РЕФЕРАТ
На тему
Реакции клеточного иммунитета
ВВЕДЕНИЕ
Термин клеточный иммунитет первоначально служил для обозначения местных реакций, осуществляемых лимфоцитами и фагоцитами без участия антител — эффекторов гуморального иммунитета. Теперь этот термин часто используют в более широком смысле, для описания такого противоинфекционного или противоопухолевого иммунного ответа, в котором антителам принадлежит не ведущая, а вспомогательная роль.
Однако полностью разделить клеточный иммунитет и гуморальный невозможно: в инициации образования антител участвуют клетки, а в некоторых реакциях клеточного иммунитета важную связующую функцию выполняют антитела. Более того, не существует, по-видимому, клеточного иммунитета без образования антител, которые способны различными путями модифицировать опосредованный клетками иммунный ответ. Так, комплексы антиген—антитело вызывают высвобождение хемотаксических фрагментов комплемента, усиленно привлекающих лейкоциты в очаг воспаления, и, кроме того, благодаря Fc-рецепторам антитела могут принимать участие в связывании антигенов с клетками и тем самым влиять на реакции клеточного иммунитета, в частности обеспечивать прикрепление фагоцитов и цитотоксических Т-клеток к клеткам-мишеням. Вообще, при скоординированном иммунном ответе происходит многосторонний обеими сигналами между различными типами вступающих в него лейкоцитов и тканевыми клетками.
Межклеточная сигнализация в иммунной системе осуществляется путем непосредственного контактного взаимодействия клеток, в котором, участвуют их поверхностные молекулы, или с помощью цитокинов, называемых «белками связи». Эти белки действуют как растворимые медиаторы межклеточных взаимодействий. Вместе с гормонами и нейромедиаторами они составляют основу языка химической сигнализации, путем которой в многоклеточном организме регулируется морфогенез, регенерация тканей и иммунный ответ. Наряду с сигналами, возникающими при взаимодействии клеток с антигеном или друг с другом, существует цитокиновая сигнальная сеть, регулирующая реакции врожденного и приобретенного иммунитета, в том числе воспаление, противовирусную защиту, клональную пролиферацию антигенспецифичных Т- и В-клеток и их функции.
ЦИТОКИНЫ И ИХ КЛЕТОЧНЫЕ РЕЦЕПТОРЫ
Цитокины — это небольшие белки, действующие аутокринно или паракринно. Образование и высвобождение этих высокоактивных молекул обычно происходят кратковременно и жестко регулируются. К настоящему времени у человека идентифицировано уже более ста различных цитокинов, и постоянно появляются сообщения об открытии новых. Цитокины воздействуют на клетку, связываясь со специфическими рецепторами на цитоплазматической мембране и вызывая этим каскадную реакцию, ведущую к индукции, усилению или подавлению активности ряда регулируемых ими генов.
Многие цитокины имеют по несколько названий. Это связано с тем, что они были независимо открыты в различных областях исследований — иммунологии, вирусологии, гематологии, клеточной биологии и онкологии. К цитокинам относятся интерлейкины, обозначаемые сейчас номерами от ИЛ-1 до ИЛ-18, интерфероны, колониестимулирующие факторы, факторы некроза опухолей, факторы роста и хемокины. Причина многих недоразумений в номенклатуре цитокинов состоит в том, что они, по крайней мере in vitro, проявляют многообразные активности; примером может служить ИЛ-6, эффекты которого очень разнообразны. Кроме того, в ряде случаев один и тот же цитокин был выделен независимо в нескольких лабораториях при использовании совершенно разных экспериментальных систем. Путаницу с названиями усугубляет еще и частичное совпадение активностей у ряда цитокинов, создающее впечатление некоторой избыточности их функций. Дополнительные трудности в изучении цитокинов возникают из-за того, что эти медиаторы редко образуются по отдельности и редко действуют поодиночке. Одним словом, для цитокинов характерен сложный сетевой характер функционирования, при котором продукция одного из них влияет на образование или проявление активности ряда других. In vivo отдельная клетка организма редко становится мишенью какого-либо одного цитокина. Гораздо чаще отдельные цитокины служат как бы буквами некоего алфавита, образующими целое цитокиновое «слово», и реакция клетки возникает в результате воздействия на ее поверхность именно такого «слова».
Наиболее важные функции цитокинов и их рецепторов в иммунном ответе будут рассмотрены ниже; вначале необходимо остановиться на основных аспектах молекулярной биологии этих белков.
Цитокины и их рецепторы подразделяются на ряд семейств
Между индивидуальными цитокинами или их группами существует лишь небольшое сходство на уровне ДНК и аминокислотной последовательности, но все же они распределяются по гомологии на несколько больших семейств. Из них наиболее значительны три семейства: первое состоит из не менее чем 15 б-интерферонов, второе из более чем 50 хемокинов и третье включает цитокины, которые связываются с рецепторами для ФНО. Гораздо легче, однако, сгруппировать цитокины не по функциям, а по характеру их трехмерной структуры, и такое подразделение четко отражает внутригрупповое сходство клеточных цитокиновых рецепторов. Наиболее крупное семейство — суперсемейство — цитокиновых рецепторов характеризуется наличием в составе молекул внеклеточных участков с гомологичной последовательностью длиной примерно 200 аминокислотных остатков. К этому суперсемейству относятся рецепторы к ИЛ-2, ИЛ-3, ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-7, ИЛ-9, ИЛ-12, гранулоцитарному колониестимулируюшему фактору и гранулоцитарно-макрофагальному колониестимулирующему фактору. В него же входят рецепторы для гуморальных факторов, действующих преимущественно вне иммунной системы, — гормона роста и пролактина. Второе по величине семейство объединяет рецепторы к интерферонам всех типов, а также рецепторы к ИЛ-Йб, ИЛ-Йв и макрофагальному колониестимулирующему фактору. Это семейство входит как составная часть в суперсемейство иммуноглобулин-подобных молекул.
Цитокиновые рецепторы третьего семейства связывают ФНОа и ЦЗПв, лимфотоксин и ряд родственных цитокинов, в том числе фактор роста нервов. К этому же рецепторному семейству относится молекула Fas, связывание которой с лигандом FasL служит сигналом клеточной гибели.
Большинство цитокиновых рецепторов — это мембранные гликопротеины 1 типа, состоящие из одного-единственного трансмембранного домена. Однако действительно функциональные рецепторы, как правило, состоят из двух или большего числа субъединиц, которые могут иметь одинаковую структуру даже у различных по специфичности рецепторных комплексов. Обычно рецептор содержит «частную» высокоспецифичную субъединицу, способную связывать определенный цитокин, и «общую» субъединицу, которая встречается в рецепторах для других цитокинов. Например, рецепторный комплекс для ИЛ-2 состоит из трех субъединиц. Субъединица КР-2Св встречается также в рецепторе для ИЛ-15, а ИЛ-2Ру — в рецепторах для ИЛ-4, ИЛ-7 и ИЛ-9. Подобным же образом ИЛ-ьСв содержится в качестве субъединицы в рецепторах для таких цитокинов, как LIF, онкостатин М и ИЛ-11. Сходная функциональная активность некоторых цитокинов отчасти объясняется, возможно, наличием одинаковых субъединиц в их клеточных рецепторах. Поэтому, видимо, ИЛ-6, ИЛ-11 и онкостатин М одинаково действуют на гепатоциты, мегакариоциты и остеокласты, а дублирующий эффект ИЛ-2 и ИЛ-4 в качестве факторов роста Т-клеток обусловлен, по всей вероятности, присутствием в рецепторах для того и другого цитокина идентичной ИЛ-2Ру-це-пи. В то же время благодаря дифференциальной экспрессии частных рецепторных субъединиц каждый цитокин обладает и уникальной активностью в отношении клеток определенного типа. Например, LIF может задерживать дифференцировку эмбриональных стволовых клеток, тогда как ИЛ-6 такой активностью не обладает, поскольку эти клетки не экспрессируют соответствующего рецептора.
Все хемокины связываются рецепторами отдельного класса, объединенными на основе их уникальной структуры под общим названием семь трансмембранных гликопротеинов. Некоторые из них настолько специфичны, что связывают только один определенный хемокин, тогда как другие обладают сродством к ряду хемокинов. Существует также один рецептор, который «без разбора» связывает многие хемокины и, вероятно, принимает участие в ликвидации образующегося в очаге воспаления избытка этих медиаторов. Хемокины связываются также с в-адренорецепторами. Это еще одно свидетельство перекрывания системы цитокинов и других сетевых сигнальных систем, образуемых растворимыми медиаторами.
Связывание цитокиновых рецепторов активирует механизм внутриклеточной передачи сигналов
Современные представления о биологической роли цитокинов основаны на данных структурного анализа их молекул и изучении механизмов внутриклеточной передачи вызываемых ими сигналов. Благодаря таким исследованиям сейчас можно уже довольно детально проследить эту цепь последовательных событий белок-белкового распознавания, от момента связывания цитокина с клеточной поверхностью до мобилизации различных факторов транскрипции в ядре клетки. Как известно, первая стадия цитокиновой сигнализации — это вызванная присоединением цитокина агрегация субъединиц рецептора. Цитоплазматические «хвосты» этих субъединиц, взаимодействуя между собой, запускают нисходящий каскад сигнализации. В самом простом случае одинаковые субъединицы рецепторной молекулы, связавшись с цитокином, образуют гомодимер, в другом случае «частная» субъединица после присоединения цитокина вызывает гетеро- или гомодимеризацию «общих» субъединиц, передающих сигнал внутрь клетки.
Все цитокиновые рецепторы, относящиеся к первому семейству, как и представители некоторых других рецепторных семейств, ассоциированы с молекулами, названными Янускиназами. Активация цитокиновых рецепторов вызывает активацию Jaks, в частности киназ, фосфорилирующих тирозин. Большая часть функции цитокиновых рецепторов осуществляются с обязательной активацией Jaks. Выполняя свою главную функцию, т. е. агрегируя субъединицы рецептора, цитокин одновременно вызывает агрегацию Jaks. Затем под действием этих Янус-киназ происходит сопряженное с присоединением цитокина фосфорилирование остатков тирозина в составе различных сигнальных белков, в том числе переносчиков сигнала и активаторов транскрипции. Димеры белков Stats перемещаются к ядру клетки и связываются непосредственно с ДНК.
Каждый цитокин индуцирует различные механизмы внутриклеточной передачи сигнала в зависимости от того, какую из активностей он проявляет — общую с другими цитокинами или специфическую, индивидуальную. Например, в Т-клетках каждый из трех интерлейкинов — ИЛ-2, ИЛ-4 и ИЛ-9, взаимодействуя с субъединицей ИЛ-2Ру, активирует Jakl и Jak3; в то же время ИЛ-10 вызывает активацию Jakl и Тук2, а ИЛ-12 таким же образом действует на Jak2 и Тук2. Дополнительная вариабельность ответа клеток на цитокины возникает на этапе фосфорилирования Stats, поскольку каждый цитокин активирует «свой» набор этих переносчиков сигнала и активаторов транскрипции. В ответе на цитокины, вероятно, и состоит главная функция.Stats. Другие пути сигнализации, в первую очередь с активацией Ras/MAP-киназы, ведут к пролиферации клеток под действием соответствующих цитокинов, однако некоторые цитокины могут активировать механизм сигнализации, ведущий к апоптотической гибели клетки. Функциональная гибкость сигнальных систем возрастает еще больше благодаря тому, что активировать Stats могут не только Jaks, но и киназы иного происхождения. Например, в случае цитокинов ФНОсс и ИЛ-Йв действует механизм внутриклеточной сигнализации с участием не Jaks и Stats, а МАР-киназ; в результате с ДНК связываются такие активаторы транскрипции, как АР-1, NFkB и NFIL-6. Как специфическое, так и плейотропное действие хемокинов в конечном итоге влияет на перемещение клетки, но это сложный эффект: вслед за присоединением хемокинов к рецепторам происходит передача сигнала на G-белки, затем мобилизация вторых, внутриклеточных посредников, реорганизация цитоскелета, образование ограниченных адгезивных контактов, прилипание и отлипание клеточной поверхности, вытяжение и сокращение псевдоподий — все эти этапы необходимы для направленной миграции. Хемокины, подобные RANTES и МЙС-1б, могут также активировать Stats, образуя шунт между путями сигнализации с участием G-белков и Stats. Итак, постепенно область исследований цитокинов превращается из запутанного клубка множественных активностей во все более понятную систему белков-регуляторов со своими рецепторами и четкими путями сетевой внутриклеточной сигнализации.
Дифференцировка Т-хелперов на субпопуляции составляет важный этап в определении эффекторных механизмов иммунного ответа
Как теперь установлено, существуют две субпопуляции Тх-клеток CD4+, различающихся по набору синтезируемых ими цитокинов, и от этого профиля зависит, какой из двух основных типов иммунного ответа будет реализован. У человека Txl-клетки, как правило, продуцируют ИФу, ЦЗПв и ИЛ-2 и участвуют в опосредованных клетками воспалительных реакциях. Некоторые из цитокинов, выделяемых Txl, обладают провоспалительной активностью, а также стимулируют цитотоксические клетки и Т-эффекторы гиперчувствительности замедленного типа. В противоположность Txl-клеткам клетки Тх2 синтезируют ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-9, ИЛ-10 и ИЛ-13 и усиливают образование антител, особенно класса IgE. В результате они стимулируют гиперпродукцию антител и аллергические реакции. Помимо всего прочего, цитокины, выделяемые Txl-клетками, подавляют активность Тх2-клеток, и наоборот. Таким образом, любой иммунный ответ развивается в направлении либо Txl-, либо Тх2-типа.
Недавно выявлены различия между Txl- и Тх2-клетками по маркерам поверхности. Для клеточной мембраны Txl характерно наличие LAG-3 — антигена, относящегося к суперсемейству иммуноглобулин-подобных молекул. Клетки Тх2 экспрессируют в гораздо большем количестве по сравнению с Txl-клетками маркер CD30, относящийся к семейству рецепторов для ФНО.
Определение типа ответа — Txl или Тх2 — имеет принципиальное значение для развития эффективного иммунитета и, по-видимому, зависит от многих взаимодействующих между собой факторов. Среди них на характер дифференцировки Тх-клеток CD4+ и тем самым на профиль цитокинов, определяющих тип ответа, могут влиять следующие:
• профиль и количественное соотношение цитокинов, выделяемых под действием антигена. Например, ИЛ-12 служит мощным стимулом продукции Т- и НК-клетками г-интерферона и, следовательно, дифференцировки Тх-клеток в Txl. В то же время б-интерферон, синтезируемый в ранней фазе вирусной инфекции, способен не только индуцировать повышенную экспрессию ИЛ-12, но и переключать Тх-клетки с синтеза Тх2-набора цитокинов на Txl. Напротив, ранняя продукция ИЛ-4 способствует появлению клеток Тх2;
• доза антигена;
• антигенпрезентирующие клетки и выделяемые ими цитокины;
• генотип организма-хозяина и
• активность костимулирующих молекул и гормонов в месте взаимодействия антигена с Тх-клетками.
Из перечисленных факторов дифференцировки Тх-клеток особый интерес представляет последний. Например, глюкокортикоидные гормоны, концентрация которых нарастает в условиях физического или психологического стресса, могут направить развитие иммунного ответа по Тх2-типу. Этому эффекту противодействуют производные дегидроэпиандростерона, способствующие иммунному ответу Тх1-типа. Концентрации обоих этих гормонов надпочечника регулируются на системном уровне, а также зависят от местного метаболизма в тех органах, где происходит встреча антигена с Тх-клетками. Поэтому соотношение концентраций кортизола и ДГЭА в лимфоидной ткани или в очаге патологии может служить фактором, определяющим тип иммунного ответа.
Иммунный ответ не всегда строго поляризован по Txl- или Тх2-типу, так как, возможно, имеются и другие субпопуляции Т-хелперов. Тем не менее роли этих двух типов ответа как в формировании протективного иммунитета, так и в иммунопатологии приниипиально различны. К настоящему времени установлено, что для многих заболеваний характерно функционирование определенной субпопуляции Т-хелперов. Так, органоспецифическая иммунопатология, острое отторжение аллотрансплантата, рецидивирующие спонтанные аборты и рассеянный склероз связаны с иммунным ответом Txl-типа. При этих заболеваниях из крови больных легко удается выделить и клонировать Txl-клетки. Напротив, все клоны Т-клеток от больных атопической бронхиальной астмой, сенсибилизированных к пыльце растений, синтезируют, как и клоны больных системной красной волчанкой, цитокиныТх2-про-филя.
Т-клетки CD8+ также дифференцированы на субпопуляции с различными профилями выделяемых цитокинов
Многие цитотоксические Т-клетки CD8+ выделяют тот же набор цитокинов, что и клетки Txl. Существуют также Т-клетки CD8+, выделяющие цитокины Тх2-типа. Эта субпопуляция выполняет регуляторные и супрессорные функции. На дифференцировку Т-клеток CD8+ может влиять цитокиновый профиль Т-клеток CD4+. Например ИФуи ИЛ-12 способствуютдифференцировке Т-клеток CD8+ в субпопуляцию Тц1-клеток, а ИЛ-4 — в субпопуляцию Тц2. Однако клетки обеих этих субпопуляций обладают цитотоксической активностью и поражают свои мишени главным образом путем высвобождения содержимого гранул.
ЗАЩИТНЫЕ МЕХАНИЗМЫ, НЕЗАВИСИМЫЕ ОТ Т-КЛЕТОК
Фагоцитоз - важный компонент антимикробной защиты
Первоначальная защитная реакция на любую инфекцию в значительной степени зависит от распознавания общих для разных микробов компонентов особыми клеточными рецепторами, которые отличаются от антигенспецифичных рецепторов Т- и В-клеток.
Многочисленные компоненты микробных клеток способны вызывать хемотаксис фагоцитов в очаг инфекции. Некоторые из этих веществ, например бактериальный эндотоксин, привлекают фагоциты, индуцируя активацию комплемента по альтернативному пути с высвобождением С5а и СЗа. Другие обладают собственной прямой хемотаксической активностью. Так, присущие всем бактериям формилпептиды вызывают хемотаксис и, кроме того, непосредственно стимулируют фагоциты, всегда имеющие к ним рецепторы.
Начальная стадия фагоцитоза — это связывание микроба на поверхности фагоцитарной клетки. Связыванию способствует активация комплемента и фиксация на поверхности микробной клетки СЗЬ, с которым затем взаимодействуют рецепторы CR3 фагоцитов. Аналогичным образом, если предварительно с микробной клеткой связываются антитела, в ее поглощении участвуют затем Fc-рецепторы фагоцитов, тем самым способствуя фагоцитозу.
Микроорганизмы, для которых характерна внутриклеточная локализация в организме-хозяине, обладают особыми возможностями связывания с поверхностью фагоцитов: несмотря на то что поглощение происходит обычным путем, последующей активации бактерицидных механизмов не происходит.
Выделение цитокинов могут вызывать компоненты микробных клеток
Другой независимый от Т-клеток и антител механизм противомикробной защиты, весьма важный в начальной стадии инфекции, — это выделение цитокинов и хемокинов из макрофагов и прочих клеток. По-видимому, все инвазивные микробы содержат или выделяют молекулы, способные вызывать такой эффект. Среди микробных активаторов высвобождения цитокинов наиболее сильное действие оказывает эндотоксин, или липополисахарид. Сложным образом ЛПС взаимодействует с мембраносвязанными рецепторами на поверхности лейкоцитов и, вероятно, эндотелиальных клеток, в результате чего происходит активация соответствующих эффекторных функций этих клеток. Подобным образом может распознаваться и действовать и ряд других консервативных микробных структур. Среди цитокинов, выделяемых макрофагами под действием микробных компонентов, особая роль принадлежит ФНОа и ИЛ-12. Высвобождаемые на ранней фазе иммунного ответа, эти и другие медиаторы выполняют три следующие фундаментальные функции:
• служат сигналами для эндотелиальных клеток, начинающих в результате их получения привлекать лейкоциты из кровотока;
• активируют фагоцитарные клетки в тканях, обеспечивая тем самым «врожденную резистентность» в тот период, когда еще только развивается Т-клеточный иммунитет;
• служат одним из сигналов, предопределяющих тип Т-клеточного иммунного ответа — Txl или Тх2.
Цитокины необходимы для привлечения лейкоцитов из кровотока
Вначале цитокины вызывают экспрессию на эндотелиальных клетках молекул адгезии, благодаря которой лейкоциты слегка прилипают к поверхности эндотелия и начинают катиться по нему в направлении кровотока. На следующей стадии происходит выделение тканевыми клетками хемокинов, которые связываются с эндотелиоцитами и активируют экспрессию ими интегринов, запуская тем самым механизм усиления лейкоцитарной адгезии. В результате лейкоциты прочно прилипают к эндотелию и прекращают движение. Последняя стадия привлечения лейкоцитов — это миграция их через эндотелий сосудов в ткань.
Существование перечисленных стадий привлечения лейкоцитов иллюстрируют два синдрома иммунодефицита человека. При недостаточности лейкоцитарной адгезии II типа лейкоциты лишены сиалированного антигена X группы крови Льюис, который служит лигандом для Е-селектина, и поэтому неспособны катиться по эндотелию. В отличие от этого при недостаточности лейкоцитарной адгезии I типа в составе молекул интегринов отсутствует в-цепь, поэтому нейтрофилы, катясь по эндотелию, не могут прочно прилипнуть к нему и мигрировать из просвета сосуда в ткань. Оба синдрома недостаточности лейкоцитарной адгезии сопровождаются рецидивирующими бактериальными инфекциями.
Т-ЗАВИСИМЫЙ КЛЕТОЧНЫЙ ИММУННЫЙ ОТВЕТ
Выделяемые на самых ранних стадиях инфекции цитокины могут служить критерием, по которому легко определить тип последующего иммунного ответа. Это важный аспект клинической иммунологии, и в настоящее время он интенсивно разрабатывается. Такие разработки требуют четкого представления о возможных типах опосредованного клетками иммунного ответа и механизмах избирательной активации каждого из них. Они и будут рассмотрены в дальнейшем изложении.
Тх-клетки различных субпопуляций, выделяя тот или иной набор цитокинов, по-разному влияют на многообразные виды клеточной кооперации. Активация Т-клеток при повторной встрече со специфическим антигеном может быть причиной гиперчувствительности замедленного типа с образованием гранулем или иммунопатологического повреждения тканей. Некоторые Т-клетки способны подавлять иммунный ответ и поэтому названы Т-супрессорами. Отдельные Тс выделяют регуляторный цитокин — трансформирующий фактор роста в, и, вполне возможно, служат истинными «супрессорными» Т-клетками; остальные же могут быть просто регуляторными клетками, которые не подавляют, а переключают иммунный ответ с наблюдаемой в опыте формы на другую, не регистрируемую экспериментатором.
Выбор эффекторных механизмов клеточного иммунитета определяют Т-хелперные клетки
Какие из эпитопов антигена станут мишенью иммунного ответа, зависит прежде всего от Т-хелперов, поскольку именно они взаимодействуют с антигенпрезентирующими клетками, несущими антигенные пептиды в ассоциации с молекулами МНС класса II. Однако, противодействуя инфекции, иммунная система должна сделать также и второй, возможно еще более важный выбор — «решить», какой эффекторный механизм иммунного ответа необходимо использовать, чтобы он был адекватен характеру инфекции. Наиболее легко определяются три следующих эффекторных механизма:
• цитотоксическое действие Т-клеток CD8+ или больших гранулярных лимфоцитов;
• активация макрофагов, регулируемая главным образом Txl-клетками,
• стимулируемый Тх2 синтез антител и их роль в эффектах тучных клеток или эозинофилов.
Адекватный выбор механизма очень важен, поскольку в случае его несоответствия вместо защиты может развиться повышенная чувствительность к возбудителю. Например, при экспериментальной гриппозной инфекции противовирусную защиту осуществляют цитотоксические Т-клетки, тогда как активация макрофагов повышает чувствительность. У мыши активация макрофагов создает иммунитет против Leishmania major, тогда как все клеточные реакции, которые не ведут к активации макрофагов, могут иметь вредное действие, несмотря на образование антител.
ЦИТОТОКСИЧНОСТЬ Т- И НК-КЛЕТОК
В защитных реакциях, направленных на устранение инфицированных вирусами клеток организма, действуют цитотоксические Т- и НК-клетки
Клеточная цитотоксичность — важный механизм защиты против внутриклеточно локализованных возбудителей, таких как вирусы, некоторые бактерии и простейшие. Цитотоксическую активность могут проявлять несколько типов клеток — цитотоксические Т-клетки, нормальные киллерные клетки и иногда клетки миелоидного ряда, причем механизмы распознавания мишеней у них различные.
• Цитотоксические Т-клетки распознают специфические антигены в ассоциации с молекулами МНС. Большая часть Тц-клеток несет маркер CD8+ и распознает антиген, презентируемый в ассоциации с молекулами МНС класса I, но примерно 10% МНС-рестриктированных цитотоксических Т-клеток относятся к субпопуляции CD4+ и распознают антиген в ассоциации с молекулами МНС класса II.
• Нормальные киллеры распознают клетки, у которых отсутствует экспрессия молекул МНС класса I. Кроме отрицательного, З К-клеткам свойственно и положительное распознавание своих мишеней с помощью рецепторов к различным лигандам. Например, благодаря Fc-рецепторам они способны связывать антитела, образовавшие иммунные комплексы с антигенами на поверхности клеток-мишеней, — так называемая антителозависимая клеточная цитотоксичность, или киллерная клеточная активность.
Главная функция Тц — это устранение клеток, зараженных вирусами. Почти все имеющие ядро клетки экспрессируют молекулы МНС класса I и в случае вирусной инфекции способны презентировать антиген возбудителя цитотоксическим Т-клеткам CD8+. Собственные молекулы клеток после частичного расщепления в протеазомах транспортируются к эндоплазматическому ретикулуму, где они образуют комплекс с молекулами МНС класса I, и затем на поверхность клетки. Таким путем каждая клетка представляет «образцы» своих молекул для «осмотра» и опознавания Тц-клеткам CD8+. Тем же способом происходит презентация антигенов из микробов, которые локализуются внутри клеток.
Иногда для стабилизации связи между Тц-клеткой и ее мишенью требуются дополнительные взаимодействия; они могут способствовать и цитолизу. Например, цитолиз клеток-мишеней связавшимися с ними Тц-клетками in vitro можно вызвать антителами к CD3 или CD2, маркерам поверхности Тц. Вероятно, аналогичным образом цитолитическую активность Тц-клеток «включает» связывание с этими молекулами физиологических лигандов.
Некоторые вирусы, в частности вирусы герпеса, пытаются избежать распознавания Тц-клетками, подавляя экспрессию молекул МНС класса I на поверхности инфицированных клеток; однако в этом случае вирус распознают НК-клетки. Следовательно, Тц- и НК-клетки можно рассматривать как два взаимодополняющих инструмента иммунитета против вирусной инфекции тканей.
Цитотоксическую активность НК-клеток подавляют молекулы МНС класса I
НК-клетки, происходящие в основном из больших гранулярных лимфоцитов, у человека составляют примерно 5% лимфоцитов периферической крови. Чаще всего они имеют фенотип CD3-CD16+CD56+CD94+ и гаметное расположение генов Т-клеточного рецептора. В первоначальных работах по определению специфичности цитотоксического действия НК было установлено, что резистентность к нему обусловлена определенными доминантными аллелями локуса HLA-C. Впоследствии оказалось, что НК-клетки действительно способны распознавать различные аллотипы молекул МНС, однако любые из этих молекул, в том числе аллотипы локусов HLA-A и HLA-B, могут подавлять цитолиз.
Недавно опубликовано новое, весьма многообещающее открытие: обнаружены молекулы HLA-G, экспрессируемые только на клетках трофобласта и представляющие собой эффективные ингибиторы НК-цитотоксичности, которые обеспечивают устойчивость ко всем типам НК-клеток. Клетки трофобласта контактируют, когда сформирована плацента, с циркулирующей кровью матери, в отношении которой они аллогенны, поскольку обладают отцовскими генами МНС. Однако в этих клетках экспрессия всех обычных антигенов МНС, за исключением HLA-G, регуляторно подавлена. Следовательно, молекулы HLA-G необходимы для защиты плаценты от действия материнских НК-клеток.
НК-клетки распознают антигены МНС класса I с помощью молекул двух видов. Молекулы одного из них относятся к лектинам С-типа и первоначально были идентифицированы у мыши и крысы, а затем у человека. Молекулы второго вида — это представители суперсемейства иммуноглобулин-подобных молекул, CD158a и CD158b, а также обладающий тремя IgSF-доменами белок р70. Рецепторные молекулы этого типа вначале были идентифицированы у человека, позже эквивалентные структуры удалось выявить на клетках грызунов.
На поверхности З К- и К-клеток имеется несколько разных рецепторов для идентификации мишеней
НК-клетки атакуют свои мишени при участии в качестве рецепторов молекул CD2, CD16 и CD69, а также лектин-подобных рецепторов, сходных с теми, которые подавляют цитотоксичность. Рецептором для Fc НК-клетки связываются с молекулами антител, присоединившихся к поверхности клеток-мишеней и тем самым опосредующих АЗКЦ. Обычно это интерпретируется как проявление киллерной клеточной активности, но помимо З К-клеток такую функцию могут выполнять и другие типы клеток с Fc-рецепторами, в частности Т-клетки. Экспрессирующие Fc-рецепторы клетки миелоидного ряда также проявляют К-клеточную активность, но, по всей вероятности, в отличие от НК-и Т-клеток, с использованием иных механизмов цитолиза.
Потенциальная мишень для К-клетки — это экспрессированные на клеточной поверхности вирусные антигены, молекулы МНС и отдельные эпитопы, свойственные опухолевым клеткам. Кроме того, моноциты и полиморфноядерные гранулоциты могут проявлять киллерную активность в отношении нагруженных антителами клеток опухолей. Такие клетки миелоидного ряда, как моноциты и эозинофилы, выполняют, несомненно, важную эффекторную роль в уничтожении нагруженных антителами шистосомул.
Активированные цитокинами клетки-киллеры относятся, вероятно, к НК-клеткам
В иммунологии разрабатывается несколько направлений экспериментальной иммунотерапии рака. Одно из них - это активация интерлейки-ном-2 in vitro собственных лимфоцитов больного с последующим их обратным введением. Такие лимфоциты, выделенные из крови или селезенки, получили название активированные цитокинами клетки-киллеры. Они проявляют не рестриктированную по МНС повышенную цитотоксичность и, по-видимому, происходят преимущественно из предшественников, не отличающихся от З К-клеток. Скорее всего, ЛАК — это продукт активации, а не какая-то особая клеточная линия. Способ противоопухолевой иммунотерапии с использованием ЛАК пока проходит этап клинических испытаний.
Цитотоксический эффект клеток-киллеров реализуется либо при контактном взаимодействии с мишенями, либо путем выделения цитокинов и экзоцитоза гранул
Для поражения мишени Тц-, З К- и К-клетки обладают несколькими механизмами воздействия. Один из них — это передача сигналов при непосредственном клеточном контакте через поверхностные структуры, другой — непрямая сигнализация с помощью цитокинов. Кроме того, в цитоплазме многих цитотоксических Т-клеток CD8+ и БГЛ обнаружены гранулы с белками, высвобождение которых вблизи цитоплазматической мембраны клетки-мишени вызывает ее повреждение. Какое из сочетаний этих трех механизмов цитотоксического действия будет использовано в конкретных условиях, зависит от типа цитотоксических клеток, в частности от субпопуляции Тц-клеток.
В гранулах цитотоксических Т-клеток содержатся перфорин и гранзимы
Гранулы НК-клеток и цитотоксических Т-клеток содержат несколько белков, в том числе перфорин и гранзимы. Как только цитотоксическая Т-клет-ка связывается со своей мишенью, гранулы внутри Тц перемещаются к тому участку мембраны, где произошел контакт с мишенью. Затем осуществляется Са2+-зависимое высвобождение содержимого из этих гранул в щель между цитотоксической клеткой и ее мишенью.
Перфорин — это мономерный белок, вызывающий образование пор в цитоплазматической мембране. По структуре и функции он близок к С9. Кроме перфорина, гранулы содержат сериновую эстеразу, которая может входить в состав литического комплекса. В присутствии Са2+ мономеры перфорина связываются с мембраной клетки-мишени и полимеризуются, образуя трансмембранный канал. Несмотря на тесный контакт собственной плазматической мембраны с перфорином, сама цитотоксическая Т-клетка не повреждается им и продолжает поражать следующие клетки-мишени. Возможно, от самоповреждения
Ф клетку защищает протеогликан хондроитин-сульфат А, который также присутствует в гранулах. Этот белок может связываться с перфорином и вызывать в результате его инактивацию.
Гранзимы — это набор сериновых эстераз, которые высвобождаются при экзоцитозе гранул и затем активируются. Действие гранзимов не обязательно для проявления цитотоксичности — клетки, лишенные этих ферментов, еще способны поражать свои мишени. Некоторые из гранзимов могут вызывать в клетках-мишенях запуск программы апоптоза и фрагментации ДНК, воздействуя на пути внутриклеточной сигнализации.
Клетки-мишени могут также воспринимать циютоксические сигналы через рецепторы для FasL и ФНО
Цитотоксические Т-клетки CD4+, как выяснилось, не содержат перфорин и поражают свои мишени в отсутствие Са2+. Таким образом, они обладают иным механизмом цитотоксического действия, что было доказано в опытах на Тц-клетках «нокаутных» мышей. У таких мышей в результате направленной, сайт-специфической мутации не экспрессируется ген перфорина. В отсутствие перфорина цитотоксическая активность Т-клеток CD4+ снижена, но не устранена полностью. Поиск дополнительного механизма цитотоксичности привел к идентификации на поверхности клеток-мишений группы молекул, связывание с которыми служит сигналом к апоптозу. Среди них прежде всего представляют интерес Fas и рецепторы для ФНО. В эту группу молекул входят также CD30 и CD40. Лиганд для Fas, т. е. FasL, экспрессируется на зрелых Т-клетках CD4+ и CD8+ после их активации. Связывание FasL с рецепторными молекулами Fas вызывает агрегацию последних и присоединение к их внутриклеточным сегментам белка MORT-1, что в итоге приводит к апоптозу клеток-мишеней. От Fas зависит также выживание и устранение определенных субпопуляций лимфоцитов в период их созревания. По своему строению молекулы Fas похожи на рецепторы для ФНО. Например, те и другие имеют внутриклеточные «смертоносные» домены — структурные мотивы, обнаруженные у ряда особых белков, контролирующих выживание клеток.
Гранулы Тц-клеток могут содержать также ФНОа и лимфотоксин. Однако сами по себе эти цитокины не могут обеспечить цитотоксичность: отдельная Тц-клетка способна вызвать гибель клетки-мишени за 3—4 ч, а действие ФНО проявляется гораздо позднее. Но сходство между ФНО-рецептором и Fas свидетельствует о потенциальном участии ФНОа и ЦЗПв в поражении мишеней.
Итак, цитотоксическое действие Г-клеток CD8+ основано на высвобождении содержимого гранул и экспрессии FasL, Т-клеток CD4+ — в основном на экспрессии FasL, а НК-клеток, в первую очередь, на высвобождении содержимого гранул. В цитотоксическую активность всех этих клеток может вносить вклад ФНО.
Цитотоксическое действие клеток миелоидного ряда основано на целом ряде механизмов поражения мишеней
Уничтожение опухолевых клеток макрофагами часто обусловлено действием только ФНОа. Вместе с ИФу, выделяемым Т- и НК-клетками. этот цитокин вызывает мощный синергичный эффект цитолиза чувствительных опухолей. Механизмы цитотоксического действия, обусловленные активностью цитокинов. пока недостаточно изучены. В ряде случаев цитокины вызывают повышение активности циклооксигеназы и липоксигеназы с последующим внутриклеточным образованием свободных радикалов. Кроме того, возможны такие цитокин-зависимые эффекты, как образование свободных радикалов в результате митохондриального транспорта электронов и нарушение белкового синтеза. Клетки миелоидного ряда способны также выделять те же токсические медиаторы, которые действуют при разрушении микробных клеток внутри фагоцитов, в частности высокоактивные нестабильные метаболиты кислорода и азота.
РОЛЬ МАКРОФАГОВ В ИММУННОМ ОТВЕТЕ
Макрофаги принимают участие в иммунном ответе на всех его этапах. Во-первых, как уже было отмечено, они осуществляют немедленную защитную реакцию до тех пор, пока не произойдет усиление иммунного ответа, регулируемое антигенспецифичнми Т-клетками. Во-вторых, они вызывают активацию Т-клеток, осуществляя процессинг и презентацию им антигена. И наконец, активированные в свою очередь Т-клетками, они выполняют важные функции в эффекторных механизмах клеточного иммунитета, вызывая воспаление и уничтожая микроорганизмы, а также опухолевые клетки.
Цитокины усиливают некоторые функции макрофагов
Циркулирующие моноциты способны уничтожать некоторые микроорганизмы. При культивировании in vitro они в значительной степени теряют эту активность, но под действием добавленных цитокинов, в частности ИФу, она восстанавливается и параллельно происходит активация дополнительных механизмов антимикробного действия, которые в норме не экспрессируются моноцитами.
Такая «активация» цитокинами необходима макрофагам in vitro для разрушения многих внутриклеточных паразитов и некоторых опухолевых клеток. Классический эксперимент, демонстрирующий этот феномен, был проведен на животных, иммунизированных БЦЖ. Введение им очищенных белков туберкулина, т. е. смеси антигенов Mycobacterium tuberculosis, стимулирующих Т-клетки, вызывает, помимо стимуляции противотуберкулезного иммунитета, резистентность и к другому патогенному микроорганизму — Listeria monocytogenes. При анализе этого эффекта выяснилось, что стимуляция макрофагов происходит по антигенспецифическому механизму, но приводит к усилению их неспецифической бактерицидной активности. Как показали дальнейшие исследования, лимфоциты мышей, иммунизированных БЦЖ, при культивировании in vitro в присутствии соответствующего антигена, выделяют в среду цитокины, усиливающие способность макрофагов сдерживать размножение или уничтожать как микобактерий, так и другие микробы.
Макрофаги весьма разнообразны по свойствам
Активность макрофагов — это сложный феномен. Активированные фагоцитарные клетки приобретают повышенную способность уничтожать одни микроорганизмы, не затрагивая другие. Например, очищенный ЙєЦг стимулирует бактерицидную активность моноцитов человека в отношении Legionella, но при этом усиливает рост Mycobacterium tuberculosis. Такой неоднозначный характер эффекта обусловлен несколькими причинами:
• множественностью эффекторных функций, выполняемых активированными макрофагами;
• большим разнообразием моноцитов и макрофагов по их свойствам; в зависимости от ткани и органа они различаются по экспрессии молекул МНС класса II и Fc-рецепторов, профилю выделяемых цитокинов и продукции пероксидазы. Тем не менее большинство исследователей считает, что все макрофаги принадлежат к одной клеточной линии, а наблюдаемые различия обусловлены последовательными стадиями их созревания и влиянием тканевого микроокружения; кроме того, активация тех или иных функций может зависеть не только от природы макрофагов, но и от конкретного «спектра» цитокинов и других провоспалительных стимулов. Предположительно активация макрофагов происходит в несколько этапов, под влиянием следующих один за другим стимулов, которыми могут служить цитокины, эндотоксин, различные медиаторы и регуляторные факторы воспаления. На каждом этапе активации макрофаги способны к осуществлению различных эффекторных функций и обладают характерными особенностями морфологии и физиологии.
В некоторых случаях для стимуляции определенной функциональной активности макрофагов требуется несколько сигналов. Например, чтобы вызвать наибольшую продукцию оксида азота NO, токсичного для бактерий и опухолевых клеток, макрофаги мыши необходимо стимулировать сначала ИФу, а затем ФНОа. На макрофагах человека данный эффект получить гораздо труднее. В большинстве случаев для этого требуется серия стимулов, например воздействие несколькими цитокинами с одновременной перекрестной сшивкой FceRII. Макрофаги человека, выделенные из воспалительного очага, иногда экспрессируют индуцибельную синтазу оксида азота, но необходимый для его синтеза кофактор тетрагидробиоптерин они содержат в низкой концентрации. Поскольку оксид азота выполняет многочисленные сигнальные функции, не связанные с его токсическим действием, можно предполагать, что токсикантом служит не само это соединение азота, а преимущественно пероксинитриты, образующиеся в результате взаимодействия N0 с продуктами восстановления кислорода. Обычно такое взаимодействие происходит только в очагах воспаления и при стимуляции фагоцитарной активности макрофагов.
У человека в активации макрофагов и регуляции баланса Тх1/Тх2 принимает участие кальцитриол
Под действием ИФу макрофаги человека экспрессируют Й-б-гидроксилазу, которая может превращать неактивный циркулирующий 25-гидроксихолекальциферол в активный 1,25-ди-гидроксихолекальциферол. На поверхности макрофагов имеются рецепторы для кальцитриола, и он дополнительно стимулирует эти клетки. Кроме того, по механизму обратной отрицательной регуляции кальцитриол оказывает мощный подавляющий эффект на Txl-лимфоциты. Этот эффект, по всей вероятности, служит одним из путей переключения иммунного ответа с Txl- на Тх2-тип в тех случаях, когда возбудитель не может быть устранен из тканей и воспаление как реакция клеточного иммунитета становится хроническим. Данный механизм имеет особое значение у человека, поскольку при саркоидозе и туберкулезе образование кальцитриола может быть настолько значительным, что из участка активации макрофагов он проникает в кровоток, вызывая повышение содержания кальция в крови.
Помимо положительной существует и отрицательная регуляция эффекторных функций макрофагов
Как установлено, макрофаги могут быть не только активированы, но и дезактивированы. Подавление их функций способны вызывать простагландин Е и отчасти глюкокортикоиды. Недавно из среды, в которой культивировались опухолевые клетки, был выделен и получен в очищенном виде фактор, дезактивирующий макрофаги, который способен отменить вызванное МЦгувеличение образования высокоактивных метаболитов кислорода и в некоторой степени NO. Таким же эффектом обладают ИЛ-4 и пептид, связанный с геном кальцитонина, а также семейство ФЦСв-подобных цитокинов.
ОБРАЗОВАНИЕ ГРАНУЛЕМ
Иногда клеточный иммунитет не обеспечивает устранения проникших в ткани микробов либо антигенный материал не элиминируется из-за того, что устойчив к ферментативному расщеплению или просто относится к собственным компонентам организма. Если при этом Т-клетки продолжают накапливаться и выделять цитокины, образуется гранулема. Появление в тканях гранулем характерно для инфекций, возбудители которых локализуются, хотя бы отчасти, внутриклеточно, либо по размерам крупнее макрофагов, либо проявляют тенденцию персистировать в тканях.
Как правило, гранулемы содержат клетки — производные макрофагов, в том числе эпителиоидные и гигантские многоядерные клетки; функции этих клеток еще неполностью известны. По морфологии эти компоненты гранулем скорее относятся к секреторным, а не к фагоцитарным клеткам, и появляются, по-видимому, в результате хронической стимуляции макрофагов цитокинами.
Входящие в состав гранулем Тх-клетки CD4+ располагаются в центре этих образований, а Т-клетки CD8+ на их периферии. Это позволяет предполагать, что Т-клетки CD4+ выполняют решающую роль в привлечении и активации других лимфоцитов и макрофагов. При культивировании гранулематозной ткани in vitro обнаружено выделение ею в среду различных цитокинов. Для максимального развития гранулем, по-видимому, требуются выделяемые Txl-клетками цитокины и ФНОа; в случае мышиного шистосомоза для этого необходимы также цитокины. выделяемые Тх2-клетками.
ИММУНОПАТОЛОГИЯ
Повреждения тканей, наблюдаемые при инфекционных заболеваниях, иногда частично или полностью обусловлены действием самих механизмов клеточного иммунитета. К повреждению тканей может привести также иммунный ответ на аутоантигены.
Цитотоксичность
Цитотоксические клетки могут поражать инфицированные вирусами клетки-мишени собственного организма, весьма важные для его жизнедеятельности, например клетки центральной нервной системы. Если это происходит при иммунном ответе на вирусную инфекцию, которая сама по себе не вызывает гибели или функциональных нарушений зараженных клеток, повреждение тканей относят к иммунопатологии.
Хроническое воспаление
Реакции клеточного иммунитета могут быть направлены против аутоантигенов и в этом случае вызывают хроническое воспаление, разрушающее ткани. Часто роль в этом предполагаемых инфекционных агентов и постинфекционных аутоиммунных реакций остается неясной, как, например, в случае разрушения островков Лангерганса поджелудочной железы и возникновения в результате инсулин-зависимого диабета.
Вытеснение гранулемами функциональной ткани
Занимая в пространстве определенный объем, гранулема может нарушать функцию той или иной ткани организма. Например, образование гранулемы, вызванное присутствием микобактерий — возбудителей проказы, может приводить к повреждению нервов, по ходу которых эти гранулемы возникают. Аналогичным образом функциональные нарушения могут вызывать гранулемы, образовавшиеся в сетчатке глаза или в тканях мозга.
Избыточное выделение цитокмнов
Ряд патологических синдромов, таких как токсический шок геморрагический некроз и реакция Шварцмана, а также локальный некроз при реакциях гиперчувствительности замедленного типа, обусловлен, по-видимому, избыточным выделением цитокинов.
СЕТЕВЫЕ ВЗАИМОДЕЙСТВИЯ ЦИТОКИНОВ
Цитокины вступают в сложные сетевые взаимодействия
Как можно разобраться в сложной сети взаимодействий одних цитокинов с другими и понять роль каждого из них в организме? Наиболее эффективным инструментом для этого служит применяемый на мышах молекулярно-генетический метод «генного нокаута» — разрушения гена того или иного цитокина либо его рецептора. Определенные цитокины. например ИЛ-7. ФНОа и ЦЗПв. существенно важны для формирования иммунной системы в период внутриутробного развития Последствия устранения других цитокинов, в частности ИЛ-2, ИЛ-4, ИЛ-12 и ИФу, проявляются ярче всего, когда животное испытывает вызванный инфекцией «стресс». На мышах, лишенных гена ИЛ-12, было установлено значение этого цитокина для продукции ИФу и развития реакций гиперчувствительности замедленного типа. Разрушение некоторых цитокиновых генов вызывает у мышей поражения, напоминающие воспалительные заболевания слизистой оболочки кишечника у человека, еще раз подтверждая тот факт, что она представляет собой ту ткань организма, которая с наибольшим постоянством испытывает антигенные воздействия. Доказательством противовоспалительной роли ФЦСв служит то, что у лишенных его мышей образуется множество очагов воспаления. Выраженная иммунологическая недостаточность у мышей, не экспрессирующих г-цепь рецептора к ИЛ-2, свидетельствует, что эта цепь служит обшей сигнальной субъединицей в составе ряда цитокиновых рецепторов.
Все эти факты следует интерпретировать с известной осторожностью. Так, нельзя правильно оценить роль конкретного цитокина в организме зрелой особи, если его отсутствие в период развития иммунной системы привело к иммунологической недостаточности. Однако представляется вполне вероятным, что каждый цитокин при всем многообразии своих функций имеет среди них одну или несколько специфических, в связи с которой он сохранился в процессе эволюции.
На мышах, лишенных соответствующих генов, можно также выяснить функциональное значение ферментов Jak и Stat. Например, дефицитные по Stat-1 мыши чрезвычайно восприимчивы к вирусным инфекциям; у них нарушено образование ИФа и ИФу, а также оксида азота N0 в макрофагах, и противоинфекционной иммунный ответ развивается по Тх2-типу. Мыши, гомозиготные по делеции гена Jak-1, в отличие от Stat-дефицитных, погибают сразу после рождения. Они не способны питаться, у них нарушен ответ клеток на ИЛ-7 и не образуются лимфоциты всех популяций. Фенотнпически это состояние напоминает тяжелый комбинированный иммунодефициту мыши и человека.
Регуляция сетевых цитокиновых взаимодействий in vivo имеет ряд особенностей. • Образование индивидуальных цитокинов происходит кратковременно и находится под жестким контролем.
• Разные цитокины действуют как синергисты или антагонисты.
• Каждый из цитокинов может индуцировать или ингибировать продукцию других цитокинов.
• Каждый из цитокинов способен регулировать экспрессию клеточных рецепторов для самого себя и для других цитокинов.
• Антагонисты цитокинов связываются со специфическими клеточными рецепторами, но не вызывают передачу внутриклеточного сигнала.
• «Децепторы» специфически связывают лиганд, но не передают сигнал внутрь клетки.
• Благодаря ферментативному отщеплению внеклеточные домены цитокиновых рецепторов могут покидать поверхность клетки и связываться со свободными молекулами многих цитокинов; их можно обнаружить в сыворотке и моче.
Интенсивность синтеза цитокинов до и после его стимуляции может быть также запрограммирована на генетическом уровне. Например, носители определенных аллелей генов ФНОа и ИЛ-Йв часто страдают некоторыми хроническими воспалительными заболеваниями, что, возможно, обусловлено наследственными особенностями продукции цитокинов.
Сетевыми взаимодействиями цитокинов обусловлено множество физиологических эффектов
В путях эволюции и функционировании иммунной и нервной систем можно найти много общего. Например, обе эти системы за счет межклеточной коммуникации обладают способностью к обучению и хранению памяти; кроме того, среди их компонентов много общих медиаторов, рецепторов и антигенов. Для функционирования обеих систем требуется внутренняя коммуникационная сеть, а также сеть, которая обеспечивает регуляцию и взаимодействие с другими органами. Посылая сигналы на периферию, центральная нервная система непосредственно «телеграфирует» по нервным стволам и волокнам почти во все органы и ткани, а также использует ось гипоталамус—гипофиз—надпочечники. В противоположность этому иммунная система, состоящая главным образом из подвижных, свободных клеток, осуществляет внутрисистемные связи на основе взаимодействия этих клеток между собой. В то же время с другими органами и тканями она связана прежде всего посредством цитокинов, но следует подчеркнуть, что эти ее регуляторные воздействия всегда локальны, а не системны. Интеграция иммунной системы с нервной и эндокринной увеличивает ее влияние на организм. Например, такие цитокины, как ИЛ-1, ФНОа и ИЛ-6, непосредственно воздействуют на гипоталамус или гипофиз. Наряду с этим различные клетки иммунной системы экспрессируют рецепторы для нейромедиаторов, опиатов и нейропептидов.
Похожие работы
... эмбрионах размножаются и многочисленные бактерии. В последнее время накопились экспериментальные материалы, указывающие на наличие в эмбриональной жизни особой иммунологической реактивности. Патология иммунитета. Долгое время считалось установленным, что организм не отвечает образованием антител на собственные антигены. Эрлих считал это проявлением своеобразного «страха ...
... на его течение профессиональные вредности и стрессовые ситуации. Все это необходимо учитывать при диагностике и лечении атопического дерматита у пациентов разных возрастных группК л и н и ч е с к и е ф о р м ы а т о п и ч е с к о г о д е р м а т и т а. Единой классификации форм атопического дерматита в литературе нет. Многие авторы, в зависимости от возраста больных выделяют несколько форм ...

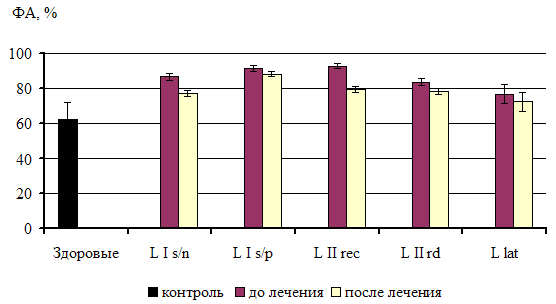
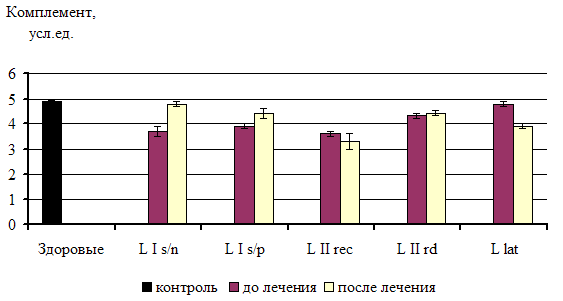
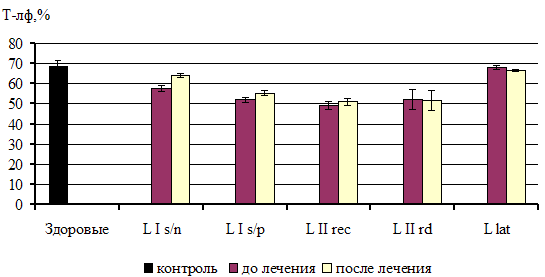
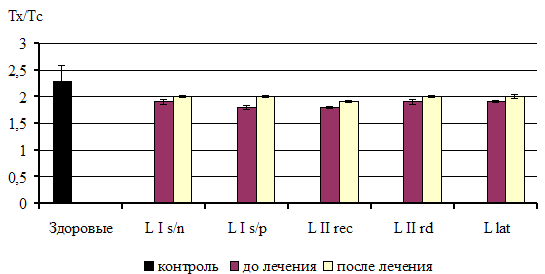
... из исследования. 3.2. Факторы неспецифической резистентности у больных сифилисом до и после специфической терапии При иммунологическом обследовании у всех больных ранними формами приобретенного сифилиса было выявлено достоверное повышение фагоцитарной активности нейтрофилов по сравнению с показателями группы контроля. Таблица 1 Показатели неспецифической резистентности у больных ...
... фиброза и деформации лёгочной ткани. Фиброзно-кавернозный туберкулёз значительно хуже подвергается заживлению, чем кавернозный. Цирротический туберкулёз. Цирротический туберкулез лёгких характеризуется развитием в лёгочной ткани грубого, деформирующего орган склероза (цирроза), бронхоэктатических, посткавернозных типа кист полостей, эмзифематонных булл или каверн без признаков прогрессирования. ...
0 комментариев