ФИЗИОЛОГИЯ РЕФЛЕКСОВ
Содержание
Рефлекс
Синаптическая передача
Односторонняя передача и синаптическая задержка
Понятие о рефлекторном центре
Облегчение и торможение
Центральная суммация и интенсивность центрального раздражения
Эффект отдачи и разряд последействия
Фракционирование
Окклюзия
Количественная рефлекторная регуляция эфферентных нейронов
Реципрокная иннервация
Заключение
Рефлекс
Согласно наиболее краткому определению, рефлекс представляет собой реакцию эффекторов на раздражение рецепторов. Центральная нервная система обеспечивает необходимую связь между афферентной и эфферентной частью рефлекторной дуги. Большинство видов нервной регуляции эффекторов осуществляется через такие дуги, которые широко варьируют по сложности. Простейшие рефлексы, такие, как коленный, мигательный, отдергивание конечности от болевого раздражителя и другие, осуществляются очень быстро через относительно простые дуги; они строго фиксированы или детерминированы раздражителем. Более сложные формы нервной интеграции в основном имеют ту же природу. Однако межнейронные связи центральной нервной системы широко варьируют по сложности, и благодаря взаимодействию между различными центральными структурами конечный эффект не отличается постоянством. Классификация отдельных видов нервной интеграции иногда представляет большие трудности: не легко разграничить рефлекторный и произвольный акты.
Многие аспекты поведения нельзя связать с определенными раздражителями, и тогда мы называем их произвольными актами. Однако раздражитель всегда имеется. Можно свести внешние раздражения к минимуму, но полностью устранить их нельзя; то же относится и к раздражениям, исходящим из самого организма. Трудно оценить общее влияние, т. е. суммарный эффект множества раздражений, которые беспрерывно посылают сигналы в центральную нервную систему. Многие на первый взгляд спонтанные акты на самом деле могут быть следствием такой суммации. Поскольку центральная нервная система получает самую разнообразную информацию (сигналы от многих видов различных рецепторов), трудно правильно предсказать характер реакции на определенный раздражитель. Кроме генетически детерминированных биохимических и морфологических форм организации нервного субстрата и состояния омывающих его жидких сред, программирование функций организма определяется общим притоком сигналов к центральной нервной системе, притоком, который включает как прошлые, так и действующие в настоящее время сенсорные сигналы. В нервной системе под влиянием получаемых ею импульсов происходят стойкие сдвиги неизвестного характера, так что регуляция функций организма лишь отчасти определяется сигналами, поступающими в данный момент. Другими словами, программа, вырабатываемая нервной системой в ответ на получаемую текущую информацию, частично определяется прошлым опытом.
Рефлекс представляет простейшую форму интегративной деятельности центральной нервной системы, и каждый сегмент располагает всеми необходимыми механизмами для его осуществления. К таким механизмам относятся рецепторы, афферентные волокна, идущие к спинному мозгу, эфферентные волокна к эффекторному органу, сам эффектор и важнейшие синаптические связи между афферентными и эфферентными нейронами в центральной нервной системе. Все эти элементы вместе составляют простую рефлекторную дугу. Такая рефлекторная дуга представляет собой моносинаптическую, или двухнейронную, организацию, поскольку афферентный нейрон образует синапс непосредственно с эфферентным. В большинстве дуг между афферентным и моторным нейронами имеются вставочные нейроны, так что рефлексы мышц на растяжение в сущности являются единственными, имеющими моносинаптическую дугу. В сегментарной системе заложены механизмы для таких рефлексов, как коленный, сгибательный, перекрестный разгибательный и многие другие. Большинство сегментарных рефлексов «а самом деле обеспечивается не одним сегментом, а несколькими связанными. Однако реакции осуществляются через наикратчайшие пути и взаимодействие между сегментами не обязательно.
Некоторые из рефлексов, осуществляемых через черепно-мозговые нервы, принципиально сходны со спинномозговыми сегментарными рефлексами, хотя для этих нервов не характерно правильное размещение, типичное для спинномозговых нервов,— слияние сегментов и неравномерный характер роста ствола мозга приводит к утрате явной сегментарности. Примером такого рефлекса может служить зрачковый рефлекс на свет, осуществляемый через II и III пару черепномозговых нервов в качестве афферентного и эфферентного путей (соответственно).
Собственные волокна спинного мозга обеспечивают функциональную связь между центрами на различных уровнях и участвуют в межсегментарных рефлексах. Координация движений передних и задних конечностей, шеи и туловища, координация мышц, осуществляющих движения в различных суставах конечности, обеспечиваются посредством таких связей. Хотя рефлексы часто рассматриваются так, как если бы их дуги были изолированы друг от друга, на самом деле между ними существует тесная связь. Облегчающие и тормозящие взаимные влияния между ними частично обусловливают описаннные реципрокные эффекты, а также доминирование некоторых рефлексов.
Рефлекторные акты, осуществляемые без изменения центральных нервных процессов прошлым опытом организма, называются врожденными, или безусловными, рефлексами. В основе таких рефлексов лежит структурная связь специфических рецепторов с определенными эффекторами. Связь эта осуществляется определенными нервными путями (рефлекторные дуги). Примером может служить выделение слюны в ответ на возбуждение вкусовых рецепторов. Коленный рефлекс, мигание, рефлекторное отдергивание конечности, опорожнение мочевого пузыря или кишечника, глотание, кашель и многие другие рефлексы относятся к той же категории. Важная особенность этих актов — их целесообразность для организма.
Условные рефлексы в отличие от врожденных основаны на прошлом опыте, т. е. связаны с тем или иным видом обучения. Все рецепторы анатомически связаны более или менее прямыми путями через центральную систему со всеми эффекторами. Вследствие избирательности, возникающей в результате особенностей синаптического проведения, многие потенциально функционирующие связи между рецептором и эффектором либо не получают адекватного возбуждающего влияния для функционального замыкания «цепи», либо подвергаются активному торможению со стороны других, параллельно протекающих процессов. Одним из наиболее известных примеров является павловский условный слюноотделительный рефлекс. Звон колокольчика не вызывает слюноотделения, если предварительно его не сочетают несколько раз с дачей пищи, которая химически раздражает вкусовые рецепторы, вызывая безусловный слюноотделительный рефлекс. После того как в центральной нервной системе возникнут изменения под влиянием комбинированного воздействия импульсов, вызываемых звуковым и вкусовым раздражителями, звук колокольчика становится столь же эффективным стимулятором секреции слюны, как и прием пищи. В обоих случаях эфферентный путь к слюнным железам один и тот же, но, поскольку афферентные импульсы поступают в центральную нервную систему из совершенно разных источников, центральные пути, несомненно, различны.
Синаптическая передача
Для синаптической передачи необходимо осуществление довольно жестких требований. Этот факт имеет первостепенное лимитирующее значение в распространении импульсов в центральной нервной системе. Для того чтобы импульсы пресинаптических нейронов вызывали эффективное раздражение постсинаптических нейронов, должны быть выполнены следующие условия:
1) нервные импульсы должны достигать почти одновременно многих возбуждающих пресинаптических окончаний, подходящих к постсинаптическим нейронам;
2) такие активные пресинаптические окончания не должны находиться слишком далеко друг от друга;
3) область густого расположения активных синаптических окончаний должна быть достаточно велика;
4) число одновременно активируемых тормозных синаптических окончаний не должно быть слишком большим.
Все эти условия легко понять в свете представлений о вероятных механизмах синаптической передачи. Считают, что синаптические окончания одних нейронов выделяют возбуждающий медиатор, а окончания других — тормозной. Возбуждающий медиатор вызывает повышение проницаемости мембраны для ионов натрия и частичную деполяризацию мембраны, находящейся в состоянии покоя. Тормозной медиатор приводит к гиперполяризации постсинаптической мембраны с повышением проницаемости для ионов калия и хлора, но не для ионов натрия. Для синаптической передачи, т. е. возбуждения постсинаптического нейрона, необходимо, чтобы концентрация возбуждающего медиатора была достаточно высока, чтобы вызвать деполяризацию постсинаптичеокой мембраны до критического (порогового) уровня. Кроме того, должна быть деполяризована достаточно обширная область, чтобы создать эффективный «сток» для ионов натрия. Для того чтобы концентрация медиатора была достаточно высокой, он должен высвобождаться рядом близко расположенных пресинаптических окончаний, а поскольку он быстро разрушается ферментами, необходимо, чтобы высвобождение медиатора из всех окончаний происходило почти одновременно. Вряд ли последовательные разряды одиночного пресинаптического окончания приведут к накоплению достаточного количества медиатора — ведь он разрушается так же быстро, как и образуется.
Односторонняя передача и синаптическая задержка
Одна из наиболее известных черт синаптической передачи — ее направленный характер. Раздражение задних корешков спинномозговых нервов, состоящих только из афферентных волокон, приводит к возникновению разряда в чисто эфферентных передних корешках, причем направление проведения никогда не меняется па обратное. Такое действие синапса как одностороннего фильтра легко объяснить, исходя из представления о химической природе синаптической передачи; медиаторы выделяются только концевыми аппаратами аксонов. Эта особенность функции синапсов имеет первостепенное, значение для поддержания упорядоченного движения сигналов в нервной системе.
Поскольку для выделения медиатора, его прохождения через синаптическую щель и возбуждения постсинаптического нейрона необходим определенный промежуток времени, неизбежно должен существовать временной интервал между приходом импульсов к пресинаптическим окончаниям и разрядом постсинаптического нейрона. Синаптическая задержка составляет приблизительно 0,5 мсек.
Понятие о рефлекторном центре
Расположенные в центральной нервной системе мотонейроны, аксоны которых иннервируют данную мышцу, можно назвать центром активации мышцы. В последние годы стало ясно, что термин центр недостаточно точен, так как группа нейронов, регулирующая данную функцию, не обязательно образует определенное скопление, обслуживающее исключительно эту функцию. Кроме того, часто трудно или невозможно выделить какую-либо одиночную группу нейронов, которой можно было бы приписать первостепенную роль в регуляции данной функции. На самом деле при любом интегративном нервном процессе наблюдается широкое вовлечение различных частей нервной системы. Несмотря на эти факты, которые делают указанный термин неясным, мы продолжаем говорить о дыхательном центре, центре ускорения сердечного ритма и многих других. Для осуществления рефлекторных актов необходимы центральные, пусть даже диффузные, связи, и если удается выделить в центральной нервной системе группу нейронов особого назначения, ее вполне можно называть рефлекторным центром.
Облегчение и торможение
Термином облегчение обозначают повышение возбудимости в отсутствие наличного возбуждения. При действии возбуждающих импульсов нейрон облегчается или возбуждается. Зная, что каждая нервная клетка в центральной нервной системе соединена посредством синапсов со многими другими нервными клетками, легко себе представить, что импульсы, поступающие к ней по одним каналам, способны так понизить ее порог, что импульсы, достигающие ее по другим каналам, вызовут возбуждение ее, хотя без предварительного облегчения они были бы неэффективны.
Торможение, наоборот, понижает возбудимость, или повышает порог. Если при алгебраической сумме влияний активных синаптических окончаний, подходящих к данной клетке, преобладает торможение, то клетка не возбуждается.
Центральная суммация и интенсивность центрального раздражения
Мы уже отметили, что синаптическая передача зависит от суммарного влияния импульсов, возникающих в близко расположенных возбуждающих синаптических окончаниях. Аналогичный эффект суммации происходит и в тормозных синаптических окончаниях. В обоих случаях эффективность импульсов определяется концентрацией медиатора. Можно сказать, что истинные раздражители, влияющие на центральные нейроны, имеют химическую природу, и что интенсивность раздражителя эквивалентна концентрации медиатора.
В прежних исследованиях подчеркивалось различие между пространственной и временной суммацией, основанное на изучении реакции мышцы в ответ на раздражение афферентных нервов. Было показано, что одновременное раздражение двух различных афферентных нервов обычно ведет к более выраженному рефлекторному сокращению, чем при изолированном раздражении каждого из этих нервов. Понятно, что эффекты суммируются в центральной нервной системе; поскольку эти два раздражения разделены не интервалом времени, а пространственно, был предложен термин пространственная суммация. Было также показано, что несколько раздражений, наносимых последовательно на одну и ту же точку афферентного нерва, вызывают более сильное рефлекторное сокращение, чем одиночное раздражение. В этом случае раздражения разделены интервалом времени, а не пространством и более высокую эффективность повторного раздражения можно считать результатом временной суммации.
В современных учебниках по физиологии приводятся понятия временной и пространственной суммации, но теперь об их механизме известно больше, чем в то время, когда они были впервые описаны. Нервные волокна многократно ветвятся, и одно волокно может участвовать в нескольких синапсах на одном нейроне. Поскольку синаптические окончания пространственно разобщены, пространственная суммация может касаться ветвей одного и того же нейрона, заканчивающихся на другом нейроне. Пространственная суммация возникает также вследствие конвергенции на одном нейроне ветвей различных нервных волокон. Поскольку между афферентными и эфферентными нейронами обычно расположено много вставочных нейронов, а задержка при прохождении возбуждения по различным центральным цепям варьирует, импульсы достигают мотонейрона не одновременно, т. е. имеет место некоторая дисперсия во времени. Если импульсы достигают близко расположенных синаптических окончаний и если интервал между моментами их появления так мал, что медиатор, высвобождающийся ранее пришедшими импульсами, не до конца разрушается к моменту прихода последующих, то происходит накопление медиатора.
Суммация имеет место как при возбуждении, так и при торможении. Более того, именно алгебраическая суммация противоположных влияний определяет, произойдет возбуждение или торможение данного нейрона.
Суммацию можно обнаружить также и в тех случаях, когда раздражение рецепторов или афферентного нерва не вызывает очевидных изменений эфферентного разряда. Само по себе раздражение может быть подпороговым в отношении способности возбуждать мотонейроны. Но при действии совместно с эффективным раздражением это «подпороговое» раздражение усиливает его эффект. Тем самым становится очевидным, что раздражение не было подпороговым для рецепторов, но разряд последних был недостаточен для создания выраженного сдвига возбудимости центральных нейронов или по крайней мере мотонейронов. Может быть и так: два афферентных раздражителя, ни один из которых сам по себе не способен вызвать возбуждение мотонейронов, при совместном воздействии вызывают рефлекторный ответ. Это показывает, что каждое раздражение вызывает облегчение центральных механизмов и что конвергирующие эффекты облегчения складываются — центральная суммация.
Эффект отдачи и разряд последействия
После прекращения действия раздражителя, тормозящего текущую активность, последняя часто усиливается — это так называемый послетормозной эффект отдачи. Он зависит от разряда последействия, который служит как бы продолжением влияния раздражения после того, как само действие раздражителя прекратилось. Если раздражитель оказывает как возбуждающее, так и тормозное влияние на мотонейроны и если возбуждающий разряд последействия сохраняется в течение более длительного времени, чем тормозной, возникает эффект отдачи.
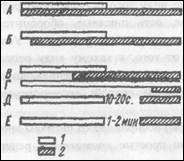
Возбуждающий и тормозной разряды последействия могут быть весьма продолжительными вследствие существования в центральной нервной системе открытых (множественных) цепей и замкнутых реверберируюишх контуров (фиг. 1). Можно видеть, что при наиболее прямых связях между афферентными и эфферентными нейронами имеет место минимальная синаптическая задержка. Импульсы, идущие по менее прямым путям, достигают мотонейронов после продолжительного лаг-периода, причем некоторые приходят к мотонейронам уже после прекращения раздражения и передачи импульсов по более прямым путям. Этот эффект сильнее

Фиг. 1. Два основных вида цепей в центральной нервной системе. А. Открытая (множественная) цепь. В. Замкнутая цепь (реверберирующий контур). Второй контур на фиг. Б (тонкие линии) иллюстрирует возможность взаимодействия многих цепей и поддержание активности в течение значительного времени.
Выражен в реверберирующих контурах, чем в открытых цепях, поскольку для первых характерно наличие обратных связей, обеспечивающих повторное самовозбуждение. С такой реверберацией, происходящей в мозге, связывают кратковременную память. До тех пор пока разряд последействия продолжается, он влияет на возбудимость центральных клеток и, таким образом, участвует в регуляции соотношения между входными и выходными сигналами в центральной нервной системе. Более того, возможно, что по крайней мере, в мозге изменения возбудимости не обязательно прекращаются, когда кончается разряд последействия, но могут сохраниться в какой-то форме неопределенно долгое время. Очевидно, не существует более адекватного объяснения для существования долговременной памяти, которая не исчезает даже после резкого снижения активности мозга, как, например, при глубоком наркозе или коме.
Фракционирование
Ни один афферентный нерв не способен вызвать возбуждение всех мотонейронов данной мышцы. Различные афферентные нервы контролируют отдельные части мышц (фракции); именно это и имеют в виду нейрофизиологи, когда говорят о «фракционировании». Некоторые нервы контролируют большие фракции мышц, другие — значительно меньшие. Кроме того, существует центральное перекрытие, в результате которого каждый мотонейрон возбуждается несколькими афферентными нервами. Это центральное перекрытие зон ведет к окклюзии.
Окклюзия
Окклюзию можно назвать эффектом «занятой линии». Если одно раздражение вызвало максимальное возбуждение центральных нейронов, дополнительные раздражения ничего не добавляют к возникшей реакции. Например, раздражение любого из двух афферентных нервов может вызвать значительное рефлекторное сокращение мышцы. Если сокращение, возникающее в результате раздражения обоих нервов, идентично тому, которое можно получить при изолированном раздражении каждого из них, это будет указывать на 100%-ную окклюзию, зависящую от полного перекрытия зон. Активность мотонейронов регулирует тот нерв, который первым их возбуждает. Такая степень перекрытия вряд ли вероятна. Значительно более обычный результат совместного раздражения нервов выражается в возникновении сокращения, которое будет несколько больше, чем при раздражении каждого из нервов в отдельности, но меньше, чем сумма сокращений, вызываемых их раздельным раздражением. Степень окклюзии можно оценить, если вычесть из суммы напряжений, вызываемых каждым первом в отдельности, напряжение, возникающее в результате одновременного их раздражения.
Количественная рефлекторная регуляция эфферентных нейронов
Рефлекторная реакция находится в количественной зависимости от интенсивности раздражения и размеров раздражаемого рецепторного поля. В определенных пределах чем сильнее раздражение, тем сильнее реакция.
Интенсивность раздражения, действующего на рефлекторный центр, определяется числом афферентных импульсов, приходящих к нему в единицу времени. (В данном случае мы не будем учитывать тормозных влияний.) По мере повышения интенсивности периферического раздражения соответствующим образом возрастает число афферентных импульсов в секунду, что приводит к более сильному раздражению рефлекторного центра. Возникает вопрос, каким образом интенсивность центрального раздражителя определяет интенсивность двигательной реакции.
Подобно тому как при повышении интенсивности периферического раздражения активируется больше рецепторов и отдельные рецепторы разряжаются с большей частотой, более сильное центральное раздражение охватывает больше мотонейронов и повышает частоту их разрядов. Более выраженная суммация возбуждающих влияний в центральной нервной системе при действии более мощного притока афферентных импульсов зависит от накопления необходимых концентраций центрального возбуждающего медиатора в более обширной области или в большем количестве. Процесс возбуждения подчиняется закону «все или ничего»; следовательно, пи импульсы эфферентных нервов, «и импульсы в мышечных волокнах не меняются по амплитуде при изменении интенсивности центрального раздражения. (Напряжение токов действия не зависит от силы раздражения.) Однако степень суммации в рефлекторных центрах определяет силу реакции эффектора путем регуляции числа участвующих элементов (вовлечение) и частоты разрядов мотонейронов.
Влияние вовлечения остается в основном одинаковым (Независимо от того, вызывается ли ответ эффектора рефлекторным путем или раздражением двигательного нерва. Иначе обстоит дело с частотой разрядов мотонейронов при рефлекторном и прямом раздражении (раздражение двигательного нерва). В последнем случае волокла двигательного нерва возбуждаются синхронно, тогда как центральная нервная система никогда (если не считать судорожных разрядов) не посылает синхронных импульсов. Пожалуй, наиболее близок к синхронному разряд мотонейронов при коленном рефлексе, однако и в этом случае большинство мотонейронов дает повторные разряды, причем каждый из них в виде коротких залпов, следующих с особой частотой. Таким образом, усиленное напряжение, наблюдающееся при -повышении частоты раздражения двигательного нерва, не отражает истинной картины влияния частоты на реакцию мышцы в нормальных условиях. В каждый данный момент напряжение является суммой напряжений активных единиц. Даже когда все мышечные волокна отвечают лишь одиночными сокращениями с полным расслаблением между сокращениями, напряжение сухожилия может поддерживаться на вполне постоянном уровне.
Это зависит от нескольких факторов:
1) длительность одиночного сокращения значительно превышает длительность вызвавшего его мышечного импульса, так что возможность суммации напряжений множества волокон выше, чем можно предположить на основании определения токов действия;
2) поскольку импульсы, поступающие в различные мышечные единицы, не синхронизированы, одни волокна достигают максимального сокращения, в то время как другие либо еще укорачиваются, либо уже удлиняются;
3) эластичность мышечной ткани также способствует сглаживанию натяжения, передающегося на сухожилие.
С увеличением частоты сокращений повышается вероятность суммации. Если некоторые элементы отвечают частичным или полным тетанусом, вероятность суммации их сокращений с реакциями других волокон еще более возрастает.
Мы выбрали мышечные реакции для иллюстрации механизма регулирующего влияния центров спинного и головного мозга на величину реакции эффекторов. Все сказанное относится к регуляции функций любых эффекторов и иллюстрирует механизмы, посредством которых осуществляется общеизвестная зависимость между интенсивностью раздражения и силой рефлекторной реакции.
Реципрокная иннервация
На принципе реципрокной иннервации мы остановимся особо, главным образом в связи с отношениями между симпатическими и парасимпатическими центрами вегетативной регуляции. Но общее представление об этом принципе удобнее дать на примере рефлекторной регуляции функции скелетных мышц.
Центры, регулирующие антагонистические функции, соединены между собой вставочными нейронами, так что повышение активности одного из центров сопровождается понижением возбудимости центра, вызывающего противоположный эффект. Например, если раздражение афферентного нерва вызывает рефлекторное сгибание конечности, происходит торможение мотонейронов соответствующих мышц-разгибателей. Предполагается, что все афферентные нервные волокна, входящие в центральную нервную систему, оказывают возбуждающее влияние на нейроны. При реципрокной иннервации приходящие сигналы, вероятно, возбуждают два вида вставочных нейронов; одни из них оканчиваются на мотонейронах сгибателей, где выделяется возбуждающий медиатор, а другие контактируют с мотонейронами разгибателей и выделяют тормозной медиатор.
Реципрокная иннервация не ограничивается мышцами одного сустава или даже одной конечности, но распространяется также на контралатеральные конечности, где реакция также носит антагонистический характер.
Так, при широко известном перекрестном разгибательном рефлексе контралатеральное разгибание сопровождается ипсилатеральным сгибанием.
Такой характер взаимодействия между центрами типичен для всей центральной нервной системы. Например, центры вдоха и выдоха в продолговатом мозге также находятся -в реципрокных взаимоотношениях: при возбуждении одного происходит торможение другого. Центры ускорения и замедления сердечной деятельности связаны такими же взаимоотношениями. Число подобных примеров можно умножить, но принцип везде один. Указав ранее, что важнейшей характеристикой интеграции является способность осуществлять в каждый данный момент времени один основной акт, мы должны теперь подчеркнуть, что реципрокная иннервация имеет первостепенное значение для обеспечения этой способности. Сходные механизмы лежат и в основе доминирования определенных рефлексов; так, когда доминируют половые рефлексы, другие рефлекторные акты подавляются.
Хотя принцип реципрокности и имеет, большое значение, не следует думать, что он является основным фактором интеграции. Его можно преодолеть, например, при произвольном напряжении конечности, когда сокращаются как сгибатели, так и разгибатели и суставы оказываются неподвижными, или при прыжке с одновременным сокращением мышц-разгибателей обеих ног.
Заключение
Все функции человека и животных связаны с изменениями в структурах организма. Жизнь требует расходования энергии для поддержания организации, без которой живой организм был бы лишен способности поддерживать возбудимость и соответствующим образом приспосабливаться к изменяющимся условиям окружающей среды. Это приспособление осуществляется главным образом эффекторами — мышцами и железами, а их деятельность зависит от нервной системы, которая собирает информацию, касающуюся окружающей среды, разрабатывает программу действия и направляет к эффекторам сигналы, регулирующие их активность. Различные виды рецепторов, каждый из которых высокочувствителен к определенной форме энергии и направляет сигналы по определенному нервному пути, обеспечивают первичный, анализ изменений в окружающей среде. Вид раздражения и расположение раздражаемой рецепторной зоны определяются главным образом на основе топографического разделения афферентных путей. Рецептор кодирует различия в интенсивности раздражения различным числом афферентных импульсов в единицу времени. Центральные рефлекторные механизмы на основе получаемой ими количественной и качественной информации определяют число, вид и расположение требующихся эффекторных элементов и интенсивность, с которой они должны реагировать. Реципрокные отношения между антагонистическими рефлекторными центрами исключают возможность хаотического и неэкономного соперничества эффекторов. Вместо этого в результате взаимодействия различных частей нервной и эндокринной системы происходит интеграция функций, позволяющая организму осуществлять одно основное действие в каждый данный момент времени.
Литература
1. Чайченко Г.М., Харченко ПД. Физиология высшей нервной деятельности. Киев, 1981.
2. Прибрам К. Языки мозга. М., 1975.
3. Воронин Л.Г. Физиология высшей нервной деятельности. М., 1979.
4. Соколов Е.Н. Физиология высшей нервной деятельности. М., 1981.
5. Шеперд Г. Нейробиология. М., 1987.
6. Хомская Е.Д. Нейропсихология. М., 1987.
Похожие работы




... подход к оценке механизмов работы высших отделов головного мозга с использованием условных рефлексов был столь успешным, что позволил Павлову создать новый раздел физиологии — «Физиологию высшей нервной деятельности», науку о механизмах работы больших полушарий головного мозга. БЕЗУСЛОВНЫЕ И УСЛОВНЫЕ РЕФЛЕКСЫ Поведение животных и человека представляет собой сложную систему взаимосвязанных ...
... только один половой инстинкт. Во-вторых, потому, что в человеческом поведении исключительная роль принадлежит социальным реакциям — поведению, определяемому положением человека в обществе, коллективе, социальной среде. К сожалению, зарубежные физиологи зачастую строят свои нейрефизиологические концепции, исходя из психоанализа Фрейда. [функцию коры, общую для человека и животных, обуславливаю- ...
... связь различными (в час-тности афферентными и эфферентными) нейронами. Благодаря многочисленным разветвлениям аксона, промежуточные нейроны могут одновременно возбуж-дать большое число других нейронов. 77. Физиология спорта. Соврем. сос-тояние, перспективы развития. Общие проблемы и задачи. Понятие о физи-ологич. резервах, классификация. Возможности управления и развития. Спорт. физиология ...
... ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ (два занятия) Занятие 1-е ПРИРОДА ВОЗБУЖДЕНИЯ 1.Что называют раздражимостью и возбудимостью? 2.Каково соотношение понятий раздражимость и возбудимость? Какие ткани в физиологии называют возбудимыми,какие- невозбудимыми? 3.Клетки каких тканей организма являются возбудимыми и невозбудимыми? 4.Дайте определение понятию "раздражитель". 5.Назовите два вида ...
0 комментариев