. – 346 с.
2. Бойчук Ю.Д., Солошенко Е.М., Бугай О.В. Екологія і охорона навколишнього природного середовища: навчальний посібник. – Суми, 2002. – 284 с.
3. Гайнріх Д., Гергт М. Екологія. - К.: Знання-прес, 2001. – 287 с.
4. Голубець М.А., Кучерявий В.П., Генсірук С.А. та ін. Конспект лекцій з курсу "Екологія і охорона природи". - К., 1990.
5. Заверуха Н.М., Серебряков В.В., Скиба Ю.А. Основи екології. – К.: Каравела, 2006. - 368 с.
6. Запольський А.К., Салюк А.І. Основи екології. – К.: Вища школа, 2003. – 358 с.
7. Злобін Ю.А., Кочубей Н.В. Загальна екологія: Навч.посібник. – Суми: ВТД «Університетська книга», 2004. – 416 с.
8. Кучерявий В.П. Екологія. – Львів: Світ, 2001. – 500 с.
9. Новиков Г.А. Основи общей экологии. Л., 1979. – 286 с.
10. Новиков Ю.В. Природа и человек. – М.: Просвещение, 1991. – 223 с.
11. Новиков Ю.В. Экология, окружающая среда и человек. - М.: ФАИР, 1998. - 320 с.
12. Одум Ю. Экология. В 2 т. - М., 1986., Т 1. – 328 с., Т.2. – 376 с.
13. Одум Ю. Экология. В 2 т. - М., 1986., Т.2. – 376 с.
0 комментариев