ХАРКІВСЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ ІМЕНІ В.Н. КАРАЗІНА
Гузь Людмила Василівна
УДК 612.822:612.015.3:612.018:577.175.4-008.61]-092.9
НЕЙРОМЕТАБОЛІЧНІ ОСОБЛИВОСТІ ТА ІНТЕГРАТИВНА ДІЯЛЬНІСТЬ ЦЕНТРАЛЬНОЇ НЕРВОВОЇ СИСТЕМИ
ЗА УМОВ ЕКСПЕРИМЕНТАЛЬНОЇ ГІПЕРТИРОКСИНЕМІЇ
У ЩУРІВ
03.00.13 – фізіологія людини і тварин
АВТОРЕФЕРАТ
дисертації на здобуття наукового ступеня
кандидата біологічних наук
Харків – 2008
Дисертацією є рукопис.
Робота виконана в Дніпропетровській державній медичній академії Міністерства охорони здоров'я України
Науковий керівник: доктор медичних наук, профессор Неруш Петро Опанасович, Дніпропетровська державна медична академія, професор кафедри фізіології;
Офіційні опоненти: доктор біологічних наук, профессор Воробйова Тамара Михайлівна, Харківський інститут неврології, психіатрії та наркології АМН України, завідувач лабораторії нейрофізіології та імунології;
доктор біологічних наук, профессор Павленко Володимир Борисович, Таврійський національний університет ім. В.І. Вернадського, професор кафедри фізіології і біофізики
Захист відбудеться “ __ ” ______ 2008 року о _____ годині на засіданні спеціалізованої вченої ради Д 64.051.17 Харківського національного університету імені В.Н. Каразіна Міністерства освіти і науки України за адресою: 61077, м. Харків, пл. Свободи, 4, ауд. 3-15.
З дисертацією можна ознайомитись у Центральній науковій бібліотеці Харківського національного університету імені В.Н. Каразіна Міністерства освіти і науки України за адресою: 61077, м. Харків, площа Свободи, 4.
Автореферат розісланий “ ___“ ________ 2008 року
Учений секретар
спеціалізованої вченої ради Дзюба В.М.
ЗАГАЛЬНА ХАРАКТЕРИСТИКА РОБОТИ
Актуальність теми. Дослідження інтегративної діяльності центральної нервової системи (ЦНС) привертають особливу увагу багатьох дослідників в усьому світі (Дмитриева Н.И., 1982; Ведяев Ф.П., Воробьева Т.М., 1983; Крауз В.А., 1988; Батуев А.С., 1990; Miyamoto S., Teramoto H., 1995).
Такий прояв вищої діяльності мозку як поведінка і когнітивні функції забезпечуються та контролюються нейрогуморальною системою регуляції. Гормонам відводиться роль дифузної нейромодулюючої системи завдяки їх геномному та позагеномному характеру дії на метаболізм клітин. Важливу роль в цьому відіграють тиреоїдні гормони (ТГ), яким відводиться домінуюча роль в процесі формування мозку та підтриманні його функціональної активності впродовж всього життя (Верещагина П.В., 1984; Дривотинов Б.В., 1989; Браверманн Л.И., 2000). Тому комплексне дослідження фізіологічних та біохімічних показників ЦНС за умов метаболічних розладів набуває ще більшої актуальності і свідчить про важливість даної проблеми (Ашмарин И.П., 1987; Недзвецький В.С. и др., 1999; Менджерицкий А.М., 2003; Соболев В.И., 2002; 2004).
Стан фізіологічних функцій організму, його життєдіяльність і адаптація до різноманітних змін навколишнього середовища контролюються нейрогуморальними механізмами (Ашмарин И.П., 1987). Гормональні порушення глибоко змінюють діяльність головного мозку і окремих його структур, що призводить до порушень психічних функцій. Важливим аспектом проблеми нейрогуморальної регуляції функцій є вивчення впливу гормональних факторів на когнітивні процеси головного мозку за умов порушення тиреоїдного статусу (Ettigi P.Y. et al., 1977; Городецкая И.В. и др., 2000; Fedotova J.O., 2000; Сапронов Н.С., 2002; Фадеев В.В., 2006).
Незважаючи на те, що за останні десятиріччя дослідниками різних країн зібрано надзвичайно великий матеріал відносно впливу ТГ на вищу нервову діяльність, на сьогодні відомі різні, відмінні експериментальні дані про можливий вплив ТГ на процес навчання і пам¢яті (Бойчук Е.Б., 2000; Сапронов Н.С. и др., 2002). В той же час молекулярні механізми, які обумовлюють функціональні порушення нервової системи, залишаються вивченими недостатньо. Результати, які представлені у сучасній літературі, є розрізненими, трактуються неоднозначно, тому не можуть бути складені у загальну концепцію. Порушення регуляції активності ендокринної системи, а також лабільності відповіді секреторних клітин на зовнішні фактори є однією з важливих причин, що ведуть до функціонального розладу у нервовій системі, порушень метаболічної рівноваги нервових клітин (Бунявичус Р., 1994). Головним чином це обумовлено погіршенням екологічної ситуації, що веде до постійного зростання кількості несприятливих чинників, а з другого боку – постійними стресовими ситуаціями, генетичною схильністю, які різними шляхами впливають на біохімічні та молекулярні процеси в клітинах нервової системи, що супроводжується порушенням психічних функцій (Тараканова Н.П., 1994; Тайулин В.И., 1998; Тананакіна М.В., 2006). Цей аспект практично залишається відкритим. Тому є доцільним дослідження особливостей нейрометаболічних процесів у функціонуванні ЦНС за умов експериментальної гіпертироксинемії. У зв¢язку зі згаданим вище, поглиблене вивчення механізмів порушення нейрометаболічних процесів може сприяти розробці раціональних методів корекції патологічних процесів за умов гіпертироксинемії.
Зв¢язок роботи з науковими програмами, планами, темами. Робота являється фрагментом планової науково-дослідної роботи кафедри фізіології Дніпропетровської державної медичної академії "Вивчення механізмів компенсаторно-адаптивних реакцій нервової системи при дії екстремальних факторів", затвердженої відділом держреєстрації УкрINTEI (м. Київ) під номером 0100U000307.
Мета і завдання дослідження. Визначити особливості нейрометаболічних процесів та поведінкову діяльність у білих щурів за умов експериментальної гіпертироксинемії.
У відповідності з метою визначено наступні завдання:
1.Вивчити рівень поведінкової активності та мнестичні реакції щурів за умов експериментальної гіпертироксинемії.
2.Дослідити параметри збудливості моносинаптичної рефлекторної дуги та оцінити вплив на них тривалої тетанізації, вивчити поріг і хронаксію еферентних волокон сідничного нерва (СН) за умов експериментальної гіпертироксинемії.
3.Дослідити стан прооксидантних процесів в різних за функціональним навантаженням структурах головного мозку щурів (кора головного мозку, гіпокамп, стовбур головного мозку) за умов експериментальної гіпертироксинемії.
4.Дослідити активність ферментів антиоксидантної системи (глутатіонпероксидази та каталази) структур головного мозку (кора головного мозку, гіпокамп, стовбур головного мозку) щурів за умов експериментальної гіпертироксинемії.
5.Визначити вміст окремих вільних жирних кислот (ЖК) в ліпідах головного мозку (кора головного мозку, гіпокамп, стовбур головного мозку) щурів за умов експериментальної гіпертироксинемії.
Об¢єкт дослідження – показники поведінкової та когнітивної діяльності щурів, рівень активності прооксидантної та антиоксидантної систем в окремих структурах головного мозку, вміст ЖК ліпідів кори, гіпокампу та стовбуру мозку.
Предмет дослідження – особливості нейрометаболічних процесів та поведінкова діяльність щурів за умов експериментальної гіпертироксинемії.
Методи дослідження. З метою дослідження інтегративних реакцій ЦНС за умов експериментальної гіпертироксинемії були застосовані фізіологічні методи: "відкрите поле" – для визначення поведінкової активності щурів; умовна реакція пасивного уникнення (УРПУ) – для визначення когнітивних функцій; електрофізіологічні дослідження – визначення параметрів збудливості моносинаптичної рефлекторної дуги та характеру посттетанічної потенціації (ПТП), вивчення порогу та хронаксії еферентних волокон СН. Показником функціональної активності клітин окремих структур мозку являлося дослідження стану прооксидантної та антиоксидантної систем, вмісту ЖК в ліпідах кори, гіпокампу та стовбуру мозку. Модель гіпертироксинемії створювалась шляхом введення з їжею L-тироксину (10 – 140 мкг/доба) впродовж двох тижнів. Після створення експериментальної гіпертироксинемії в плазмі крові піддослідних тварин визначався рівень тироксину. Всього було поставлено 1080 дослідів на 120 щурах.
Наукова новизна одержаних результатів. Вперше шляхом комплексного дослідження показано, що зміни поведінкової діяльності та мнестичної активності при експериментальній гіпертироксинемії супроводжувались істотним зростанням збудливості моносинаптичних рефлекторних дуг, свідченням чого є збільшення амплітуди моносинаптичної рефлекторної відповіді (МРВ), зменшення латентного періоду та порогу виникнення моносинаптичної рефлекторної реакції, що вказує на стан пластичності синапсів і може призводити до порушення інтегративної діяльності ЦНС. Показано, що за умов експериментальної гіпертироксинемії тетанізація призводила до депресії рефлекторних відповідей. На основі комплексного аналізу роботи, яка включає фізіологічний та біохімічний рівень дослідження, була встановлена залежність між функціональними процесами ЦНС та структурними клітинними перебудовами метаболічного характеру. Показано, що експериментальна гіпертироксинемія супроводжувалась змінами балансу прооксидантної та антиоксидантної систем у вивчаємих структурах головного мозку залежно від рівня ТГ.
Практичне значення одержаних результатів. Робота є експериментальним дослідженням, яке розширює сучасні уявлення відносно механізмів порушення поведінкової та умовно-рефлекторної діяльності за умов експериментальної гіпертироксинемії. Вивчені особливості нейрометаболічних процесів за умов підвищеного рівня ТГ. Результати досліджень можуть бути передумовою для розробки методів корекції порушень інтегративних функцій ЦНС за умов дисфункції щитоподібної залози. Отримані факти можуть бути використані в практичній фізіології та гігієні праці при розробці засобів та методів підвищення стійкості організму до струмогенних факторів навколишнього середовища та фізичних навантажень; в клінічній практиці – для корекції нервово-психічних розладів за умов порушеного гіпоталамо-гіпофізарно-тиреоїдного статусу, в клінічній ендокринології та патофізіології – при виборі засобів та методів фармакокорекції лікарськими засобами, які впливають на пластичність синаптичних утворень, порушеного тиреоїдного статусу за умов тиреоїдної патології. В навчальному процесі ці дані можуть бути використані при вивченні фізіології вищої нервової діяльності, ендокринної системи, патофізіології. Результати дисертаційної роботи впроваджені в навчальний процес кафедри фізіології та кафедри патофізіології Дніпропетровської державної медичної академії, кафедри фізіології людини та тварин Дніпропетровського національного університету, кафедри фізіології Вінницького національного медичного університету ім. М.І. Пірогова, кафедри фізіології Львівського державного національного медичного університету ім. Данила Галицького.
Особистий внесок здобувача. Проведено патентно-інформаційний пошук і аналіз наукової літератури з теми дисертаційної роботи, проведені експериментальні дослідження на щурах. Дисертант особисто проводив статистичну обробку отриманих даних, узагальнив результати дослідження, сформулював основні висновки, написав всі розділи дисертації, підготував матеріали до публікації. У дисертації не використані ідеї та розробки, що належать співавторам опублікованих наукових праць.
Апробація результатів дисертації. Матеріали досліджень, що включені до дисертації, викладені автором на наукових семінарах кафедри фізіології Дніпропетровської державної медичної академії (Дніпропетровськ, 2001 – 2007), у доповідях на наукових форумах різних рівнів: VII та IX міжнародні конференції “Центральные и периферические механизмы вегетативной нервной системы», присвяченій пам¢яті О.Г. Баклаваджяна (Донецьк, 2003, 2007), наукова конференція «Эколого-физиологические проблемы адаптации» (Партенит, 2003), V Українська конференція молодих вчених, присвячена пам¢яті ак. В.В. Фролькіса (Київ, 2004), наукова конференція «Нейронауки: теоретичні та клінічні аспекти» (Донецьк, 2005), II Міжнародна наукова конференція “Гомеостаз: фізіологія, патологія, фармакологія і клініка” (Одеса, 2005), IV Національний конгрес геронтологів і геріатрів України (Київ, 2005), Всеукраїнська науково-практична конференція “Проблеми вікової фізіології” (Луцьк, 2005), Всеукраїнська науково-практична конференція “Довкілля і здоров¢я” (Тернопіль, 2006), XVII з¢їзд Українського фізіологічного товариства з міжнародною участю пам¢яті академіка О.О. Богомольця (Чернівці, 2006), III Всеукраїнська наукова конференція, присвячена 70-річчю з дня народження Г.М. Чайченка (Київ, 2006), Всеукраїнський науковий симпозіум “Особливості формування та становлення психофізіологічних функцій в онтогенезі” (Черкаси, 2006), III з`їзд фармакологів Росії «Фармакология – практическому здравоохранению» (Санкт-Петербург, 2007).
Публікації. За темою дисертації опубліковано 21 наукових робіт, з них 5 статей у фахових наукових виданнях, та 16 тез у матеріалах з¢їздів, симпозіумів та конференцій.
Структура та обсяг роботи. Робота викладена на 144 сторінках печатного тексту і складається з введення, огляду літератури, трьох розділів об'єкту і методів дослідження, сьоми розділів власних досліджень, заключення, висновків, переліку використанної літератури (315 джерел, з них 141 – іноземні, включно до 2007 року). Робота ілюстрована 14 рисунками і 11 таблицями.
ОСНОВНИЙ ЗМІСТ РОБОТИ
Матеріали та методи дослідження. Об`єктом дослідження були білі щури лінії Вістар різної статі віком 5 – 6 місяців, маса тварин в середньому становила 150 – 250 грамів. Було поставлено 1080 дослідів на 120 тваринах. Моделювання гіпертироксинемії проводили шляхом введення з їжею перемелених таблеток L-тироксину („Berlin-Chemie” АУ, Німеччина). Всі тварини знаходились на стандартному харчовому раціоні віварію. Формування гіпертироксинемії супроводжувалось зниженням маси тварин, підвищенням їх агресивності, збільшенням частоти серцевих скорочень в порівнянні з вихідним станом, і проводилось за методикою, апробованою на кафедрі фізіології (Макий Е.А. и др., 2000; 2002). В дослідженні використовували зростаючі дози L-тироксину. На початку вводилась доза препарату, яка була вищою від добової продукції тироксину (в нормі біля 3 – 5 мкг/доба) в декілька разів, звичайно брали дозу 10 мкг/доба. Далі щоденно, впродовж двох тижнів, збільшували дозу в середньому на 10 мкг/доба, так що під кінець досліду тварини отримували біля 140 мкг препарату на добу. Через два тижні після початку формування моделі гіпертироксинемії імуноферментним методом визначали концентрацію тироксину (Т4), використовуючи набір реактивів та апаратуру фірми Humareader (Німеччина). Вміст тироксину в сироватці крові у тварин з гіпертироксинемією зростав в 5 разів.
В другій серії експериментів (n=92) вивчалась орієнтовно-дослідницька діяльність методом „відкритого поля” (Ведяев Ф.П. и др.,) і УРПУ (1983Буреш Я. и др., 1991).
У третій серії дослідів проводились дослідження МРВ. В гострому досліді під наркозом проводили ламінектомію в сегментах L1 – L6 СМ. Виділяли та перерізали вентральний та дорсальний корінці сегменту L5 у місці їх відходження у міжпозвонковий отвір. Через 3 – 4 години після ламінектомії приступали до відведення біопотенціалів. Подразнення здійснювали прямокутним електричним імпульсом тривалістю 0,3 мс. Сила подразнення була надпороговою для виникнення МРВ і складала 2 пороги. Для вивчення феномену ПТП дорсальний корінець подразнювали пачкою з 5000 стимулів частотою 300 Гц.
Параметри МРВ – амплітуда, латентний період, поріг виникнення, зміни його амплітуди при зростаючій силі подразнень, характер ПТП – порівнювали з такими ж параметрами у інтактних тварин, отриманих іншими дослідниками (Гайдина Г.А., 1970; Мантуло П.М., 1980; Макій Є.А. та ін., 2000; 2002).
У четвертій серії дослідів (n=18) проводилось визначення вмісту вільних ЖК кислот в структурах головного мозку (кора великих півкуль, гіпокамп, стовбур мозку) методом Фолча (Прохорович М.И., 1982). Естерифіковані ЖК розчиняли в 1 мл хлороформу і визначення їх вмісту (в мкмоль/г тканини мозку) проводили на хроматографі CHROM-5 в ізотермічному режимі з полум`яно-іонізуючим детектором на колонці 5Ч1000 мм із сорбентом хромотон-N-super 5% SE-2100. ЖК виявляли в температурному діапазоні від 185 до 250°С при лінійному безперервному підвищенні температури на 5°С за хвилину. Як внутрішні стандарти використовували набір етилових ефірів ЖК С ("Sigma" США).
В п`ятій серії дослідів (n=25) визначався стан прооксидантної системи в корі головного мозку, гіпокампі і стовбуровому відділі мозку шляхом вивчення вмісту як первинних продуктів ПОЛ – дієнових кон'югатів (ДК), так і вторинних на основі визначення малонового діальдегіду (МДА) за методами (Стальная И.Д., 1979; Гаврилов В.Б. и др., 1983).
В шостій серії дослідів (n=25) досліджували стан антиоксидантної системи шляхом визначення активності каталази і глутатіонпероксидази в вивчаємих структурах головного мозку (Королюк М.А., 1988; Мальцев Г.Ю. и др., 2002).
Дані експериментів оброблювали методами варіаційної статистики та за допомогою програмного забезпечення Microsoft Excel, Statistica 6,0. Вірогідність отриманих результатів оцінювали за допомогою критерію t-Ст'юдента для параметричних та непараметричних методів (Лакін Г.В., 1990).
РЕЗУЛЬТАТИ ДОСЛІДЖЕНЬ ТА ЇХ ОБГОВОРЕННЯ
Стан умовно-рефлекторної і поведінкової діяльності щурів за умов експериментальної гіпертироксинемії. Метою першого напрямку наших досліджень було вивчення стану мнестичних (умовно-рефлекторних) функцій та орієнтовно-дослідницької діяльності у щурів за умов гіпертироксинемії (табл. 1).
Таблиця 1Порівняльна характеристика умовної реакції пасивного уникнення у інтактних щурів та за умов експериментальної гіпертироксинемії
| Група тварин | Кількість амнезованих, % | Латентний період заходу в темний відсік, с (M±m) |
| контроль (n = 35) | 43 | 100,60 ± 9,60 |
| гіпертироксинемія (n = 45) | 78* | 55,60 ± 9,60* |
Примітка. * - вірогідність результату у порівнянні з відповідним контролем при рівні достовірності р<0,05.
Як свідчать факти, експериментальна гіпертироксинемія супроводжувалась порушенням інтегративної діяльності ЦНС, стан якої оцінювали на основі визначення поведінково-дослідницької діяльності тварин методом „відкритого поля”, та умовно-рефлекторної діяльності визначенням УРПУ. Тварини були з приблизно однаковою емоційною характеристикою, що визначалось перед дослідом на основі тесту „відкрите поле”.
У тварин з гіпертироксинемією в порівнянні з інтактними тваринами горизонтальна рухливість збільшувалась на 26% (р<0,01) (рис. 1). Орієнтовно-дослідницька активність характеризувалась тенденцією до підвищення, що проявлялось в зростанні кількості обстежених "нірок" від 5,07±0,38 в групі інтактних тварин до 6,15±0,36 в експерименті. Показники емоційно-вегетативного реагування (реакція „грумінг” та кількість болюсів) змінювалась різноспрямовано. Тривалість грумінгу у щурів була на рівні контролю, а кількість болюсів дефекації була зменшена на 61% (р<0,05).
Щодо вивчення мнестичних реакцій за умов гіпертироксинемії – отримані результати дають підставу говорити про погіршення показників умовно-рефлекторної діяльності. Нами встановлено, що кількість амнезованих тварин в інтактній групі становила 43% (р<0,05) (табл. 1). Разом з тим у дослідних тварин з (експериментальною гіпертироксинемією) амнезовані тварини становили 78% (р<0,05). Латентний період реакції тестування навику УРПУ у дослідних тварин (перехід з світлого в темний відсік) зменшувався на 55,6% (р<0,05).
Таким чином, за матеріалами розділу було зроблено висновок, що в дослідженнях мнестичних (умовно-рефлекторних) реакцій і орієнтовно-дослідницької активності (рис. 1) за умов експериментальної гіпертироксинемії стану має місце порушення інтегративної діяльності ЦНС, що виявляється як з боку пізнавальних, так і мнестичних (умовно-рефлекторних) показників. Амнестичний тип умовно-рефлекторної діяльності в експерименті проявляється феноменом ретроградної амнезії, для формування якої широко використовується УРПУ електричного струму. Ця реакція формується в одній пробі. УРПУ представляє собою відносно просту оборонну реакцію уникнення аверсивного струму порогового значення. Вона дає можливість досліджувати основні закономірності формування, зберігання і репродукції умовних рефлексів. Феномен ретроградної амнезії дає можливість, як видно із нашого дослідження, характеризувати зміни пам'яті, які проходять з виробленою навичкою.
Це дає підстави заключити, що тиреоїдні гормони, зокрема Т4, відіграють певну роль в умовно-рефлекторній діяльності. Як показано нами, це стосується як зберігання УРПУ, так і орієнтовно-дослідницької діяльності за умов експериментальної гіпертироксинемії. Тому наступні наші дослідження були присвячені вивченню можливих механізмів порушення цих функцій.
![]()
![]()
![]()
![]()
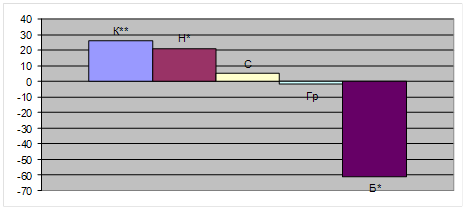
Рис. 1. Показники орієнтовно-дослідницької діяльності щурів у "відкритому полі" за умов експериментальної гіпертироксинемії.
Примітки: *- вірогідність результату у порівнянні з контролем при рівні достовірності р<0,05; ** - вірогідність результату у порівнянні з контролем при рівні достовірності р<0,01.
Сегментарні рефлекторні реакції за умов експериментальної гіпертироксинемії та вплив на них тривалої тетанізації. Результати вище означених дослідів можна проаналізувати за допомогою визначення впливу надмірної кількості тироксину на функціонування найпростішої рефлекторної моносинаптичної дуги спинного мозку.
Результати дослідження параметрів МРВ на поодиноке подразнення силою два пороги (2П) наведено в таблиці 2.
Таблиця 2
Параметри моносинаптичних рефлекторних відповідей у інтактних щурів та шурів за умов системної дії тироксину при стимуляції дорсальних корінців
| Групи тварин | Параметри МРВ | ||
| Амплітуда (мВ) | Поріг (мкА) | Латентний період (мс) | |
| 1. Інтактні тварини n=13 | 1,80±0,07 | 2,40±0,10 | 1,23±0,10 |
| 2. Тварини з гіпертироксинемією n=10 | 3,87±0,27 р<0,001 | 0,64±0,05 р<0,001 | 0,78±0,05 р<0,01 |
Встановлено, що за умов експериментальної гіпертироксинемії зростала збудливість моносинаптичних рефлекторних дуг. Амплітуда МРВ збільшувалась більше, ніж у 2 рази. При цьому поріг виникнення МРВ зменшувався майже в 4 рази, а латентний період зменшився в 1,5 рази.
Додатковим доказом збільшення збудливості мотонейронів в експерименті є дані щодо зростання амплітуди МРВ при збільшенні сили стимуляції від 1,2 до 2П. Ці дані систематизовано в таблиці 3.
Аналіз результатів, які наведено в таблиці 3, вказує, що зростання амплітуди МРВ відбувається значно скоріше у тварин на фоні гіпертироксинемії.
Таблиця 3
Зміни амплітуди МРВ у інтактних щурів та у щурів за умов системної дії тироксину при зростанні сили стимуляції дорсальних корінців від 1,2 до 2,0 П (% до амплітуди МРВ при подразненні стимулом в 2 П)
| Групи тварин | 1,2 П | 1,4 П | 1,6 П | 1,8 П | 2,0 П |
| 1. Інтактні тварини n=13 | 53,3±6,9 | 72,4±7,2 | 95,2±2,0 | 100,00± 1,00 | 100,00± 1,00 |
| 2. Тварини з гіпертироксинемією n=10 | 87,70± 7,20 р<0,01 | 96,80± 2,10 р<0,01 | 100,10± 0,60 р<0,05 | 100,00± 1,50 p>0,05 | 100,00± 1,00 р>0,05 |
Подальший етап досліджень був пов`язаний з вивченням характеру змін ПТП за умов експериментальної гіпертироксинемії. Показано, що тривала тетанізація дорсального корінця при його подразненні пачкою з 5000 стимулів з частотою 300 Гц супроводжувалась підсиленням амплітуди МРВ упродовж майже 5 хв. У експериментальних тварин, навпаки, спостерігалась виражена депресія амплітуди МРВ (рис. 2).
В усіх випадках між групами інтактних тварин та тварин з гіпертироксинемією існує різниця з достовірністю р<0,001. Як видно з рис. 2, тривала тетанізації призводить до виразного підсилення амплітуди МРВ на протязі щонайменше 5 хв. (крива а). У випадку ж вивчення цього процесу у тварин з гіпертироксинемією, замість підсилення спостерігається виразна депресія амплітуди МРВ – на протязі 5 хв. вона навіть підсилюється (крива б).
Також додатково було з'ясовано, що в експерименті поріг і хронаксія еферентних волокон СН змінювались. У експериментальних тварин поріг винекнення потенціала дії в дистальній ділянці вентрального корінця (ВК) L5 підвищувався, а хронаксія була істотно зменшеною.
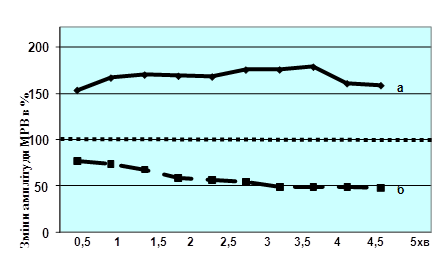
Рис. 2. Зміни амплітуди моносинаптичної рефлекторної відповіді вентрального корінця сегменту L5 спинного мозку білих щурів після нанесення подразнення пачкою в 5000 стимулів з частотою 300 Гц на дорсальний корінець цього ж сегменту.
Примітки: а – інтактні тварини (n=13); б – тварини з експериментальною гіпертироксинемією (n=10); за 100 % прийнята амплітуда МРВ на подразнення силою в 2 П до проведення тетанізації.
Вивчення метаболізму ліпідів в структурах головного мозку. Нами вивчено обмін фосфоліпідів, які являються головними компонентами біологічних мембран і мієліну. Стан мембран являється одним з найважливіших факторів підтримання гомеостазу та регуляції біохімічних та фізіологічних процесів у клітинах (Владимиров Ю.А. и др., 1972; Воскресенский О.Н., 1987; Богацкая Л.Н. и др., 1990; Бурлакова Е.Е., 1990). Фосфоліпідні фракції включають як насичені, так і ненасичені ЖК. Присутність вільних ЖК в тканинних системах, зокрема в головному мозку, інтенсивно впливає на перебіг в них метаболічних процесів.
Проведений нами газохроматографічний аналіз складу ЖК ліпідів мозку показав, що в філогенетично різних відділах головного мозку вміст вільних ЖК відрізнявся. В корі головного мозку інтактних щурів рівень насичених кислот (пентадеканової і пальмітинової) був більш високим. В стовбурі головного мозку звертало увагу підвищення рівня насиченої стеаринової кислоти (С18:0), а також ненасичених олеїнової (С18:1) і арахідонової (С20:4) кислот. В гіпокампі мало місце зростання концентрації олеїнової кислоти (С18:1). Порівняльна характеристика рівня ЖК свідчить, що в стовбурі головного мозку інтактних щурів показники насичених (С14:0, С18:0), а також ненасичених (С18:1, С18:2 і С20:4) ЖК були вище, ніж в гіпокампі і корі головного мозку.
Нами встановлено, що за умов експериментальної гіпертироксинемії змінюється обмін ліпідів в структурах головного мозку. Звертало увагу зниження концентрації насичених ЖК в корі головного мозку, тоді як рівень ненасичених ЖК в спектрі С18:1 - С20:4, навпаки, зростав. Збільшувалась концентрація лінолевої (С18:2) кислоти в 2 рази по відношенню до контрольних тварин.
Основою ліпідного матриксу клітинних мембран являються поліненасичені ЖК, що входять до складу фосфоліпідів і зумовлюють структурні та функціональні властивості мембран. Одна з найважливіших ненасичених ЖК у складі фосфоліпідів клітинних мембран є арахідонова, рівень якої в корі головного мозку зростав на 120%. Вона являється основним джерелом синтезу ейкозаноїдів та одним із головних субстратів ПОЛ.
Дещо несподіваним, з нашої точки зору, являється результат в гіпокампі, який виконує важливу роль в процесах навчання і запам’ятовування. Так, якщо в корі головного мозку за даних умов рівень насичених ЖК (С14:0 – С18:0) знижувався, то в гіпокампі, навпаки, відмічено зростання концентрації ЖК в даному спектрі. При цьому рівень ненасичених кислот в гіпокампі змінювався різноспрямовано. Встановлено зниження рівню олеїнової (С18:1) та арахідонової (С20:4) кислот, та збільшення концентрації лінолевої (С18:2) на 162% (р<0,01) (табл. 4).
Таблиця 4
Динаміка вмісту вільних жирних кислот (мкмоль/г) в структурах головного мозку щурів за умов експериментальної гіпертироксинемії
| Жирні кислоти | Кора головного мозку М±m, % змін | Гіпокамп М±m, % змін | Стовбур мозку М±m, % змін |
| Міристинова С14:0 Пентадеканова С15:0 Пальмітинова С16:0 Стеаринова С18:0 Олеїнова С18:1 Лінолева С18:2 Арахідонова С20:4 | 0,03±0,01 * -57 0,79 ±0,01 * -26 7,68±0,75 +7 7,22±0,62 * -42 14,65±0,53 +8 0,68±0,10 * +100 1,06±0,15 ** +120 | 0,02±0,001 ** +100 0,22±0,02 * +22 6,36±0,67 * +19 8,79±1,13 +15 16,69±1,90 -11 0,76±0,11 ** +162 1,38±0,18 ** -34 | 0,04±0,01 *** -89 0,05±0,004 *** -91 5,81±0,64 0 8,25±1,13* -56 30,96±0,83 -4 0,82±0,04* +64 1,95±0,21 ** -56 |
Примітки: наведено середнє значення та похибка середнього M±m; * - р<0,05; ** - р<0,01; *** - р<0,001 по відношенню до інтактних; n = 9.
Ці факти дають підстави стверджувати, що фосфоліпіди гіпокампу також являються об’єктом впливу ТГ. Це співпадає з більш ранніми дослідженнями (Gen P. et al., 1994) щодо підвищеного рівню ТГ, які стимулюють синтез насичених ЖК і інтенсифікують процеси ПОЛ в тканинах. На думку деяких дослідників (Бурлакова Е.Б. и др., 1980), особливість окиснення ЖК в тканинах заключається в тому, що в першу чергу метаболізуються ненасичені ЖК. Це вказує на зміни рівня ненасичених (С18:1, С20:4) ЖК в гіпокампі.
Аналізуючи обмін ліпідів в стовбурі головного мозку, нами встановлено, що на відміну від кори головного мозку і гіпокампу, більш виражені зміни в спектрі ЖК характерні для насичених кислот – міристинової, пентадеканової і стеаринової. Це проявилось в зниженні їх рівня в порівнянні з інтактними тваринами, тоді як характер змін в спектрі ненасичених ЖК С18:1 – С20:4 був аналогічний з такими в корі головного мозку і гіпокампі. Таким чином, зміни спектру ЖК в різних відділах головного мозку мають певну специфічність.
Будь-які зміни складу ЖК мембрани, у свою чергу, позначаються на функціях рецепторів, які беруть участь у передачі сигналів клітинами, на транспортні білки та ферменти, які розташовуються в ліпідному бішарі мембран, що підтверджено в дослідах на культурах неонатальних кардіоміоцитів.
Слід звернути увагу на ті обставини, що ТГ і катехоламіни являються похідними однієї і тієї ж амінокислоти – тирозину. Все сказане свідчить на користь передбачення про наявність загальних закономірностей в дії катехоламінів і ТГ (Воробьева Т.М., 1972; 1983; Кубарко А.И. и др., 1998).
За рівнем витрачання або накопичення поліненасичених ЖК можна оцінювати ступінь вільнорадикального окиснення ліпідів мозку, що дає можливість характеризувати стан прооксидантної та антиоксидантної систем головного мозку за умов експериментальної гіпертироксинемії.
Стан прооксидантної та антиоксидантної системи в структурах головного мозку щурів за умов експериментальної гіпертироксинемії. На сьогодні особливе значення надається вивченню процесів ПОЛ за наявності нервово-психічних розладів, які мають місце при дисфункції щитоподібної залози. Відомо, що для головного мозку характерний високий рівень ліпідів. Загальна кількість сухої речовини мозку більше, ніж на 50% складається з ліпідів. Головний мозок активно забезпечується киснем і має широкий спектр ферментів, що генерують його активні форми. Таким чином, головний мозок являється тим органом, в якому в значному ступені виконуються умови, необхідні для інтенсивного перебігу процесів ПОЛ. На це дійсно вказують дослідження (Таранова Н.П. и др., 1984; Тараканова Н.П. и др., 1994), якими встановлено, що вміст МДА в тканинах мозку був значно вищим, ніж в периферичній нервовій системі та інших органах і тканинах (таблиця 5).
Отримані нами результати корелюють з більш ранніми фактами (Silva J.E. et al., 1978) відносно того, що в ЦНС виявлена значна каталітична активність ферментів антирадикального захисту – каталази та глутатіонпероксидази. Встановлено також, що антиокисна активність ліпідів в різних відділах головного мозку людини значно вище, ніж в інших органах (Бурлакова Е.Б. и др., 1985) (таблиці 6 та 7).
Таблиця 5
| Структури мозку | Інтактні тварини (n=10) | Гіпертироксинемія (n=15) | ||
| ДК (довж. хвилі) Л 232 | МДА (нмоль/мг білка) | ДК (довж. хвилі) Л 232 | МДА (нмоль/мг білка) | |
| Кора головного мозку Гіпокамп Стовбур мозку | 0,12±0,02 0,095±0,005 0,15±0,02 | 0,52±0,05 0,63±0,08 0,59±0,03 | 0,11±0,008*** -8% 0,13±0,003*** +37% 0,14±0,01*** -7% | 0,75±0,07*** +44% 1,07±0,1*** +70% 0,82±0,06*** +39% |
Порівняльна характеристика вмісту дієнових кон’югатів та малонового діальдегіду в структурах головного мозку при гіпертироксинемії
Примітки: наведено середнє значення та похибка середнього M±m; *** - р<0,001 по відношенню до інтактних.
Таблиця 6
Порівняльна характеристика активності каталази в утвореннях головного мозку за умов гіпертироксинемії
| Структура мозку | Активність каталази (мкат/мг білка) | |
| Інтактні тварини (n=10) | Гіпертироксинемія (n=10) | |
| Кора головного мозку М±m % змін Гіпокамп М±m % змін Стовбур мозку М±m % змін | 34,80±3,30 36,80±3,20 37,10±4,80 | 85,30±5,70 ***+145% 74,30±6,10 ***+102% 76,60±2,90 ***+106% |
Примітки: наведено середнє значення та похибка середнього M±m; *** - вірогідність результату у порівнянні з відповідним контролем при рівні достовірності р<0,001.
Таким чином, слід підкреслити, що експериментальна гіпертироксинемія у щурів характеризувалась активацією вільнорадикального окиснення в структурах головного мозку. Це проявилось зростанням вмісту як первинних продуктів – ДК в гіпокампі, так і, особливо, вторинних продуктів ПОЛ в усіх утвореннях головного мозку.
Таблиця 7
Порівняльна характеристика глутатіонпероксидази в структурах головного мозку щурів з експериментальною гіпертироксинемією
| Структура мозку | Активність глутатіонпероксидази (мкмоль/мг білка) | |
| Інтактні тварини (п = 10) | Гіпертироксинемія (п = 15) | |
| Кора головного мозку Гіпокамп Стовбур мозку | 4,10±0,30 6,30±0,55 5,60±0,50 | 8,30±0,50 *** +102% 3,70±0,50 *** -41% 7,91±0,56 *** +41% |
Примітки: наведено середнє значення та похибка середнього M±m; *** - вірогідність результату у порівнянні з відповідним контролем при рівні достовірності р<0,001.
Відомо, що вільнорадикальні реакції відіграють важливу роль у пластичних перебудовах клітин, необхідних для пристосування до змін внутрішнього та зовнішнього середовища, завдяки тому, що активні форми кисню виконують функцію вторинних месенджерів, здатних викликати експресію генів, модуляцію активності протеїнкіназ і впливати на функціональну активність клітин, що, в свою чергу, може приводити до порушення інтегративної діяльності нервової системи.
Висновки
1. Експериментальна гіпертироксинемія суттєво впливає на інтегративну діяльність ЦНС. Підвищений рівень Т4 в сироватці крові викликав порушення орієнтовно-дослідницької поведінки та умовно-рефлекторних реакцій як з боку пізнавальних, так і мнестичних показників. Також спостерігалось зниження емоційно-вегетативних реакцій.
2. За умов експериментальної гіпертироксинемії при подразненні силою в два пороги суттєво зростає збудливість моносинаптичних рефлекторних дуг, що виявлялось в зменшенні порогу виникнення рефлекторної відповіді майже в 4 рази, латентного періоду в 1,5 рази і збільшенні амплітуди МРВ більше, ніж в 2 рази. Тривала тетанізація дорсального корінця сегменту L5 спинного мозку за умов гіпертироксинемії характеризувалась депресією амплітуди моносинаптичної відповіді вентрального корінця цього ж сегменту. Виявлена значна депресія амплітуди МРВ вентрального корінця сегменту L5 спинного мозку. Одержані дані щодо групи тварин з гіпертироксинемією, відносно контролю, вказують на зниження стабільності та ефективності синаптичної передачі. Прогресивне зниження амплітуди може бути результатом "виснаження" нервових клітин через розлад метаболізму, викликаний надмірною стимуляцією ТГ.
3. Стан прооксидантної системи за умов експериментальної гіпертироксинемії в різних утвореннях головного мозку характеризувався підвищенням рівня дієнових кон’югатів в гіпокампі на 37% (р<0,001) та наростанням концентрації малонового діальдегіду, вміст якого зростав в напрямку стовбур мозку < кора головного мозку < гіпокамп.
4. За умов експериментальної гіпертироксинемії вміст жирних кислот змінювався неоднаково. В корі головного мозку рівень насичених кислот переважно знижувався, а ненасичених зростав, зокрема арахідонової (С20:4), концентрація якої збільшувалась на 120% (р<0,01). В гіпокампі відмічено збільшення концентрації в спектрі насичених жирних кислот. Концентрація ненасичених змінювалась різноспрямовано; рівень лінолевої кислоти (С18:2) зростав на 162% (р<0,01), а арахідонової (С20:4) знижувався на 34% (р<0,05). В стовбурі мозку більш виражені зміни встановлені відносно насичених кислот - міристинової (С14:0), пентадеканової і стеаринової, концентрація яких знижувалась.
5. Активність каталази за умов гіпертироксинемії в структурах головного мозку зростала. Більш високою вона була в корі головного мозку (+145%, р<0,001), в гіпокампі і стовбурі мозку вона збільшувалась, відповідно на 102% і 106% (р<0,001). Зміни активності глутатіонпероксидази характеризувались різноспрямованістю. Найбільш високою вона відмічена в корі головного мозку (+102%, р<0,001), в стовбурі мозку вона зростала на 41% (р<0,001), а в гіпокампі мало місце зниження на 41% (р<0,001). Це підтверджує наявність змін активності антиоксидантної системи в вивчаємих структурах головного мозку.
Список праць, опублікованих за темою дисертації
1. Гузь Л.В. Сравнительная характеристика процессов перекисного окисления липидов в различных отделах головного мозга крыс при гиперфункции щитовидной железы // Таврический медико-биологический вестник. – 2004. – Т. 7, № 1. – С. 140 – 142.
2. Макій Є.А., Родинський О.Г., Гузь Л.В. Характер моносинаптичних рефлекторних реакцій спинного мозку білих щурів за умов системної дії тироксину та 4-амінопіридину // Експериментальна та клінічна фізіологія і біохімія. – Львів, 2004. – № 2 (26). – С. 16 – 20. (Здобувач провела експериментальну частину роботи, пов'язану із дослідженням МРВ; вивчались параметри МРВ – амплітуда, латентний період, поріг виникнення, зміни його амплітуди при зростаючій силі подразнень, ПТП).
3. Демченко О.М., Гузь Л.В. Порівняльна характеристика жирнокислотного складу ліпідів головного мозку щурів у процесі онтогенезу // Вісник Луганського національного педагогічного університету ім. Т. Шевченка. – 2005. – № 6 (86). – С. 48 – 50. (Здобувач провела експериментальну частину роботи, пов'язану із визначенням вмісту вільних жирних кислот в структурах головного мозку).
4. Гузь Л.В., Неруш П.О., Демченко О.М. Інтегративна діяльність центральної нервової системи за умов дисфункції щитоподібної залози // Вісник проблем біології і медицини. – 2004. – Вип. 3. –– С. 9 – 17. (Здобувач провела експериментальну частину роботи, пов'язану із вивченням орієнтовно-дослідницької діяльності і УРПУ за умов експериментальної гіпертироксинемії; провела аналіз отриманих даних; підготувала статтю до друку).
5. Гузь Л.В., Родинський О.Г., Гузь В.А. Характеристика збудливості еферентної ланки спинномозкових дуг за умов експериментального гіпертиреозу // Вісник морфології. – 2008. – Т. 14 (1). – С. 171 – 174.
6. Гузь Л.В., Родинський О.Г. Центральні та периферичні механізми вегетативної нервової системи // Збірка та доповіді міжнародної конференції, присвяченої пам¢яті ак. О.Г. Баклаваджяна. “Архив клинической и экспериментальной медицины”. Донецьк, 3 – 6 червня 2003 р. – Донецк, 2003. – Т. 12, № 1. – С. 33 – 34. (Здобувач провела експериментальну частину роботи, пов'язану із визначенням функціонального стану серцевої системи, провела статистичний аналіз, підготувала тези до друку).
7. Неруш П.О., Демченко О.М., Гузь Л.В., Романенко Л.А. Метаболизм фосфолипидов в мозгу и мнестические функции при тиреопатических состояниях: возрастной аспект // Тезисы докладов: “Нейронауки: теоретичні та кліничні аспекти”. Донецьк, 7-10 червня 2005 р. – С. 82 – 83. (Здобувач провела експериментальну частину роботи, пов'язану із вивченням орієнтовно-дослідницької діяльності і УРПУ; провела статистичний аналіз даних).
8. Неруш П.О., Демченко О.М., Гузь Л.В., Романенко Л.А. Особливості формування адаптивних реакцій організму за умов дисфункції щитоподібної залози // Тези доповідей II Міжнародної наукової конференції “Гомеостаз: фізіологія, патологія, фармакологія і клініка”. Одеса, 28 – 29 вересня 2005 р. – Одеса, 2005. – С. 232 – 235. (Здобувач провела експериментальну частину роботи, пов'язану із вивченням орієнтовно-дослідницької діяльності і УРПУ; провела статистичний аналіз даних).
9. Неруш П.О., Демченко О.М., Гузь Л.В. Стан вищої нервової діяльності та обмін ліпідів у головному мозку щурів за умов гіперфункції щитоподібної залози // Тези доповідей Всеукраїнської науково-практичної конференції “Проблеми вікової фізіології”. Луцьк, 29 – 30 вересня 2005 р. – Луцьк, 2005. – С. 86 – 87. (Здобувач провела експериментальну частину роботи, пов'язану із вивченням орієнтовно-дослідницької діяльності і УРПУ; провела статистичний аналіз даних).
10. Неруш П.О., Демченко О.М., Гузь Л.В., Романенко Л.А. Адаптивні механізми центральної нервової системи за умов дисфункції щитоподібної залози // Тези доповідей XVII з¢їзда Українського Фізіологічного товариства з міжнародною участю (пам¢яті ак. О.О. Богомольця). Чернівці, 18 – 20 травня 2006 р. – Чернівці, 2006. – С. 50. (Здобувач відтворила модель гіпертиреоїдного стану; провела експериментальну частину роботи, пов'язану із вивченням орієнтовно-дослідницької діяльності і УРПУ; провела статистичний аналіз даних).
11. Неруш П.О., Демченко О.М., Гузь Л.В. Нейрохімічна організація властивостей вищої нервової діяльності у щурів за умов дисфункції щитоподібної залози в процессі старіння (онтогенезу) // Тези доповідей Всеукраїнського Наукового симпозіуму до 100-річчя з дня народження В.О. Трошихіна, Черкаси, 13-15 листопада 2006р. - Черкаси, 2006. – С.76 – 77. (Здобувач провела експериментальну частину роботи, пов'язану із моделюванням гіпертиреоїдного стану та вивчення поведінкових реакцій та когнітивних розладів у щурів; провела аналіз статистичних даних).
12. Родинський О.Г., Гузь Л.В., Гузь В.А. Механизмы действия блокатора потенциал-управляемых калиевых каналов 4-аминопиридина на вызванные стимуляцией хронически денервированного седалищного нерва моносинаптические разряды, отводимые от вентрального корешка спинного мозга белых крыс // Тезисы докладов III съезда фармакологов России «Фармакология – практическому здравоохранению». Санкт-Петербург, 23 – 27 сентября 2007 г. – Санкт-Петербург, 2007. – Т. 2. – С. 1919. (Здобувач провела експериментальну частину роботи, пов'язану із вивченням основних характеристик збудливості сегментарних моносинаптичних рефлекторних систем у білих щурів; провела огляд літератури).
АНОТАЦІЯ
Гузь Л.В. Нейрометаболічні особливості та інтегративна діяльність центральної нервової системи за умов експериментальної гіпертироксинемії у щурів. – Рукопис.
Дисертація на здобуття наукового ступеня кандидата біологічних наук за спеціальністю 03.00.13 – фізіологія людини і тварин. – Харківський національний університет імені В.Н. Каразіна, Харків, 2008.
Тиреоїдний дисбаланс характеризувався розвитком поведінкового дефіциту і порушенням мнестичних функцій. Даний ефект супроводжувався накопиченням продуктів перекисного окислення ліпідів (ПОЛ) в різних структурах головного мозку у білих щурів. Зроблено висновок, що гіпертироксинемія супроводжується активацією ПОЛ, що проявляється в накопиченні первинних і вторинних продуктів ПОЛ. Порушення інтегративної діяльності центральної нервової системи супроводжувалось зміною спектра жирних кислот (кора великих півкуль, гіпокамп, стовбур мозку) і зниженням захисних властивостей глутатіонпероксидази (гіпокамп). Вивчали дію підвищеної концентрації тироксину на характер моносинаптичних рефлекторних реакцій спинного мозку білих щурів.
Ключові слова: гіпертироксинемія, інтегративна діяльність центральної нервової системи, перекисне окислення ліпідів, антиоксидантна система, кора великих півкуль, гіпокамп, стовбур мозку, спинний мозок, моносинаптична рефлекторна реакція.
АННОТАЦИЯ
Гузь Л.В. Нейрометаболические особенности и интегративная деятельность центральной нервной системы в условиях экспериментальной гипертироксинемии у крыс. – Рукопись.
Диссертация на соискание ученой степени кандидата биологических наук по специальности 03.00.13 – физиология человека и животных. – Харьковский национальный университет имени В.Н. Каразина, Харьков, 2008.
В опытах на белых крысах линии Вистар путем введения с пищей L-тироксина (10 мкг/сутки) в течение двух недель с ежедневным увеличением на 10 мкг от предыдущей дозы формировали состояние гипертироксинемии с определением в конце эксперимента в сыворотке крови концентрации тироксина иммуноферментным методом.
Показано, что гипертироксинемия сопровождалась нарушением ориентировочно-исследовательской деятельности в тесте «открытое поле», а также условно-рефлекторной реакции пассивного избегания. Анализ механизмов расстройств интегративной деятельности показал, что состояние гипертироксинемии характеризовалось повышением возбудимости моносинаптических рефлекторных дуг. Это проявилось увеличением амплитуды моносинаптического рефлекторного ответа (МРО), уменьшением латентного периода и порога возникновения рефлекторной реакции, что является свидетельством пластичности синапсов.
Показано, что длительная тетанизация дорсального корешка, при его раздражении пачкой из 5000 стимулов и частотой 300 Гц, сопровождалась усилением амплитуды МРО в течение почти 5 мин. При гипертироксинемии, наоборот, отмечалась выраженная депрессия амплитуды МРО. Возникновение депрессии МРО можно объяснить длительной деполяризацией постсинаптической мембраны мотонейронов в случае гипертироксинемии, что в свою очередь существенно уменьшает возбудимость в моносинаптической рефлекторной дуге. Изучение порога и хронаксии эфферентных волокон седалищного нерва показало, что у интактных животных среднее значение порога возникновения потенциала действия в дистальном участке вентрального корешка L5 составляло 17,5±1,3 мкА. При гипертироксинемии порог существенно повышался, и в среднем колебался в пределах 161,0±5,7% (р<0,005). В этих условиях хронаксия эфферентных волокон седалищного нерва была существенно снижена.
Анализ содержания свободных жирных кислот показал, что их количество в филогенетически различных отделах головного мозга различалось. В коре головного мозга интактных крыс уровень насыщенных кислот (пентадекановой и пальмитиновой) был более высоким. В стволе мозга отмечено повышение содержания стеариновой кислоты (С18:0), а также ненасыщенных олеиновой (С18:1) и арахидоновой (С20:4). В стволе головного мозга интактных крыс показатели насыщенных (С14:0, С18:0), а также ненасыщенных (С18:1, С18:2, С20:4) кислот были выше по сравнению с гиппокампом и корой мозга. Повышение уровня Т4 сказывалось и на содержании жирных кислот. Гипертироксинемия сопровождалась снижением концентрации насыщенных жирных кислот в коре головного мозга, содержание ненасыщенных кислот в спектре С18:1 – С20:4 увеличивалось.
Нарушение интегративной деятельности ЦНС сопровождалось изменениями прооксидантной и антиоксидантной систем структур мозга. Отмечено, что гипертироксинемия вызывала накопление как первичных (диеновые конъюгаты), так и вторичных (малоновый диальдегид) продуктов ПОЛ. При снижении защитных свойств глутатионпероксидазы это вызывало дезинтеграцию мембранных систем, ответственных за энергообеспечение, транспорт и управление.
Таким образом, в диссертации приведено теоретическое обоснование и экспериментальное исследование механизмов интегративной деятельности ЦНС в условиях модельной гипертироксинемии.
Ключевые слова: гипертироксинемия, интегративная деятельность центральной нервной системы, перекисное окисление липидов, антиоксидантная система, кора больших полушарий, гиппокамп, ствол мозга, спинной мозг, моносинаптическая рефлекторная реакция.
SUMMARY
Guz L.V. The neurometabolic features and integrative activity ofcentral nervous system in conditions of experimental hyperthyroxinemia in rats. – Manuscript.
The dissertation on competition of the scientific degree of candidate of biological sciences behind the speciality 03.00.13 – Human and animals physiology. –V.N. Karazin Кharkiv National University, Kharkiv, 2008.
Present-day ideas about a role of thyroid hormones in integrative activity of the central nervous system, genomic and extragenomic effects, and influence on activity of receptor system and neurotransmitter mechanisms of brain are mentioned. The interaction of nervous and humoral factors in their influence on behavioral activity and processes are discussed.
Thyroid disbalance was characterized by development of behavioral dificite and distress of near expressed mnestic function. The effect was accompanied by accumulation of the products of lipid peroxidation (LPO). The conclusion was made, that activity the lipid peroxidation accompanies HT and expresses in the accumulation of the primary and secondary products of lipid peroxidation. Integrate activity of CNS distress was accompanied by the reduction of protective mechanisms of glutathione peroxidase in hippocampus structures. Hyperthyroxinemia was characterized by changes of a spectrum of fatty acids in structures of a brain (cerebral cortex, hippocampus and brain stem) which affected integrative of CNS.
Key words: hyperthyroxinemia, integrative activity of the central nervous system (CNS), products of lipid peroxidation (LP), antioxidant system (AOS), cerebral cortex, hippocampus, brain stem, spinal cord (SC), monosynaptic reflex reactions.
0 комментариев