Реферат:
НАРУШЕНИЯ БЕЛКОВОГО ОБМЕНА
Белковый обмен занимает особое место в многообразных превращениях веществ, характерных для всех живых организмов. Биологическое значение белков определяется их многообразными функциями. Белки определяют микро– и макроструктуру отдельных субклеточных образований, клеток, органов и целостного организма, т.е. выполняют пластическую функцию. Белковый обмен обеспечивает непрерывность воспроизводства и обновления белковых тел организма. Энгельс охарактеризовал белки как материальный носитель жизни и подчеркнул динамичность белкового обмена. Он писал: "Жизнь есть способ существования белковых тел, и этот способ существования состоит по своему существу в постоянном самообновлении химических составных частей этих тел".
Помимо пластической роли, белки выполняют уникальную, функциональную, т.е. каталитическую роль. Этой функцией не наделены ни углеводы, ни жиры.
Белки (соответственно и продукты их гидролиза – аминокислоты) принимают непосредственное участие в биосинтезе рада гормонов, биологически активных веществ и медиаторов. К ним относятся либерины и статины гипоталамуса, инсулин, ангиотензин, кинины, гистамин, серотонин и др. В последние годы стали известны пептиды, снижающие болевую чувствительность – эндорфины.
Белки (особенно альбумины) поддерживают онкотическое давление крови. Являясь гидрофильными коллоидами, они связывают определенное количество воды и удерживают ее в кровеносном русле.
Белки участвуют в сложной системе регуляции гомеостаза. Они поддерживают рН крови, представляя собой так называемый белковый буфер.
Главную роль в процессах мышечного сокращения и расслабления выполняют актин и миозин – специфические белки мышечной ткани. Сократительная функция присуща не только мышечным белкам, но и белкам ряда субклеточных структур, что обеспечивает тончайшие процессы жизнедеятельности клеток.
Основную функцию защиты в организме выполняет иммунная система» которая обеспечивает синтез специфических защитных белков–иммуноглобулинов. В качестве другого примера защитной роли можно привести участие ряда белков крови в процессе свертывания,
Белки выполняют транспортную функцию: они соединяются с различными веществами (гормонами, витаминами, жирами, медью, железом и др.), обеспечивая их доставку в ткани–мишени.
При определенных условиях, например, голодании, сахарном диабете белки могут использоваться как энергетический материал.
Таким образом, белковый обмен координирует, регулирует и интегрирует процессы обмена веществ в организме, подчиняя его сохранения вида, непрерывности жизни. Состояние белкового обмена определяется множеством экзо– и эндогенных факторов. Любые отклонения от нормального физиологического состояния организма отражаются на белковом обмене. Поэтому знание закономерностей этих изменений при конкретном патологическом процессе имеет важное значение для правильного понимания механизмов болезни и выбора тактики терапевтических мероприятий.
Нарушения биосинтеза и распада белков в органах и тканяхБелковый обмен обеспечивает непрерывность воспроизводства и обновления белков организма. Показано, что в среднем каждые 3 недели половина белковых компонентов человеческого тела полностью обновляется путем распада и ресинтеза. При этом общая скорость синтеза белков в организме в состоянии азотистого равновесия достигает 500 г в день, т.е. почти в. 5 раз превосходит среднее потребление с пищей. Естественно, что такой результат, может быть обеспечен только за счет повторного использования аминокислотных предшественников и продуктов распада белков. Белки органов и тканей нуждаются в постоянном обновлении. В конечном счете, животным необходим не белок как таковой, а определенные аминокислоты, освобождающиеся при его гидролизе. Известно, что у детей продолжительный недостаток гистидина приводит к нарушению образования гемоглобина к возникновению экземы. Дефицит основной аминокислоты аргинина не сказывается на нормальном росте, но может привести к нарушению сперматогенеза.
На величины потребностей в определенных аминокислотах существенно влияет состав общей смеси аминокислот, получаемой организмом; так, например, потребность в фенилаланине и метионине значительно уменьшается при достаточном обеспечении тирозином и цистеином, так как фенилаланин превращается в организме в заменимую аминокислоту тирозин, а метионин метаболизируется с образованием заменимой аминокислоты цистеина. Если в эксперименте молодые крысы получают лишь минимальные количества незаменимых аминокислот, то поразительным стимулом для роста животных оказывается введение в рацион некоторых заменимых аминокислот, например, глутаминовой кислоты и аргинина. В то же время введение в рацион больших количеств других аминокислот, особенно глицина, может привести к значительному замедлению роста. Для оптимального синтеза белков, следовательно, необходима сбалансированная смесь аминокислот.
В основе развития ряда патологических состояний в организме лежат нарушения динамического равновесия двух фаз метаболизма: анаболизма и катаболизма белковых структур. Патологические изменения в анаболической фазе могут возникать вследствие дефектов генетического кода и нарушений отдельных этапов биосинтеза белков: репликации, транскрипции, трансляции и посттрансляционных модификаций молекул. Чаше всего следствием таких нарушений является дефицит одного иди нескольких белков, что в зависимости от их функциональной значимости приводит к развитию различных клинических проявлений. Так, например, при сахарном диабете снижается белок–синтезирующая активность рибосом, что, по–видимому, связано с нарушениями процесса инициации. Установлено, что некоторые виды наследственных анемий человека – талассемии – характеризуются нарушением процесса трансляции β–мРНК или нехваткой белковых факторов трансляции. Есть данные, свидетельствующие о том, что прикрепление мРНК к рибосомам, а также процессу транслокации могут тормозиться дифтерийным токсином. К ингибиторам инициации можно отнести и ряд широко используемых антибактериальных антибиотиков, тормозящих присоединение аминоацил–тРНК на всех стадиях трансляции. К ним относятся антибиотики тетрациклинового ряда, аминогликозидные антибиотики (стрептомицин, неомицин, канамицин и др.). Образование аминоацил–тРНК и белка может быть подавлено препаратами салициловой кислоты. Показано, что афлатоксины – продукты жизнедеятельности гриба Aspergillus flavus, подавляют синтез ДНК и митоз клеток. Установлено, что некоторые антибиотики (рифамицины, рифампицин) нарушают матричный синтез РНК, подавляя активность ДНК–зависимой РНК–полимеразы.
В подавляющем большинстве случаев не представляется возможным говорить об изолированных нарушениях анаболической фазы белкового обмена, так как они обязательно сочетаются с нарушениями катаболизма. Это наблюдается при общем и белковом голодании, дефиците отдельных незаменимых аминокислот, изменениях в последовательности поступления аминокислот, так как белковый синтез подчинен закону "все или ничего". Выраженный дисбаланс процессов синтеза и распада белков имеет место при нарушении гормональной регуляции и влияний центральной нервной системы.
Наследственные дефекты биосинтеза белков
Генетически обусловленные нарушения структуры, а следовательно, и свойств белков представляют собой в сущности группу моногенных наследственных болезней. Она возникают в результате точечных мутаций как структурных, так и регуляторных генов и передаются в поколениях в соответствии с законами Менделя.
Фенотипические проявления этих заболеваний обусловлены функциональными свойствами белков, их органной и тканевой принадлежностью, значимостью для метаболизма и т.д. Следует отметать, что первичный генетический дефект ряда моногенных болезней (муковисцедоз, ахондроплазия, мышечные дистрофии и др.) пока на выяснен, но роль нарушений биосинтеза белков несомненна.
Энзимопатии являются наиболее изученной и представительной группой этих заболеваний, характеризующихся наследственной недостаточностью каталитической активности отдельных ферментов. Этот дефект наследуется, как правило, по аутосомно–рецессивному типу. Фенотипические проявления во многом обусловлены нарушениями биохимических закономерностей течения реакция. К ним относятся избыток субстрата, нерасщепляемого мутантным ферментом; недостаток продуктов реакции, катализируемой этим ферментом и, наконец, появление соединений, являвшихся продуктами функционирования смежных или побочных метаболических путей (например, при алкаптонурии, альбинизме, фенилкетонурии).
Иногда симптомы заболевания появляются под влиянием провоцирующих факторов. Примером могут служить гемолитические кризы у больных с дефицитом глюкозо–6–фосфатдегидрогеназы в эритроцитах при назначении противомалярийных и некоторых других лекарственных препаратов. В ряде случаев энзимопатии представляют собой тяжелые заболевания, симптомы которых появляются в раннем детском возрасте, как это имеет место при фенилпировиноградной олигофрении.
Выделение других групп генетически обусловленных дефектов белков, помимо энзимопатий, довольно условно, так как один и тот же белок может выполнять в организме несколько функций, в том числе и каталитическую, через которую и опосредуется физиологический эффект. Примером может служить выделение в отдельную группу наследственных дефектов синтеза факторов свертывания крови.
Общность функций белков, ответственных за транспорт веществ в организме, объединяет в одну группу заболевания с самыми различными клиническими проявлениями. Наиболее распространенными среди них являются гемоглобинопатии – наследственные дефекты синтеза гемоглобина, при которых возможно развитие серьезных нарушений транспорта газов в организме. К этой же категории относятся различные типы наследственных дислипопротеидемий, проявления которых связаны с нарушением транспорта холестерина и других липидов.
Генетические дефекты могут явиться также причиной нарушения синтеза структурных белков организма. Чаще всего это связано с патологическими изменениями структуры пластических белков соединительной ткани. Биохимические дефекты фибриллярного белка – коллагена объединяются в группу коллагеновых болезней, характеризующихся полиморфностью клинических проявлений. Врожденный адреногенитальный синдром связан с генетически обусловленной неполноценностью ферментных систем стероидогенеза в коре надпочечников. Недостаток ферментов (в первую очередь 21–гидроксилазы), участвующих в биосинтезе кортикостероидов, приводит к снижению продукции кортизола и накоплению в крови предшественников его метаболизма (17–окси–прогестерон, прогестерон). Растормаживание секреции кортикотропина стимулирует выработку андрогенов, которые оказывают вирилизирующее действие на детский организм. При глубоком дефиците 21–гидроксилазы также происходит резкое снижение биосинтеза альдостерона, в результате развивается сольтеряющий синдром.
В ряде случаев нарушения функционирования иммунной системы связаны с патологическими изменениями в геноме. Наследственная недостаточность иммунного ответа может затрагивать специфические механизмы гуморального и клеточного иммунитета. В этих случаях речь идет о первичных иммунодефицитах. Наследственные дефекты неспецифических факторов защиты (фагоцитоза, системы комплемента и др.) могут встречаться как в изолированных вариантах, так и в комбинации с дефектами специфических факторов иммунной защиты.
Вторичные нарушения биосинтеза и распада белков в организмеИнтенсивность фаз анаболизма и катаболизма белков в клетках зависит от их функционального состояния, изменения регуляторных влияний, характера развивающихся патологических процессов. Знание активности и соотношения этих фаз представляет определённый практический интерес. В ряде случаев реальная возможность для оценки метаболизма белков в органах и тканях появляется при исследовании белков крови. Это связано с тем, что белки плазмы крови синтезируются в клетках различных органов и систем: в печени, иммуноцитах, клетках системы мононуклеарных фагоцитов и т.д. Патологические и компенсаторные процессы в этих структурах отражаются в конечном итоге на показателях белкового состава плазмы крови. Другой механизм изменения белкового спектра крови заключается в том, что при нарушении целостности мембран клеток в кровь могут поступать белки, несвойственные нормальному составу крови. В данном случае речь идет о ферментах, определение изменений активности которых в крови имеет большое диагностическое и прогностические значение. Ряд патологических процессов в органах сопровождается повышением активности некоторых ферментов в крови – гиперферментемией. При развитии некрозов в отдельных органах (инфаркт миокарда, острые гепатиты и панкреатиты, отравления CCl4 и др.) вследствие разрушения клеток тканевые трансаминазы (аспартат– и аланинаминотгрнсферазы) поступают в кровь и повышение их активности в таких случаях является одним из диагностических тестов.
Активность кислой фосфатазы в сыворотке крови увеличивается при раке простаты, активность альдолазы возрастает при мышечной дистрофии. Увеличение активности амилазы в плазме крови наблюдается при острых панкреатитах, язве двенадцатиперстной кишки, в то время как при гепатитах, раке печени, остром алкоголизме активность этого фермента уменьшается. При заболеваниях печени и отравлении фосфорорганическими соединениями падает активность холинэстеразы. При инфаркте миокарда в плазме увеличивается содержание четвертой и пятой изоформ лактатдегидрогеназы.
Количественные изменения в белковом составе крови могут проявляться в виде: гиперпротеинемии и гипопротеинемии. Однако эти показатели далеко не всегда отражают и моющиеся изменения в белковом составе. В случае разнонаправленных изменений белковых фракций, а также при нарушениях синтеза отдельных белков, концентрация которых в плазме невелика, суммарное содержание белков остается неизменным. В связи с этим получил широкое распространение термин диспротеинемия. Его используют не только для обозначения изменений суммарного количества белков в крови, но и в случаях изменения соотношения в содержании отдельных белковых фракций. Появление в крови белков с измененной структурой, несвойственных здоровому организму, обозначается как парапротеинемия.
Чрезвычайно тесные взаимоотношения существуют между процессами биосинтеза белков в печени и белковым составом плазмы крови. В гепатоцитах синтезируются все альбумины крови, до 90% альфа–глобулинов. В клетках Купфера образуется до 50% бета–глобулинов плазмы крови. Поэтому патологические процессы в печени (воспаление, дистрофия, опухоли, цирроз и др.) сопровождаются нарушением биосинтеза белков плазмы крови.
Уменьшение альбуминов проявляется не только в снижении содержания этой фракции в крови (гипоальбуминемии), но и сказывается на общем количестве белков – развивается гипопротеинемия, главным следствием которой является снижение онкотического давления крови и развитие отеков. Следует откатить, что гипопротеинемии могут быть связаны и с активацией процессов катаболизма при острых инфекциях, некоторых эндокринных заболеваниях и т.д. Часто встречаются гипопротеинемии, связанные с потерей белков, например, при кровопотерях, нефротическом синдроме, ожогах, обширных раневых поверхностях, в послеоперационном периоде, при злокачественных опухолях, лейкозах и т.д. (см. рис. 1).
|
| Компенсаторный синтез глобулинов при нарушении синтеза альбуминов (хронические диффузные поражения печени) | Синтез защитных белков – антител, С–реактивного белка и др. (инфекции, иммунитет, воспаление) | Переход клеточных белков в кровь | |||||||||
|
| ||||||||||||
| Гиперпротеинемия, увеличение относительного или абсолютного содержания всех или некоторых белков | ||||||||||||
|
| ||||||||||||
| Эупротеинемия | ||||||||||||
|
| ||||||||||||
| Гипопротеинемия, уменьшение относительного или абсолютного содержания всех или некоторых белков | ||||||||||||
|
| ||||||||||||
|
| Ускоренный распад отдельных белков | Потеря белков при нарушении целостности или проницаемости сосудистой стенки | ||||||||||
| в наружную среду (протеинурии при заболеваниях почек) | во внешнюю среду (отёки, шок, некоторые виды укусов и др.) | |||||||||||
| смешанные формы (ожоги, кровопотери) | ||||||||||||
| Уменьшение синтеза вследствие недостаточности или качественной диспропорции аминокислот (полное или частичное голодание, лихорадка, нарушение пищеварения и всасывания) | ||||||||||||
| ||||||||||||
Рис. 1. Классификация диспротеинемий по И.А. Ойвину
Нарушения биосинтеза некоторых белков плазмы крови часто не отражаются на общем их содержании, однако могут явиться причиной развития ряда патологических состояний. Примерами могут служить:
– геморрагические синдромы при нарушениях образования протромбина, фибриногена и других факторов свертывания крови;
– нарушения механизмов антимикробной резистентности при недостаточном синтезе компонентов системы комплемента;
– анемии, обусловленные недостаточностью трансферрина и ферритина – белков, необходимых для реутилизации железа;
– гепатоцеребральная дистрофия (болезнь Вильсона) при нарушениях образования церулоплазмина, участвующего в транспорте меди.
Чрезвычайно многообразны проявления нарушений процессов биосинтеза и распада белков в организме при синдроме белковой недостаточности.
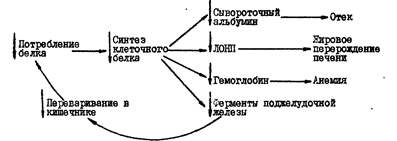
Рис. 2. Порочный круг при квашиоркоре
В экономически бедных районах тропической Америки, Центральной и Южной Африки, Индии недостаточное количество белка в рационе является важным этиологическим фактором заболевания детей – квашиоркора – на языке Ганы "красный мальчик" (син.: гидрокахексия, пеллагра детская, синдром "депигментация – отёк").
Заболевание характеризуется задержкой роста, анемией, гипопротеинемией (часто с отеком), жировой инфильтрацией печени, атрофией ацинарных клеток поджелудочной железы, сопровождавшейся диарреей и стеаторреей (см. рис. 2).
Поступление в организм недостаточного количества белка приводит к нарушению функций кишечника и неэффективному использованию тех небольших количеств белка, которые поступают о пищей (ЛОНП, липопротеиду очень низкой плотности).
Патологические изменения белкового обмена на этапах биосинтезе и распада белков могут происходить из-за нарушения процессов регуляции, которые осуществляются нервной я эндокринной системами, Нервные влияния могут реализоваться либо прямым воздействием на метаболизм белков, либо опосредованно через железы внутренней секреции. Денервация органов и тканей приводит к нарушению их питания развитию атрофии. При этом происходит активация протеолиза и торможение биосинтеза белков. Примерами таких состояний могут служить прогрессирующие мышечные дистрофии, развивающиеся вследствие поражений вегетативной нервной система на различных уровнях.
Механизмы влияний гормонов на белковый обмен различны. Эти влияния могут осуществляться путем воздействия на геном клетки, что определяет в конечном итоге количество вновь образующегося белка. Под гормональным контролем находится активность ферментов. Это предопределяет возможность регуляторных влияний как на скорость течения отдельных биохимических реакций, так и на различные фазы обмена. Примерами патологических состояний, обусловленных нарушением эндокринных влияний на фазы белкового обмена, могут быть акромегалия и гигантизм, гипофизарная кахексия (болезнь Симмондса), гипофизарный нанизм; истощение с выраженным отрицательным азотистым балансом, наблюдающиеся при тиреотоксикозе и др.
Усиление распада белков в тканях может наблюдаться при различиях типических патологических процессах (воспаление, аллергия, ишемия и т.д.) и при ряде инфекционных заболеваний, сопровождающихся лихорадкой, при интоксикациях, обширных ожогах и травмах мягких тканей. Повышенный катаболизм белков в этих случаях может носить как локальный, так и генерализованный характер.
Среди других факторов, ограничивающих синтез белка, следует отметить гипоксию. Дефицит кислорода в тканях вызывает нарушение всех видов обмена, в том числе и пластического. Биосинтез белка (как весьма энергоемкий процесс) снижается. Это особенно влияет на синтез тех белков, которые имеют короткое время полужизни, например, факторы свертывания. Концентрация аминокислот в крови повышается. Увеличивается содержание аммиака, снижается количество глутамина, устанавливается отрицательный азотистый баланс.
Нарушения выделения и конечных этапов метаболизма аминокислотОдним из нарушений метаболизма аминокислот в организме является повышенное выделение их с мочой – гипераминоацидурии. Они могут быть следствием повышенного содержания аминокислот в крови, когда фильтрация аминокислот превышает возможность их реабсорбции. В этих случаях возникает генерализованная аминоацидурия, наблюдающаяся при белковом голодании, раневом истощении, тяжелых ожогах, кахектической стадии злокачественных новообразований, болезни Иценко–Кушинга и т.д. В развитии этих нарушений имеет значение относительная недостаточность процессов дезаминирования аминокислот в печени.
Гипераминоацидурии, развивающиеся вследствие нарушения реабсорбции в почечных канальцах, могут носить и наследственный характер. Генерализованная гипераминоацидурия наблюдается при синдроме Фанкони, для которого характерно усиление выведения аминокислот с мочой в сочетании с гиперфосфатурией и глюкозурией. При гепатоцеребральной дистрофии значительная экскреция аминокислот носит вторичный характер и связана с тем, что избыток выделяющейся меди образует в почках комплексные соединения с аминокислотами, которые не могут подвергаться реабсорбции.
Может наблюдаться также и избирательное снижение реабсорбции отдельных аминокислот. Примером может служить цистинурия, при которой нарушение реабсорбции цистина, обусловленное поражением транспортной системы диаминомонокарбоновых кислот, сочетается с повышенным выделением аргинина, лизина и орнитина. Ведущим в клинической картине этого заболевания является нефролитиаз – образование камней почки и мочевыводящих путей вследствие плохой растворимости цистина.
Аминокислоты в организме расщепляются до конечных продуктов. При этом углеродный скелет в общих путях катаболизма – цикле трикарбоновых кислот и терминальных механизмах биологического окисления распадается до углекислого газа и вода, а азотсодержащие функциональные группы – до конечных продуктов азотистого обмена: аммиака и мочевины. Процесс дезаминирования аминокислот осуществляется во всех органах и тканях. Синтез мочевины протекает в основном в печени. Транспортной формой аммиака в организме являются амиды дикарбоновых кислот: глутаминовой и аспарагиновой – глутамин и аспарагин. Азотистые группировки этих соединений являются поставщиками азота для синтеза мочевины в печени и процесса аммониогенеза в почках. Таким образом, ведущую роль в развитии нарушений конечных этапов метаболизма аминокислот играют патологические процессы печени и почек.
Индикаторным показателем, способным охарактеризовать состояние конечных этапов обмена аминокислот и адекватность функций печени к почек по обеспечению этих процессов, является остаточный азот крови. Наибольшей информативностью обладают не интегральные изменения этого показателя, то есть – азотемия – избыточное содержание всех азотсодержащих компонентов крови, а изменения содержания каждого из компонентов остаточного азота.
Нарушение функций гепатоцитов сказывается на активности процессов дезаминирования и трансаминирования аминокислот. Это проявляется в увеличении аминного компонента остаточного азота. Параллельно может наблюдаться нарушение мочевинообразовательной функции печени. В этих случаях увеличение содержания азота аминокислот, глутамина и аммонийных солей в крови сопровождается уменьшением концентрации мочевины. Возможно развитие изолированных нарушений мочевинообразовательной функции при наследственной недостаточности ферментов орнитинового цикла.
Наиболее частым следствием нарушения синтеза мочевины является накопление в крови аммиака, обладающего выраженным цитотоксическим эффектом, особенно для клеток нервной системы. В организме сформирован ряд компенсаторных процессов, обеспечивающих связывание аммиака. Установлено, что 80% аммиака, поступающего из кишечника через аортальную вену в печень, превращается в мочевину, четвертая часть образующейся мочевины секретируется в кишечник, остальное количество выводится почками. Аммиак, не прошедший через орнитиновый цикл, в присутствии глутаминсинтетазы и АТФ превращается в глутаминовую кислоту, а затем – в глутамин. Глутамин переносится кровью в печень и почки, где гидролизуется глутаминазой до свободного аммиака, превращающегося затем в мочевину или связывающегося с ионами водорода и выделяющегося в виде солей аммония с мочой. Последний процесс, названный аммониогенезом, является важным механизмом, направленным на поддержание кислотно-щелочного равновесия. Таким образом осуществляется цикл глутамина, представляющего собой одну из форм транспорта и резерва аммиака.
Даже при небольшом содержании ионов NH4 в крови, они оказывают вредное действие, так как их токсичность зависит от рН среды. Установлено, что степень проникновения аммиака через гематоэнцефалический барьер прямо пропорциональна рН крови. По теории "неионной диффузии" аммиак диффундирует из пространства с более высоким рН в пространство с менее высоким рН, что способствует его проникновению внутрь мозговых клеток. Считают, что ионы аммония снижают уровень α–кетоглутаровой кислоты, нарушая реакции цикла Кребса, угнетая тканевое дыхание. Наряду с уменьшением образования энергии в мозге, возможно и повышение ее распада под действием АТФ–азы, активность которой возрастает при небольших дозах аммиака. Существуют и другие объяснения церебротоксического действия аммиака: образование или накопление в мозге гипотетического вещества, тормозящего передачу нервных импульсов (гамма–аминобутират), нарушение включения в цикл Кребса пировиноградной кислоты, недостаток ацетилхолина, прямое токсическое действие аммиака на клеточные мембраны, нарушение транспорта ионов, снижение внутриклеточного содержания калия, что тесно связано с активностью транспортной АТФ–азы и содержанием ATФ.
Таким образом, аммиак повреждает метаболические процессы в мозговой ткани, приводя к истощению энергетические ресурсы, расходуемые на его обезвреживание, что влечет за собой падение электрической активности клеток мозга и развитие психомоторных нарушений.
Наиболее выраженная азотемия наблюдается при нарушении выделительной функции почек. При этом содержание остаточного азота повышается главным образом за счет азота мочевины. Максимальная степень выраженности этого нарушения проявляется уремией. Уремия возникает при острой или хронической почечной недостаточности.
Общая активация катаболических реакций белкового обмена в организме приводит к равномерному повышению содержания всех компонентов остаточного азота. При этом повышение содержания аминокислот – гипераминоацидемия – сопровождается потерей их с мочой. Это наблюдается при преобладании катаболических гормонов – глюкокортикоидов, тироксина; при голодании, тяжелых хронических заболеваниях.
Литература основная1. Патологическая физиология / Под ред. А.Д.Адо и Л.М.Ишимовой. – К.: Медицина, 1980. – С. 233–239, 310–312.
2. Патологическая физиология / Под ред. Н.Н.Зайко. – Киев: Вища школа, 1985. – С. 265–272.
3. Овсянников В.Г. Патологическая физиология (типовые патологические процессы). Учебное пособие.– Изд–во Ростовского университета, 1987. – С. 60–66.
4. Вапцаров И., Иомтов М., Савов С., Дюкмеджиев И., Эшкенази М. Диспротеинемии. – София: Медицина и физкультура, 1978. –336 с.
5. Мак–Мюррей У. Обмен веществ у человека. – М.: Мир, 1980. – 366 с.
6. Мусил Я. Основы биохимии патологических процессов.– M.: Медицина, 1985. – 430 с.
7. Вельтищев Ю.Е., Ермолаев М.В., Ананенко А.А., Князев Ю.А. Обмен веществ у детей. – М.: Медицина, 1983. – 462 с.
Похожие работы

... клетки, ткани и органы тела человека. Белки составляют основу гормонов, ферментов, антител и других образований, выполняющих сложные функции в жизни человека (пищеварение, рост, размножение, иммунитет и др.), способствуют нормальному обмену в организме витаминов и минеральных солей. Белки участвуют в образовании энергии, особенно в период больших энергетических затрат или при недостаточном ...
... с различной чувствительностью к глюкозе. Это еще один элемент глюкозосенсорного механизма: при высокой концентрации глюкозы увеличивается число клеток, секретирующих инсулин. НАРУШЕНИя метаболизма углеводов и липидов при сахарном диабете При сахарном диабете инсулин-глюкагоновый индекс снижен. Это связано не только с уменьшением секреции инсулина, но и с увеличением секреции глюкагона ( ...
... в такой последовательности: поглаживание, разминание, потряхивание, растирание, движения, ударные приемы. Заканчивается массаж поглаживанием. 2. Физическая реабилитация при подагре и сахарном диабете Подагра – одно из наиболее часто встречающихся нарушений белкового обмена. Это общее заболевание организма, характеризующееся отложением мочекислых солей в сухожилиях, слизистых сумках, ...
... селезенки, печени, лимфатических узлов и костного мозга вследствие генетически предопределенного дефекта фермента гликоцереброзидазы. Прежде чем перейти к рассмотрению частных вопросов нарушений липопротеидного обмена и атеросклероза необходимо остановиться на некоторых общих положениях метаболизма холестерина а организме, характеристике состава и образования транспортных форм липидов крови. ...
0 комментариев