МІНІСТЕРСТВО ОХОРОНИ ЗДОРОВ’Я УКРАЇНИ
ЛЬВІВСЬКИЙ НАЦІОНАЛЬНИЙ МЕДИЧНИЙ УНІВЕРСИТЕТ
ІМЕНІ ДАНИЛА ГАЛИЦЬКОГО
КОНОВАРТ ОКСАНА ВІКТОРІВНА
УДК 612.112.94.015.11.014.23.014.333
МЕХАНІЗМИ ДІЇ БЛОКАТОРІВ Н2-ГІСТАМІНОВИХ І
М1-ХОЛІНЕРГІЧНИХ РЕЦЕПТОРІВ У ЛІМФОЦИТАХ
ПЕРИФЕРИЧНОЇ КРОВІ
14.03.03 – нормальна фізіологія
Автореферат
дисертації на здобуття наукового ступеня
кандидата медичних наук
Львів – 2008
Дисертацією є рукопис.
Робота виконана у Львівському національному медичному університеті імені Данила Галицького МОЗ України
Науковий керівник
доктор біологічних наук, професор, Воробець Зіновій Дмитрович, Львівський національний медичний університет імені Данила Галицького МОЗ України, завідувач кафедри медичної біології та генетики
Офіційні опоненти:
доктор медичних наук, професор Скляров Олександр Якович,
Львівський національний медичний університет імені Данила Галицького МОЗ України, завідувач кафедри біологічної хімії
доктор біологічних наук, професор Берегова Тетяна Володимирівна, Науково-дослідний інститут фізіології імені академіка Петра Богача біологічного факультету Київського національного університету імені Тараса Шевченка МОН України, завідувач відділу фармако-фізіології
Захист відбудеться „16” травня 2008 р. об 11 годині на засіданні спеціалізованої вченої ради Д 35.600.03 Львівського національного медичного університету імені Данила Галицького МОЗ України за адресою: 79010, м. Львів, вул. Пекарська, 52.
З дисертацією можна ознайомитись у Науковій бібліотеці Львівського національного медичного університету імені Данила Галицького (79010, м. Львів, вул. Січових Стрільців, 6).
Автореферат розісланий „ 7 ” квітня 2008 р.
Вчений секретар
спеціалізованої вченої ради Томашова С.А.
ЗАГАЛЬНА ХАРАКТЕРИСТИКА РОБОТИ
Актуальність теми. Розвиток багатьох захворювань супроводжується оксидативним стресом внаслідок інтенсивного утворення у клітинах активних форм кисню (АФК) (Барабой В.А., Сутковой Д.А., 1997; Владимиров Ю.А., 1989). Завдяки високій реакційній здатності, АФК можуть призводити до ураження клітин, викликаючи окиснення біомолекул та ініціювання ланцюгових процесів пероксидного окиснення у мембранних ліпідах. Зміна ліпідного оточення мембран та посилення процесів пероксидації призводять до структурних порушень у клітинах і модуляції активності мембранозв’язаних ферментів, таких як АТФаза, аденілатциклаза тощо (Барабой В.А., Сутковой Д.А., 1997; Krinsky N.I., 1991). Недостатнє функціонування систем антиоксидантного захисту спричиняє порушення активності Са2+-транспортувальних систем і, відповідно, призводить до зміни концентрації іонізованого кальцію у клітині, який є внутрішньоклітинним месенджером і прямо чи опосередковано регулює більшість клітинних функцій. Каталітична активність ферментів антиоксидантної системи (АОС), яка регулює процеси вільнорадикального окиснення у клітинах, забезпечує підтримання стабільності їх плазматичних мембран і внутрішньоклітинних структур в умовах оксидативного стресу (Krinsky N.I., 1991; Ursini F., Bindoli A., 1987). Важлива роль у цих процесах належить ферментам глутатіонової антиоксидантної системи.
Відомо, що гострі та хронічні панкреатити, підвищена шлункова секреція супроводжуються оксидативним стресом (Білоусов Ю.Б., Асецкая И.Л., 1993; Дегтярьова І.І. та ін., 2000; Полотнюк С.Я., Штанова Л.Я., Берегова Т.В., 2006). Для їх лікування широко використовуються блокатори Н2-гістамінових і М1-холінергічних рецепторів (Дегтярьова І.І. та ін., 2000; Gespach C., 1990; Kulkarni P. N., 1997; Leurs R. et all., 1991; Singer M. V. et all., 1991; Sklyarov A. Y. et all., 2001; 2002). Для зниження гіперпродукції хлоридної кислоти парієтальними клітинами слизової оболонки шлунку (СОШ) застосовують блокатори протонної помпи (Burdan F. et all., 2000). Відомо, що блокатор М1-холінергічних рецепторів пірензепін, на відміну від неселективних середників, вибірково пригнічує секрецію кислоти та пепсину у СОШ, зменшує функціональну активність ацинарних клітин підшлункової залози, покращує мікроциркуляцію та кровопостачання. Селективний блокатор Н2-гістамінових рецепторів – фамотидин – не тільки гальмує секрецію шлункових залоз, але й інгібує генерацію АФК, активність аденілатциклази, синтез панкреатичних ферментів.
Хоча Н2 - і М1-рецептори експресуються переважно у парієтальних клітинах СОШ та ацинарних клітинах підшлункової залози, вони також наявні і у лімфоцитах (Ganther F. et all., 2002; Jutel M. et all., 2001; Nomura J. et all., 2003).
Адаптація організму до стресових чинників та підтримання гомеостазу здійснюється за участю нервової, ендокринної та імунної систем (Давтян Т.К., Аванесян Л.А., 2001). Виявлена функціональна та фенотипова подібність між нервовою та імунною системами. Лімфоїдні та нервові клітини здатні синтезувати одні і ті ж біологічно активні речовини, експресувати одні і ті ж рецептори, зокрема М1 і Н2. Це обумовлює те, що лімфоцити, як й нервові клітини, можуть активно і оперативно брати участь в індукції та регуляції стрес-реакції організму шляхом синтезу і секреції різноманітних факторів (Давтян Т.К., Аванесян Л.А., 2001). Виходячи з цього, можна припустити, що лімфоцити крові можуть бути зручною, адекватною та актуальною моделлю для вивчення багатьох процесів, зокрема механізмів дії Н2 - і М1 - антагоністів. Стимуляція В - і Т-лімфоцитів супроводжується каскадом біохімічних реакцій і різноманітних шляхів мобілізації Са2+ (Choquet D. et all., 1994). Са2+ задіяний у всіх регуляторних системах та у формуванні клітинної відповіді на зовнішні впливи (Костюк П.Г., Чазов Е.И., 1988; Choquet D. et all., 1994; Gallo E. M. et all., 2006). Низькі концентрації іонізованого кальцію у клітині (10-6-10-7 М) підтримуються завдяки скоординованій роботі Са2+-транспортувальних та інших іон-транспортувальних систем, зокрема Са2+, Мg2+-АТФази та Nа+, К+-АТФази. Отже, для розкриття механізму дії блокаторів Н2 - та М1 - рецепторів і протонної помпи при ряді захворювань травної системи доцільно вивчати на модельних системах, зокрема лімфоцитах периферичної крові, їх ефект на іон-транспортувальні системи і на системи антиоксидантного захисту.
Зв’язок роботи з науковими програмами, планами, темами. Дисертаційна робота виконана відповідно до плану науково-дослідних робіт Львівського національного медичного університету імені Данила Галицького у рамках теми „Вивчення регуляторної ролі кальцію у функціонуванні глутатіонової антиоксидантної системи клітин” (№ реєстрації ІН.07.00.0001.04). Тема дисертації затверджена на засіданні Вченої Ради медичного факультету № 2 Львiвського національного медичного унiверсистету iмені Данила Галицького, протокол № 3 від 21 грудня 2005 р., на засіданні проблемної комісії з медико-біологічних дисциплін Львівського національного медичного університету імені Данила Галицького, протокол № 1 від 7 грудня 2005 р. та на засіданні Проблемної комісії "Фізіологія людини" АМН та МОЗ України, протокол № 1 від 15 лютого 2007 р.
Мета роботи: з’ясувати роль глутатіонової антиоксидантної системи і транспортувальних АТФаз у реалізації ефектів блокаторів Н2-гістамінових та М1-холінергічних рецепторів, а також блокатора протонної помпи у лімфоцитах периферичної крові.
Завдання дослідження:
1. Встановити оптимальні умови для функціонування глутатіонпероксидазної, глутатіонтрансферазної, глутатіонредуктазної, Na+,K+-АТФазної та Са2+,Mg2+-АТФазної активностей лімфоцитів периферичної крові, використовуючи як пермеабілізуючий агент сапонін.
2. Вивчити вплив блокатора Н2-гістамінових рецепторів фамотидину на активність ферментів глутатіонової антиоксидантної системи та активність транспортувальних АТФаз у лімфоцитах периферичної крові.
3. Вивчити вплив блокатора М1-ацетилхолінових рецепторів пірензепіну на активність ферментів глутатіонової антиоксидантної системи та активність транспортувальних АТФаз у лімфоцитах периферичної крові.
4. Дослідити вплив омепразолу як блокатора протонної помпи на активність ферментів глутатіонової антиоксидантної системи та активність транспортувальних АТФаз у лімфоцитах периферичної крові.
Об’єкт дослідження: механізми дії ферментів глутатіонової антиоксидантної системи і транспортувальних АТФаз в опосередкуванні ефектів блокаторів Н2-гістамінових та М1-холінергічних рецепторів, а також блокатора протонної помпи у лімфоцитах периферичної крові.
Предмет дослідження: ферменти глутатіонової антиоксидантної системи – глутатіонпероксидаза, глутатіонредуктаза та глутатіонтрансфераза, Na+,K+-АТФаза та Ca2+,Mg2+-АТФаза у лімфоцитах крові при дії фамотидину, пірензепіну та омепразолу.
Методи дослідження: виділення моноядерних лімфоцитів з периферичної крові практично здорових донорів у градієнті густини фікол-урографіну, біохімічні методи (пермеабілізація клітин сапоніном, визначення концентрації білка за Лоурі, визначення активностей глутатіонпероксидази, глутатіонредуктази, глутатіонтрансферази, Са2+, Мg2+ - та Nа+, К+-АТФаз, оцінка стану пероксидації ліпідів, визначення вмісту неорганічного фосфату), статистично-математичне опрацювання результатів за допомогою t-критерію Стьюдента.
Наукова новизна одержаних результатів. Уперше комплексно вивчено функціонування окремих ланок глутатіонової антиоксидантної системи, транспортувальних АТФаз та їх чутливість до іонів кальцію у моноядерних лімфоцитах периферичної крові. Встановлені оптимальні концентрації сапоніну, які розкривають латентні активності глутатіонпероксидази, глутатіонредуктази, глутатіонтрансферази, Са2+, Мg2+ - та Nа+, К+-АТФаз у лімфоцитах периферичної крові, що дає змогу виявити функціональні зв’язки між цими ферментами. Підібрані оптимальні умови для вивчення активностей цих ферментів у пермеабілізованих лімфоцитах.
З’ясовано, що блокатор Н2-гістамінових рецепторів фамотидин дозозалежно впливає на активність глутатіонпероксидази та глутатіонредуктази – при низьких концентраціях активує ці ферменти, а при високих (>10-3 М) інгібує. Водночас цей препарат у концентраціях >10-4 М практично повністю інгібує Са2+, Мg2+ - та Nа+, К+-АТФазні активності лімфоцитів периферичної крові.
Блокатор М1-ацетилхолінових рецепторів пірензепін інгібує пероксидацію ліпідів та стимулює активність глутатіонпероксидази, а також, цей же блокатор дозозалежно інгібує активність Са2+, Мg2+ - та Nа+, К+-АТФаз.
Блокатор протонної помпи омепразол у концентраціях до 10-4 М стимулює активність глутатіонпероксидази та глутатіонредуктази, а у вищих концентраціях інгібує їх. Омепразол також, подібно до фамотидину та пірензепіну, дозозалежно інгібує активність Са2+, Мg2+ - та Nа+, К+-АТФаз.
Практичне значення одержаних результатів. Зміна активності іон-транспортувальних систем і ферментів антиоксидантної системи при дії різних блокаторів може супроводжуватись модуляцією синтезу лімфоцитами біологічно активних речовин і здатністю клітин до хемотаксису. Оскільки Н2-гістамінові та М1-ацетилхолінові рецептори широко експресуються не тільки у клітинах СОШ чи ацинарних клітинах підшлункової залози, але й у лімфоцитах периферичної крові, ці клітини можуть бути зручною моделлю для вивчення механізмів дії блокаторів вказаних рецепторів, які використовуються для лікування кислотозалежних захворювань і панкреатитів. Отримані дані можуть слугувати основою для розробки методів корекції патологічних змін клітинного гомеостазу. З огляду на це можна ставити питання про створення та підбір нових фармакологічних препаратів, які модулюють активність глутатіонової антиоксидантної системи і транспортувальних АТФаз.
Результати дисертаційної роботи впроваджені у навчальний процес кафедр медичної біології та генетики, нормальної фізіології і біологічної хімії Львівського національного медичного університету імені Данила Галицького.
Особистий внесок здобувача. Дисертант самостійно проаналізувала наукову літературу за темою дослідження, сформулювала та обґрунтувала основні положення дисертаційної роботи і налагодила методи досліджень. Експериментальні дослідження, результати яких викладені у дисертації, здобувач проводила особисто. Постановка завдань, аналіз та обговорення отриманих результатів здійснено спільно з науковим керівником і співавторами статей.
Апробація результатів дисертації. Апробація дисертації проведена на спільному засіданні кафедр нормальної фізіології і медичної біології та генетики Львівського національного медичного університету імені Данила Галицького 20 вересня 2007 року, протокол № 3. Основні положення дисертаційної роботи викладені у доповідях та обговорені на 63-й науковій конференції студентів і молодих вчених Львівського національного медичного університету імені Данила Галицького (Львів, 2003), Міжнародних наукових конференціях „Нейрогуморальні та клітинні регуляторні механізми процесів травлення” (Львів, 2003, 2007), Міжнародній науковій конференції „Клітинні та субклітинні механізми функціонування травної системи” (Львів, 2004), Установчому з’їзді товариства клітинної біології (Львів, 2004), ІV Львівсько-Люблінській конференції з експериментальної та клінічної біохімії (Люблін, 2006); ІХ Українському біохімічному з’їзді (Харків, 2006), Міжнародній науковій конференції „Механізми функціонування фізіологічних систем” (Львів, 2006).
Публікації. Основні результати дисертаційної роботи висвітлені у 13 наукових працях: 7 статей (5 у наукових фахових виданнях, рекомендованих ВАК України, 2 – у зарубіжних часописах), 6 робіт у матеріалах конференцій та з’їздів.
Структура та обсяг дисертації. Дисертація викладена на 131 сторінках, побудована за традиційною схемою та складається зі вступу, огляду літератури, опису матеріалів і методів досліджень, 4-х розділів, де викладено отримані результати, проведено їх аналіз та узагальнення, а також із висновків і списку використаної літератури. Робота містить 25 рисунків і 4 таблиці. Бібліографічний список налічує 232 джерела.
ОСНОВНИЙ ЗМІСТ РОБОТИМатеріали та методи дослідження. Моноядерні лімфоцити периферичної крові людини виділяли з гепаринізованої свіжоотриманої крові практично здорових донорів віком 21-28 років у градієнті концентрації фікол-урографіну (480 зразків) (A. Boum, 1968). Підраховували клітини у камері Горяєва, використовуючи як барвник 0,1% трипановий синій. Життєздатність лімфоцитів, яка в усіх дослідах складала не менше 95%, оцінювали за забарвленням трипановим синім.
Для розкриття глутатіонредуктазної, глутатіонтрансферазної та Nа+, К+-АТФазної латентної активностей до суспензії лімфоцитів додавали 0,2% сапонін, а для визначення глутатіонпероксидазної та Са2+, Мg2+-АТФазної активностей - 0,1% сапонін. Дана методика грунтується на роботах, виконаних на еритроцитах, лімфоцитах і сперматозоїдах (Орлов С.Н. и др., 1985; Підковка Н.О., 2003; Maksymjuk H. V., Vorobets Z. D., 2000).
Інтенсивність пероксидної оксидації ліпідів (ПОЛ) оцінювали за вмістом одного із кінцевих метаболітів – малонового діальдегіду (Тимирбулатов С.А., Селезнев Е.И., 1988).
Глутатіонпероксидазну активність визначали за зменшенням вмісту GSH (Моин В.М., 1986), глутатіонредуктазну активність – за зменшенням вмісту NADPH (Mannervik V., 1991), глутатіонтрансферазну активність – за зменшенням вмісту GSH (Власова С. Н и др., 1990; Булавин Д.В., 1996).
Визначення Ca2+,Mg2+-АТФазної активності проводили при 37° С у середовищі, що містило 5 мМ Nа2АТФ, 5 мМ МgС12, 5 мкМ СаС12, 0,02 М трис-НCl буфер (рН=7,4). До інкубаційного середовища додавали лімфоцитарну суміш. Реакцію зупиняли додаванням 1 мл 20% ТХО. Зразки 10 хв центрифугували при 800 g. У супернатанті визначали вміст неорганічного фосфату за методом Фіске-Субароу (Меншиков В.В., 1987). Активність Са2+, Мg2+-АТФази оцінювали за величиною, що інгібувалась 1 мМ розчином ЕГТА, та відображали у мкмолях Фн (хв · мг білка) - 1. Мg2+-АТФазну активність визначали у тих же умовах, але за відсутності СаС12 із додаванням 1 мМ ЕГТА та 0,1 мМ оуабаїну (Орлов С.Н., 1977). За Na+,K+-ATФазну активність приймали таку, що інгібувалась 0,1 мМ розчином оуабаїну.
Препарати фамотидину, пірензепіну, омепразолу використовували в концентраціях 10-6...10-3 М.
Варіаційно-статистичне опрацювання отриманих результатів проводили з використанням критерію Стьюдента за допомогою комп’ютерної програми Microsoft Excel 2003.
РЕЗУЛЬТАТИ ДОСЛІДЖЕНЬ ТА ЇХ ОБГОВОРЕННЯФункціональні властивості глутатіонзалежних ферментів і транспортувальних АТФаз лімфоцитів крові при пермеабілізації клітин сапоніном. Відомо, що очищені ферменти глутатіонової антиоксидантної системи не є Ca2+-залежними (Барабой В.А., Сутковой Д.А., 1997; Doroshow J. H., 1995; Mannervik B., 1991). Однак, невідомо, як вони функціонують in vivo. Значна кількість даних вказує на те, що на активність цих ферментів, якщо не прямо, то опосередковано, впливає іонізований кальцій (Підковка Н.О., Воробець З.Д., 2002).
Враховуючи те, що ряд ферментів, які ми вивчали, локалізовані з внутрішнього боку плазматичної мембрани (АТФази, глутатіонпероксидаза - ГП, глутатіонтрансфераза - ГТ), інші - у цитозольній фракції (глутатіонредуктаза - ГР), для розкриття їх латентної активності доцільною є пермеабілізація клітинних мембран.
Згідно завдань досліджень, був проведений підбір оптимальних умов для визначення ГР-, ГП - та ГТ-ої активностей лімфоцитів периферичної крові, з використанням як пермеабілізуючого агента сапоніну. Для цього в інкубаційне середовище для визначення ферментативної активності додавали сапонін у діапазоні концентрацій 0,02...0,3%. ГП-а активність практично не змінювалась при низьких концентраціях сапоніну (до 0,04%), вона становила 9,2±0,9 нмоль GSН (хв · мг білка) - 1. Цей показник значно зростав, до 75,0±6,8 нмоль GSН (хв · мг білка) - 1 (р < 0,05), при вмісті сапоніну 0,1%. За наявності в інкубаційному середовищі вищих концентрацій (0,2%) цієї сполуки ГП-а активність різко знижувалась.
Встановлено, що ГР-а активність лінійно зростає від 4,2±0,3 нмоль NADРН (хв · мг білка) - 1, за відсутності сапоніну, до 30,0±3,1 нмоль NADРН (хв · мг білка) - 1, із підвищенням концентрації сапоніну до 0,2%.
У діапазоні концентрацій сапоніну 0...0,04% спостерігались дуже низькі значення ГТ-ої активності. При 0,04% вмісті сапоніну в інкубаційному середовищі ферментативна активність становила 3,2±0,3 нмоль GSН (хв · мг білка) - 1. За більших концентрацій (0,1-0,2%) сапоніну було виявлено лінійне зростання ГТ-ої активності до 68,1±6,7 нмоль GSН (хв · мг білка) - 1 (р < 0,05).
Таким чином, отримані результати довели необхідність підбору оптимальних концентрацій сапоніну для визначення активності глутатіонових антипероксидних ферментів нативних лімфоцитів периферичної крові.
При визначенні Са2+, Мg2+-АТФазної активності у діапазоні концентрацій сапоніну 0,02...0,1% спостерігалось зростання показника до 7,9±1,3 мкмоль Фн (хв · мг білка) - 1. Більші концентрації сапоніну пригнічували активність даного ферменту.
Виявлено, що Na+, К+-АТФазна активність зростає лінійно відповідно до підвищення концентрації сапоніну в інкубаційному середовищі. Максимальна активність даного ферменту становила 10,6±0,6 мкмоль Фн (хв · мг білка) - 1, за 0,2% концентрації сапоніну.
Отже, низькі концентрації сапоніну (0,02...0,04%) є недостатніми для розкриття латентної активності досліджуваних ферментів. Оскільки це є здебільшого мембранозв’язані або цитозольні ферменти, а ГР - виключно цитозольний, для їх активації необхідна більша концентрація сапоніну - 0,1% для ГП, Са2+, Мg2+-АТФази та 0,2% для ГР, ГТ, Na+, К+-АТФази.
Оскільки функціонування будь-яких ферментів залежить від багатьох факторів, завданням наступного етапу роботи був добір оптимальних значень таких показників, як рН, концентрація білка, АТФ, співвідношення катіонів Na+ і К+, час інкубації для визначення Са2+, Мg2+-АТФазної та Na+, К+-АТФазної активностей лімфоцитів крові.
Було показано, що оптимум рН для Са2+, Мg2+-АТФази становить 7,0. За цих умов спостерігалась максимальна активність ферменту - 16±1,4 мколь Фн (хв · мг білка) - 1. Na+, К+-АТФазна активність сягала максимального значення при рН 6,8 і становила 10±0,7 мкмоль Фн (хв · мг білка) - 1. Зниження рН інкубаційного середовища від 7,4 до 6,0 призводило до істотного посилення інгібувального впливу Са2+ на АТФ-гідролазну активність ферменту. Зміщення рН у лужний бік, зокрема до 8,0, зменшувало інгібувальний ефект катіонів кальцію на активність Na+, К+-АТФази.
У результаті вивчення процесу гідролізу АТФ Са2+, Мg2+-АТФазою лімфоцитів залежно від часу виявлено, що активність цього ферменту становить 2,0±0,1 мкмоль Фн (хв · мг білка) - 1, за 20 хв інкубації досягає максимального значення 4,5±0,3 мкмоль Фн (хв · мг білка) - 1, а при збільшенні часу інкубації виходить на плато.
Регуляторна роль іонів кальцію у пероксидації ліпідів і функціонуванні антиоксидантної системи. Для визначення залежності Са2+, Мg2+-АТФазної, ГП-, ГР-, ГТ-ої активностей від концентрації Са2+, його вносили у вигляді СаСl2 в інкубаційне середовище у концентраціях 0,01 мМ...2 мМ. Як контроль використовували проби без додавання Са2+.
Інтенсивність ПОЛ оцінювали за вмістом одного з кінцевих метаболітів - малонового діальдегіду (МДА). Згідно з результатами досліджень, вміст МДА зростав від 228,3±12,5 нмоль/мг білка, у контрольних пробах, до 365,2±18,1 нмоль/мг білка, за наявності 0,5 мМ Са2+.
ГП-а активність лінійно зростала із підвищенням концентрації Са2+ до 0,01 мМ і сягала 340,1±18,5 нмоль GSН (хв · мг білка) - 1 (рис.1).
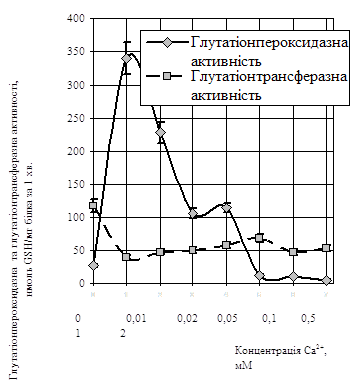
Рис.1. Вплив різних концентрацій Са2+ на глутатіонпероксидазну і глутатіонтрансферазну активності у лімфоцитах периферичної крові (М±m, n=15).
Більші концентрації Са2+ (0,05 мМ) зумовлювали зниження ГП-ої активності до 120,0±10,0 нмоль GSН (хв · мг білка) - 1. За наявності 0,1 мМ Са2+ у середовищі інкубації спостерігався незначний пік зростання ГП-ої активності до 134,5±10,4 нмоль GSH (хв мг білка) - 1. Більші концентрації Са2+ спричиняли інгібування активності даного ферменту.
Додавання Са2+ у середовище інкубації пригнічувало ГТ-у активність. Цей показник був найбільшим за відсутності Са2+, тобто у контрольних зразках, де він становив 117,1±10,1 нмоль GSH (хв · мг білка) - 1. Концентрації Са2+ 0,1...2 мМ зумовлювали зниження активності ферменту до 46,4±3,5 нмоль GSH (хв · мг білка) - 1 (р < 0,05) (рис.1).
ГР-а активність при 0,01 мМ Са2+ виявляла тенденцію до зниження, порівняно з контролем 24,9±3,1 нмоль NADРН (хв · мг білка) - 1, до 20,0±1,6 нмоль NADРН (хв мг білка) - 1. Із підвищенням концентрації Са2+ до 0,1 мМ спостерігалось зростання ГР-ої активності до 60,1±6,6 нмоль NADРН (хв · мг білка) - 1. Подальше збільшення вмісту Са2+ призводило до зниження активності даного ферменту.
Виявлену залежність активності ферментів глутатіонової АОС від вмісту Са2+ у середовищі можна пояснити з одного боку зростанням інтенсивності ПОЛ, а з другого - активацією власне Са2+-транспортувальних систем.
Отже, катіони Са2+ здатні регулювати процес ПОЛ та функціонування системи знешкодження наслідків його дії, здійснюючи не тільки стимулюючий вплив на глутатіонову АОС при низьких концентраціях (0,01 мМ...0,1 мМ), а й інгібувальний вплив при вищих концентраціях Са2+.
У результаті вивчення процесу гідролізу АТФ Са2+, Мg2+-АТФазою лімфоцитів було показано, що активність цього ферменту лінійно зростає із підвищенням концентрації Са2+ до 0,1 мМ і сягає 2,41±0,11 мкмоль Фн (хв · мг білка) - 1. Стосовно контролю активність ферменту зростала у 3,5 рази. Більші концентрації Са2+ призводили до зниження Са2+, Мg2+-АТФазної активності.
Продемонстрований нами вплив Са2+ на перебіг процесів ПОЛ та функціонування ферментів глутатіонової системи, ймовірно, полягає у збільшенні проникності плазматичної мембрани лімфоцитів для Са2+ внаслідок модифікуючого впливу ПОЛ, індукованого мілімолярними концентраціями власне Са2+. Підвищення в'язкості клітинних мембран внаслідок ПОЛ призводить до пригнічення роботи ферментативних систем, що виводять Са2+ з клітин, а отже концентрація Са2+ у клітині зростає та розвивається оксидативний стрес.
Зміни властивостей глутатіонової антиоксидантної системи і транспортувальних АТФаз при дії фамотидину. Відомо, що одним з основних „пускових механізмів” розвитку виразки шлунку, панкреатитів є вплив стресових чинників на організм (Циммерман Я.С., 2000).
На даний час накопичилось багато даних про інтенсифікацію процесів ПОЛ у патогенезі виразок різного походження. Посилення процесів вільнорадикального окиснення у мембранах клітин СОШ призводить до пошкодження їх структури. У процесі пероксидації ліпідів вільні радикали ушкоджують основні біомолекули (нуклеїнові кислоти, білки, ліпіди, вуглеводи) (Sohal R. S. et all., 1989).
Парієтальні клітини СОШ продукують хлоридну кислоту, яка відноситься до агресивних факторів, тому дослідження стану вільнорадикального окиснення у цих клітинах є досить актуальним. Одним із основних регуляторів функціонування парієтальних клітин є гістамін, що реалізує свою дію через Н2-рецептори. Однак, у лімфоцитах теж експресуються гістамінові рецептори. Тому можна припустити, що з метою вивчення механізму дії агоністів чи антагоністів гістамінових рецепторів можна використовувати лімфоцити периферичної крові.
У результаті проведених досліджень було виявлено, що гістамін активує ПОЛ. Вміст МДА при цьому зростає на 16%, з 226,4 ± 20,3 до 262,2 ± 22,1 нмоль/мг білка, а вміст МДА, індукованого адріаміцином, зростає на 31%, з 343,8 ± 29, 4 до 449, 5 ± 38,9 нмоль/мг білка.
У лікуванні захворювань травної системи широко використовується фамотидин - один з найефективніших представників селективних блокаторів Н2-гістамінових рецепторів (Н2-ГР) (Хомерики С.Г., Хомерики Н.М., 2000). Фамотидин дещо знижував вміст МДА у лімфоцитах стосовно контролю, з 226,4 ± 20,3 до 191,3 ± 17,7 нмоль/мг білка. Індукований адріаміцином вміст МДА також при цьому знижувався, з 343,8 ± 29,4 до 264,2 ± 18,1 нмоль/мг білка (р < 0,05).
Отримані нами результати стосовно впливу гістаміну та фамотидину на процеси ПОЛ у лімфоцитах узгоджуються з даними, отриманими на клітинах СОШ (Bliss P. W. et all., 2001; Hung C. R., 2000; Velinov H. et all., 2001).
Результати проведених досліджень свідчать про те, що антагоніст Н2-ГР фамотидин дещо знижує рівень ПОЛ у лімфоцитах периферичної крові. Таким чином, можна зробити висновок про залучення Н2-ГР у регулювання процесів вільнорадикального окиснення. Активування Н2-ГР призводить до зростання вмісту ПОЛ, а антагоніст Н2-ГР фамотидин знижує рівень ПОЛ.
Тоді як гістамін найбільш виразно стимулює продукцію НCl парієтальними клітинами шлунку, Н2-антагоністи інгібують кислотну секрецію. Антисекреторний ефект Н2-антагоністів дозволяє застосовувати їх для лікування захворювань шлунково-кишкового тракту, причиною яких є підвищене утворення НCl та пепсину (Хомерики С.Г., Хомерики Н.М., 2000; Cooke H. J. et all., 1995).
Імовірно, що моноядерні лімфоцити периферичної крові можуть бути моделлю для вивчення механізму дії Н2-антагоністів. Оскільки каталітична активність ферментів глутатіонової АОС регулює рівень процесів вільнорадикального окиснення у клітинах, забезпечує підтримання стабільності плазматичних мембран і внутрішньоклітинних структур за умов окиснювального стресу, одним із наслідків якого є порушення Са2+ транспорту, а Са2+, Мg2+ - та Nа+, К+-АТФази прямо і посередньо беруть участь у підтриманні гомеостазу цього катіона у клітині (Костерін С.О., 2000; Carafoli E., 1991), метою даної частини роботи було дослідити вплив різних концентрацій фамотидину на активність цих ферментів у лімфоцитах крові.
Встановлено, що низькі концентрації фамотидину (10-7-10-5 М) практично не впливають на ГП-у активність, тоді як при його вмісті 10-4 М, ферментативна активність зростає до 150,6±12,5 нмоль GSН (хв · мг білка) - 1, у контрольних зразках ГП-а активність становила 119±12,5 нмоль GSН (хв · мг білка) - 1 (р < 0,05). Підвищення концентрації фамотидину до 10-3 М спричиняє зниження ГП-ої активності до 75,0±6,5 нмоль GSН (хв · мг білка) - 1.
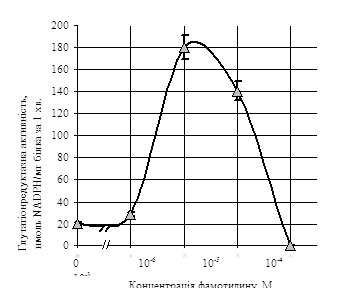 |
ГР-а активність зростала від 20,7±1,9 нмоль NADPН (хв · мг білка) - 1, за відсутності фамотидину, до 179,5±12,9 нмоль NADPН (хв · мг білка) - 1, при його концентрації 10-5 М (р < 0,05). Низькі концентрації препарату (до 10-6 М) практично не впливали на ГР-у активність. Фамотидин у концентраціях більших, ніж 10-5 М обумовлював поступове дозозалежне зниження ГР-ої активності до нульових значень (рис.2).
Рис.2. Вплив різних концентрацій фамотидину (квамателу) на глутатіонредуктазну активність лімфоцитів (М±m, n=15).
При вивченні залежності Са2+, Мg2+-АТФазної активності лімфоцитів крові людини від концентрацій фамотидину виявлено, що у контрольних пробах вона становить 1,1±0,5 мкмоль Фн (хв · мг білка) - 1. При підвищенні концентрації фамотидину в інкубаційному середовищі до 10-5 М, Са2+, Мg2+-АТФазна активність зменшувалась у 10 разів, до 0,1±0,05 мкмоль Фн (хв · мг білка) - 1. Вищі концентрації фамотидину повністю пригнічували активність даної АТФази (рис.3).
При 10-5 М концентрації фамотидину, Nа+, К+-АТФазна активність, яка у контрольних пробах становила 8,0±0,6 мкмоль Фн (хв · мг білка) - 1, пригнічувалась повністю, на відміну від Са2+, Мg2+-АТФазної активності. Нижча концентрація антагоніста (10-6 М) пригнічувала Na+, К+-АТФазну активність у 1,3 рази (р < 0,05). Активність цього ферменту залежить від ліпідного оточення, яке підлягає модифікаціям процесами ПОЛ.
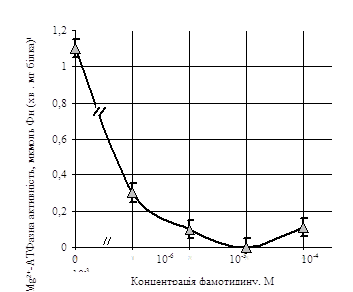
Рис.3. Залежність Са2+, Мg2+-АТФазної активності лімфоцитів від концентрації фамотидину (М±m, n=12).
Са2+, Мg2+-АТФази у клітинах слугують не лише одним із шляхів випомповування надлишку Са2+ із цитозоля, приймають участь у підтримуванні його низького рівня та залучені у сигнальні шляхи, але є одними із маркерів за умов патології. Відомо, що зростання концентрації іонізованого Са у цитоплазмі є одним із факторів, що беруть участь у стресових ураженнях тканин, а також при некрозних змінах на пізніх етапах розвитку виразкової хвороби. Отримані також дані про важливу роль [Са2+] і, як регулятора протонної помпи на ранніх етапах захворювання (Ashby M. S., Tepikin A. V., 2002). Тому визначення активностей АТФаз є важливим для вивчення подальших шляхів трансдукції сигналу гістамінових рецепторів.
Таким чином, нами встановлено, що Н2-ГР залучені у регулювання активності ферментів глутатіонової АОС і транспортувальних АТФаз. Встановлено антиоксидантні властивості фамотидину та продемонстровано його стимулюючий вплив на ГП - та ГР-у активності. Для АТФазних активностей засвідчено інгібувальний вплив препарату. Виявлені зміни активності Nа+, К+ - та Са2+, Мg2+-АТФаз можна пояснити як зміною ліпідного оточення, так і змінами інтенсивності ПОЛ.
Вплив блокатора М1-холінергічних рецепторів пірензепіну на активність ферментів глутатіонової антиоксидантної системи і транспортувальних АТФаз лімфоцитів крові. При пептичних виразках, ерозивних гастритах, невиразковій диспепсії, панкреатитах широко використовуються не тільки блокатори Н2-ГР (Дегтярьова I.І. і ін., 2000; Gespach C. et all., 1990; Kulkarni P. N. et all., 1997), але й блокатори М1-холінергічних рецепторів (Otsuki M. et all., 1986; Singer M. V. et all., 1991; Teyssen S. et all., 1995). Загальновизнаним у клінічній практиці М1-холінолітиком є пірензепін. Відомо, що він блокує ацетилхолінові рецептори СОШ і секреторних клітин підшлункової залози, зменшує їх функціональну активність та стимулює рівень кровоплину (Бендиков Э.А. и др., 1985; Omura N. et all., 1988; Otsuki M. et all., 1986; Singer M. V. et all., 1991; Teyssen S. et all., 1995). Відомо, що лімфоцити, як і нервові клітини, можуть активно та оперативно брати участь в індукції і регуляції стрес-реакцій організму шляхом синтезу та секреції різноманітних факторів (Давтян Т.К. и др., 2001). Лімфоцити, як і ацинарні клітини підшлункової залози, і клітини СОШ здатні експресувати різні субтипи М-холінергічних рецепторів, зокрема М1 (Bronzetti E. et all., 1996; Nomura J. et all., 2003). Можна припустити, що механізм дії інгібіторів М1 - холінорецепторів у цих клітинах подібний.
Встановлено, що вміст одного з кінцевих метаболітів ПОЛ - МДА у лімфоцитах із підвищенням концентрації пірензепіну зменшується. Тоді як із підвищенням концентрації пірензепіну вміст МДА у лімфоцитах зменшувався, активність ГП зростала із 156 ± 11 (контроль) до 189 ± 12 нмоль GSН (хв · мг білка) - 1. Пірензепін не тільки призводить до зростання активності ГП, як нами продемонстровано, але і до зростання активності супероксиддисмутази. Так, у результаті лікування хворих хронічним панкреатитом за допомогою пірензепіну, активність супероксиддисмутази у крові зростала у 2,5 рази (Дегтярьова I.І. і ін., 2000).
Оскільки функціонування ГП залежить від наявного пулу відновленого глутатіону, який забезпечується функціонуванням ГР, наступним етапом роботи було вивчення активності цього ферменту. Показано, що із підвищенням концентрації пірензепіну у пробах від 0 до 10-3 М ГР-а активність лімфоцитів збільшувалась.
За відсутності пірензепіну ГР-а активність складала 48±6 нмоль NADPH (хв · мг білка) - 1. По мірі підвищення концентрації препарату з 10-6 до 10-3 М активність зростала до 77,8±8 нмоль NADPH (хв · мг білка) - 1, виходячи на плато. Порівнюючи активності ГП і ГР при дії пірензепіну можна бачити, що ГР чутливіша до дії препарату, її активність змінюється у ширших межах.
Таким чином, отримані результати показують, що ефект пірензепіну на лімфоцити периферичної крові обумовлений не тільки його зв'язуванням із М1-холінергічними рецепторами (Nomura J. et all., 2003; Tayebati S. K. et all., 1999) і подальшою передачею сигналу через фосфатидилінозитидну систему, але і впливом на ПОЛ і глутатіонову АОС.
Отже, інгібітор М1-холінергічних рецепторів пірензепін інгібує ПОЛ та активує глутатіонову АОС лімфоцитів крові. Лімфоцити крові можна використовувати для вивчення механізмів дії препаратів, які реалізують свій ефект через мускаринові холінергічні рецептори.
При вивченні впливу пірензепіну на Са2+, Мg2+-АТФазну активність встановлено, що із підвищенням концентрації препарату активність ферменту знижується (табл.1). Са2+, Мg2+-АТФазна активність зменшувалась з 2,8±0,02 мкмоль Фн (хв · мг білка) - 1, у контрольних зразках, до 1,3±0,1 мкмоль Фн (хв · мг білка) - 1, за наявності в інкубаційному середовищі 10-3 М пірензепіну.
Na+, К+-АТФазна активність, яка у контрольних пробах становила 5,9±0,5 мкмоль Фн (хв · мг білка) - 1, пригнічувалась відповідно до підвищення концентрації пірензепіну, за 10-3 М концентрації препарата становила 0,4±0,04 мкмоль Фн (хв · мг білка) - 1. Отже, пірензепін також виявляє дозозалежний інгібувальний вплив щодо Na+, К+-АТФазної активності лімфоцитів.
Таблиця 1
Залежність АТФазних активностей лімфоцитів крові від концентрації пірензепіну (М±m)
| Концентрація пірензепіну, М | |||||
| 0 | 10-6 | 10-5 | 10-4 | 10-3 | |
| Кількість досліджень | n = 18 | n = 18 | n =18 | n = 18 | n = 18 |
| Са2+, Мg2+-АТФазна активність, мкмоль Фн (хв × мг білка) - 1 | 2,8±0,1 | 1,8 ±0,2 | 1,5± 0,1 | 1,4±0,1 | 1,3±0,1 |
| Na+, К+-АТФазна активність, мкмоль Фн (хв × мг білка) - 1 | 5,9±0,5 | 4,4±0,3 | 1,4±0,1 | 0,7±0,06 | 0,4 ±0,04 |
Можливо, така пригічувальна дія пірензепіну на АТФази пов'язана із антиоксидантними властивостями препарату, які вже встановлені на сьогоднішній час (Бендиков Э.А. и др., 1985; Дегтярьова I.І. і ін., 2000; Bronzetti E. et all., 1996). У дослідженнях на лімфоцитах периферичної крові людини нами засвідчено функціональний зв’язок між глутатіоновою антиоксидантною та Са2+-транспортувальною системами.
Активність ферментів глутатіонової антиоксидантної системи і транспортувальних АТФаз лімфоцитів крові при дії блокатора протонної помпи омепразолу. Відомо, що Н+, К+-АТФаза (протонна помпа) СОШ відповідає за секрецію соляної кислоти. При цьому активний транспорт іонів водню та калію здійснюється у протилежних напрямках для генерації надмірного градієнта Н+ через мембрану за фізіологічних умов (Asano S. et all., 2001; 2004).
При лікуванні широкого спектру кислотоасоційованих захворювань верхніх відділів травного тракту широко використовується блокатор протонної помпи омепразол (Заїка С.В., 2000; Burdan F. et all., 2000; Nakamura T. et all., 1995). Однак, ймовірно, механізм дії цього препарату більш широкий. Представляє інтерес вивчення його впливу на активність ферментів глутатіонової АОС, оскільки розвиток багатьох захворювань супроводжується оксидативним стресом і, відповідно, інтенсивною продукцією активних форм кисню (Барабой В.А. и др., 1997; Владимиров Ю.А., 1989).
Хоча Н+, К+-АТФаза експресується переважно у СОШ, вона також виявлена і у лімфоцитах (Bergman M. P. et all., 2003). На пермеабілізованих лімфоцитах периферичної крові нами встановлено, що із підвищенням концентрації омепразолу до 10-4 М активність ГП поступово зростає, досягаючи рівня 1,46 ± 0,1 мкмоль GSH (хв · мг білка) - 1, що в 4,8 рази вище контрольних значень (0,3 ± 0,2 мкмоль GSH (хв · мг білка) - 1). Концентрації препарату понад 10-4 М призводили до зниження активності ГП (рис.5).
Зміни активності ГР полягали у її зростанні з 0,04 ± 0,05 мкмоль NADPH (хв · мг білка) - 1, за відсутності омепразолу, до 0,22 ± 0,03 мкмоль NADPH (хв · мг білка) - 1, при його концентрації 10-4 М. При подальшому підвищенні концентрації цього препарату спостерігалось деяке зменшення активності ферменту.
Встановлено, що ефект омепразолу на активність обох АТФаз є дозозалежним. Оптимальні концентрації препарату (10-3 М) пригнічують активність Са2+, Мg2+-АТФази у 2,4 рази, з 2,8±0,1 до 1,2 ± 0,1 мкмоль Фн (хв · мг білка) - 1 (Рис.6), а активність Nа+, К+-АТФази - у 9,8 рази, з 5,9±0,5 до 0,6±0,06 мкмоль Фн (хв · мг білка) - 1.
Таким чином, у результаті виконання цієї роботи встановлено, що блокатори Н2-гістамінових рецепторів, М1-холінергічних рецепторів, протонної помпи – фамотидин, пірензепін, омепразол, відповідно, окрім основної дії, пов’язаної з їх взаємодією з відповідними рецепторами чи протонною помпою та залученням у цей процес G-білків і фосфатидилінозитидної та аденілатциклазної систем, мають і інший механізм дії. Цей механізм має не менш важливе значення, оскільки у ньому задіяна пероксидація ліпідів, ферменти глутатіонової антиоксидантної системи, іон-транспортувальні системи. Оскільки, як вказувалось вище, у лімфоцитах периферичної крові експресуються Н2-гістамінові, М1-холінергічні рецептори, Н+, К+-АТФаза, вони можуть бути адекватною та зручною моделлю для вивчення механізму дії цих чи інших блокаторів вказаних рецепторів та протонної помпи.
ВИСНОВКИ
У дисертаційній роботі, відповідно до поставленої мети та завдань дослідження, на пермеабілізованих моноядерних лімфоцитах периферичної крові людини вперше отримані дані про роль глутатіонової антиоксидантної системи і транспортувальних АТФаз у реалізації ефектів блокаторів Н2-гістамінових, М1-холінергічних рецепторів і блокатора протонної помпи та про взаємозв’язок між антиоксидантною системою і Са2+. З отриманих результатів можна зробити наступні висновки:
1. Пермеабілізація лімфоцитів сапоніном (0,1... .0,2%) розкриває латентну активність глутатіонпероксидази, глутатіонредуктази, глутатіонтрансферази, Са2+, Мg2+ - та Nа+, К+-АТФаз, що дає можливість вивчити функціональні зв’язки між цими ферментами та з’ясувати їх роль у реалізації ефектів фармакопрепаратів.
2. Катіони кальцію регулюють процес пероксидації ліпідів і систему антиоксидантного захисту – активності глутатіонпероксидази, глутатіонредуктази, глутатіонтрансферази, здійснюючи стимулюючий вплив на ферменти при низьких концентраціях та інгібуючий вплив у концентраціях, що перевищують 0,1 мМ.
3. Показано, що один із шляхів реалізації фізіологічного ефекту різних за механізмом дії фармакологічних препаратів (фамотидин, пірензепін, омепразол) опосередковується глутатіоновою антиоксидантною системою.
4. Гістамінові Н2-рецептори приймають участь у регулюванні процесів вільнорадикального окиснення. Їх активування призводить до зростання вмісту продуктів пероксидації ліпідів у лімфоцитах крові.
5. Встановлено, що інгібітор Н2-гістамінових рецепторів фамотидин дозозалежно впливає на активність глутатіонпероксидази та глутатіонредуктази – при низьких концентраціях активує ці ферменти, а при високих (>10-3 М) інгібує. Найчутливішою до дії оптимальних концентрацій (10-4 М) фамотидину, пірензепіну та омепразолу є глутатіонредуктаза, активність якої зростає у 8,7, 1,6 та 5,5 разів, відповідно.
6. Встановлено, що фамотидин, пірензепін і омепразол реалізують свої ефекти також через Са2+, Мg2+ - і Nа+, К+-АТФази. Фамотидин у концентрації 10-4 М повністю пригнічує функціонування вказаних АТФаз. Пірензепін у тій же концентрації пригнічує їх у 2,0 і 8,4 рази, а омепразол - у 2,0 та 4,9 рази, відповідно.
7. Моноядерні лімфоцити периферичної крові є адекватною моделлю для вивчення механізмів функціонування ферментів глутатіонової антиоксидантної системи і транспортувальних АТФаз. Вони можуть використовуватись як експериментальна модель для виявлення індивідуальної чутливості клітин до деяких фармакологічних препаратів, а також для дослідження їх мембранозалежних механізмів дії.
СПИСОК ОПУБЛІКОВАНИХ ПРАЦЬ
1. Підковка Н.О., Зіменковський А.Б., Кімакович О. В.,* Воробець З.Д. Фармакологічна активність квамателу та функціонування іон-транспортуючих систем у лімфоцитах крові // Практична медицина. - 2002. - Т.8, № 3. - С.109-112. (Здобувач проводила літературний пошук, аналізувала літературні дані, визначила мету і задачі роботи, налагодила методики та здійснила фрагмент досліджень, опрацювала статистично результати, визначила наукову новизну, підготувала матеріал до друку).
2. Підковка Н.О., Кімакович О.В., Воробець З.Д. Функціонування глутатіонової антиоксидантної системи та Са2+, Мg2+-АТФази лімфоцитів крові за умов окиснювального стресу // Acta Medica Leopoliensia / Львівський медичний часопис. - 2003. - Т. VIV, № 1. - С.16-19. (Здобувач визначила мету роботи, провела літературний пошук, планування схеми наукового дослідження, виконала фрагмент експериментальної частини, проаналізувала отримані результати, сформулювала висновки, підготувала матеріал до друку).
3. Воробець З.Д., Кімакович О.В. Активність ферментів глутатіонової антиоксидантної системи лімфоцитів периферичної крові при дії пірензепіну // Медична хімія. - 2004. - Т.6, № 3. - С.95-97. (Здобувач провела літературний пошук, брала участь у проведенні експерименту, інтерпретувала отримані результати, підготувала матеріал до друку).
4. Кімакович О.В., Підковка Н.О., Воробець З.Д. Дія квамателу та пірензепіну на активність транспортних АТФаз лімфоцитів периферійної крові // Практична медицина. - 2004. - Т.10, № 2. – С.86-89. (Здобувач визначила мету роботи, постановку задач дослідження, відпрацювала схему наукового дослідження, виконала фрагмент експериментальної частини, проаналізувала отримані результати, підготувала матеріал до друку).
5. Кімакович О.В., Воробець З.Д. Вплив омепразолу на активність ферментів глутатіонової антиоксидантної системи лімфоцитів периферичної крові // Буковинський медичний вісник. - 2005. – Т.9, № 2. – С.112-114. (Здобувач провела аналіз даних літератури, визначила мету роботи, приймала участь у постановці експерименту, проаналізувала отримані результати, підготувала матеріал до друку).
6. Vorobets Z., Kimakovich О. Effect of pirenzepine on glutathione antiperoxide system function in human blood lymphocytes // Annales Universitatis Mariae Curie-Sklodowska. – 2004. – Vol. XVII, № 2. – С.319-320. (Здобувач визначила мету роботи, постановку задач, провела літературний пошук, виконала фрагмент експериментальної частини, проаналізувала отримані результати, сформулювала висновки, підготувала матеріал до друку).
7. Vorobets Z., Kimakovich O., Kimakovich V. Еffect of proton pump blocker on enzyme activity of glutathione antioxidant system of the peripheral blood lymphocytes // Annales Universitatis Mariae Curiae-Sklodowska. 2006. – Vol. XIX, № 1. – P.131-134. (Здобувач обґрунтувала мету роботи, визначила задачі, провела літературний пошук, виконала фрагмент експериментальної частини, проаналізувала отримані результати, сформулювала висновки, підготувала матеріал до друку).
8. Kimakovich O., Podkovka N., Vorobets Z. Modulation of glutathione and ion-transporting systems activity of blood lymphocytes by Quamatel // International Conference Neuro-humoral and cellular regulatory mechanisms of digestion processes. Abstracts. – Lviv, 2003. – P.15-16. (Здобувач провела літературний пошук, визначила мету роботи, постановку задач, виконала фрагмент експериментальної частини, проаналізувала отримані результати, сформулювала висновки, підготувала матеріал до друку).
9. Воробець З.Д., Кімакович О.В. Властивості Са2+, Мg2+-АТФази пермеабілізованих лімфоцитів крові // Установчий з”їзд Українського товариства клітинної біології. Тези доп. – Львів, 2004. – С.238. (Здобувачем здійснено відпрацювання ідеї, планування схеми наукового дослідження, аналіз літературних даних, участь у плануванні та проведенні експерименту, аналіз отриманих результатів, також сформулювала висновки, підготувала матеріал до друку).
10. Кімакович О.В., Воробець З.Д. Блокатор М1-холінергічних рецепторів гастроцепін у функціонуванні транспортних АТФаз // Матеріали міжнародної конференції "Клітинні та субклітинні механізми функціонування травної системи". Тези доп. – Львів, 2004. – С.30. (Здобувач проводила літературний пошук, здійснювала підготовку до експерименту, брала участь у його проведенні, аналізі та узагальненні отриманих результатів, підготувала матеріал до друку).
11. Воробець З.Д., Кімакович О.В. Взаємозв’язок між активністю транспортувальних АТФаз і системою глутатіонового антиоксидантного захисту у лімфоцитах периферичної крові // Матер. ІХ Українського біохімічного з’їзду. Тези доп. – Харків, 2006. - С.109-110. (Здобувач відпрацювала ідею, планування схеми наукового дослідження, аналіз літературних даних, участь у плануванні та проведенні експерименту, аналіз та узагальнення отриманих результатів, сформулювала висновки, підготувала матеріал до друку).
12. Воробець З.Д., Кімакович О.В. Са2+, Мg2+ - та Nа+, К+-АТФазні активності лімфоцитів периферичної крові за дії блокатора протонної помпи омепразолу // Міжнародна наук. конф. „Механізми функціонування фізіологічних систем”. Тези доп. – Львів, 2006. – С.30. (Здобувач провела літературний пошук, визначила задачі дослідження, виконала фрагмент експериментальної частини, проаналізувала отримані результати, сформулювала висновки, підготувала матеріал до друку).
13. Kimakovich O. V., Vorobets Z. D. Humen lymphocytes glutathione antioxidant system function under omeprazole action // III International Conference Neuro-humoral and cellular regulatory mechanisms of digestion processes. Abstracts. – Lviv, 2007. – P.31-32. (Здобувач відпрацювала концепцію наукового дослідження, налагодила та обґрунтувала схему експерименту, опрацювала, здійснила інтерпретацію та узагальнення отриманих даних, визначила наукову новизну, підготувала статтю до друку).
АНОТАЦІЯ
Коноварт О.В. Механізми дії блокаторів Н2-гістамінових і М1-холінергічних рецепторів у лімфоцитах периферичної крові.
Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю 14.03.03 – нормальна фізіологія. – Львівський національний медичний університет імені Данила Галицького, Львів, 2008.
Вивчені у комплексі окремі ланки глутатіонової антиоксидантної системи і транспортувальних АТФаз та їх чутливість до іонів кальцію у моноядерних лімфоцитах периферичної крові. Встановлені оптимальні концентрації сапоніну, які розкривають латентні активності глутатіонпероксидази, глутатіонредуктази, глутатіонтрансферази, Са2+, Мg2+ - та Nа+, К+-АТФаз у лімфоцитах периферичної крові, що дає змогу простежити функціональні зв’язки між цими ферментами. Підібрані оптимальні умови для вивчення активностей цих ферментів у пермеабілізованих лімфоцитах.
Виявлено, що блокатори Н2-гістамінових рецепторів (фамотидин), М1-холінергічних рецепторів (пірензепін) та протонної помпи (омепразол) дозозалежно впливають на активність ферментів глутатіонової антиоксидантної системи і транспортувальних АТФаз, а саме, при низьких концентраціях активують глутатіонпероксидазу та глутатіонредуктазу, при високих – інгібують, одночасно дані препарати інгібують активність Са2+, Мg2+ - та Nа+, К+-АТФаз.
Ключові слова: лімфоцити, пероксидація ліпідів, глутатіонпероксидаза, глутатіонредуктаза, АТФази, Н2-гістамінові рецептори, М1-холінергічні рецептори, фамотидин, пірензепін, омепразол.
АННОТАЦИЯ
Коноварт О.В. Механизмы действия блокаторов Н2-гистаминовых и М1-холинергических рецепторов у лимфоцитах периферической крови.
Диссертация на соискание ученой степени кандидата медицинских наук по специальности 14.03.03 – нормальная физиология. – Львовский национальный медицинский университет имени Данила Галицкого, Львов, 2008.
Изучены в комплексе отдельные звенья глутатионовой антиоксидантной системы и транспортирующих АТФаз и их чувствительность к ионам кальция в моноядерных лимфоцитах периферической крови. Определены оптимальные концентрации сапонина, которые раскрывают латентные активности глутатионпероксидазы, глутатионредуктазы, глутатионтрансферазы, Са2+, Мg2+ - и Nа+, К+-АТФаз в лимфоцитах периферической крови, что дает возможность проследить функциональные связи между этими ферментами. Подобраны оптимальные условия для изучения активности этих ферментов у пермеабилизированных лимфоцитах.
Извесно, что острые и хронические панкреатиты, повышеная желудочная секреция сопровождаются оксидативным стрессом. Для их лечения используют блокаторы Н2-гистаминовых и М1-холинергических рецепторов. Также для знижения гиперпродукции хлоридной кислоты париетальными клетками слизистой оболочки желудка используют блокаторы протонного насоса. Извесно, что блокатор М1-холинергических рецепторов пирензепин, в отличии от неселективних веществ, избирательно угнетает секрецию кислоты и пепсина в слизистой оболочке желудка, уменшает функциональную активность ацинарных клеток поджелудочной железы, улутшает микроциркуляцию и кровознабжение. Селективный блокатор Н2-гистаминовых рецепторов – фамотидин – не только тормозит секрецию желудочных желез, но и ингибирует генерацию активных форм кислорода, активность аденилатциклазы, синтез панкреатичных ферментов.
Показано, что ингибиторы Н2-гистаминовых рецепторов фамотидин дозозависимо влияет на активность глутатионпероксидазы и глутатионредуктазы – при низких концентрациях активирует эти ферменты, а при высоких (>10-3 М) ингибирует. Одновременно этот препарат в концентрациях >10-4 М практически полностью ингибирует Са2+, Мg2+ - и Nа+, К+-АТФазные активности лимфоцитов периферической крови.
Блокатор М1-ацетилхолинергических рецепторов пирензепин ингибирует пероксидацию липидов и стимулирует активность глутатионпероксидазы. Этот же блокатор дозозависимо ингибирует активность Са2+, Мg2+ - и Nа+, К+-АТФаз.
Блокатор протонного насоса омепразол в концентрациях до 10-4 М стимулирует активность глутатионпероксидазы и глутатионредуктазы, а в больших концентрациях ингибирует их. Омепразол также, подобно к фамотидину и пирензепину, дозозависимо ингибирует активность Са2+, Мg2+ - и Nа+, К+-АТФаз.
Изменение активности ион-транспортирующих систем и ферментов антиоксидантной системы во время действия разных блокаторов может соправаждатся модуляцией синтеза лимфоцитами биологически активных веществ и возможностю клеток к хемотаксису. Так как Н2-гистаминовые и М1-ацетилхолинергические рецепторы широко экспресируются не только в клетках слизистой оболочки желудка или ацинарных клетках поджелудочной железы, но и в лимфоцытах периферической крови, эти клетки могут быть удобной моделью для изучения механизмов действия блокаторов данных рецепторов, которые используются для лечения кислотозависимых заболеваний и панкреатитов. Полученные данные могут служить базой для разроботки методов коррекции патологических изменений клеточного гомеостаза. В связи с этим можно ставить вопрос о создании и подборе новых фармакологических препаратов, которые модулируют активность глутатионовой антиоксидантной системы и транспортирующих АТФаз.
Ключевые слова: лимфоциты, пероксидация липидов, глутатионпероксидаза, глутатионредуктаза, АТФазы, Н2-гистаминовые рецепторы, М1-холинергические рецепторы, фамотидин, пирензепин, омепразол.
SUMMARY
Konowart O. V. Mechanism of effect of blockers of H2-hystamine and M1-cholinergic receptors in peripheric blood limphocytes. – Manuscript.
The Dissertation for obtaining the scientific degree of Candidate of medical sciences, speciality 14.03.03 – Normal physiology. Danylo Galickyj Lviv National Medical University, Lviv, 2008.
Separate branches of glutathione antioxidant system and ATPase transporters, their sensitivity to calcium ions in mononuclear lymphocytes in peripheral blood have been studied in complex. Optimal saponin concentrations have been established, which reveal latent activities of glutathione peroxidase, glutathione reductase, glutathione transferase, Ca²+,Mg²+ - and Na+,K+-ATPase in lymphocytes in peripheral blood, that enables to follow functional connections between these enzymes. Optimal conditions have been found to investigate the activity of these enzymes in permeable lymphocytes.
It has been detected, that inhibitor of H2-histamine receptors (phamotydine), M1–acetylcholine receptors (pirenzepine), blocker of proton pump (omeprazole) in certain doses affects the activity of glutathione antioxidant system and ATPase transporters - in low concentrations it activates of glutathione peroxidase and glutathione reductase, in high concentrations inhibits them. This blockers in certain doses inhibits the activity of Ca²+,Mg²+ - and Na+,K+-ATPase.
Key words: lymphocytes, peroxide lipid oxidation, glutathione peroxidase, glutathione reductase, ATPase, H2–histamine receptors, M1-choline receptors, phamotydine, pirenzepine, omeprazole.
0 комментариев