ХАРКІВСЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ
імені В.Н. КАРАЗІНА
УДК: 616-009-092.9
ВПЛИВ ЕМОЦІЙНОГО СТРЕСУ НА ЦЕРЕБРАЛЬНУ НЕЙРОДИНАМІКУ ТА ВАРІАБЕЛЬНІСТЬ СЕРЦЕВОГО РИТМУ ЩУРІВ
03.00.13 - фізіологія людини і тварин
Автореферат
дисертації на здобуття наукового ступеня
кандидата біологічних наук
Харків – 2008
Дисертацією є рукопис
Робота виконана в НДІ біології Харківського національного університету імені В.Н. Каразіна Міністерства освіти і науки України та в НДІ охорони здоров’я дітей та підлітків АМН України, м. Харків.
Науковий керівник: доктор медичних наук, професор
Майоров Олег Юрійович,
Харківська медична академія післядипломної освіти
МОЗ України,
завідувач кафедри клінічної інформатики та інформаційних
технологій в управлінні охороною здоров’я
Офіційні опоненти: доктор біологічних наук, професор
Воробйова Тамара Михайлівна,
Інститут неврології, психіатрії та наркології
АМН України, м. Харків,
завідувач лабораторії нейрофізіології та імунології
доктор медичних наук, професор
Міщенко Віталій Петрович,
Українська медична стоматологічна академія
МОЗ України, м. Полтава,
професор кафедри нормальної фізіології
Захист відбудеться “ 13 ” 02 2008 р. о 1515 годині на засіданні спеціалізованої вченої ради Д 64.051.17 Харківського національного університету імені В.Н. Каразіна Міністерства освіти і науки України за адресою: 61077, м. Харків, пл. Свободи, 4, ауд. III-15.
З дисертацією можна ознайомитись у Центральній науковій бібліотеці Харківського національного університету імені В.Н. Каразіна (61077, м. Харків, пл. Свободи, 4).
Автореферат розісланий “ 11 ” 01 2008 р.
Вчений секретар спеціалізованої ради В.M. Дзюба
ЗАГАЛЬНА ХАРАКТЕРИСТИКА РОБОТИ
Актуальність теми. Найменш вивченим аспектом проблеми емоційного стресу є його церебральні механізми, які знаходять висвітлення в біоелектричній активності мозку [Ведяев Ф.П., Воробьёва, 1983; Майоров О.Ю., 1989; Судаков К.В., 1994]. Не викликає сумніву, що в основі нейрофізіологічних механізмів, які обумовлюють розвиток якісно нового стану мозку в умовах емоційного стресу, лежать зміни його системної діяльності [Майоров О.Ю., 1989]. У той же час, дослідження ЕЕГ корелятів формування стійких змін функціонального стану, системної нейродинаміки в умовах емоційного стресу одиничні.
Процеси самоорганізації, які лежать в основі адаптації до емоційного стресу, знаходять відображення на ЕЕГ [Jensen B.H., 1996]. На сьогодні результати досліджень свідчать про те, що здоровий мозок має хаотичну динаміку функціонування, яка властива інтеграційній діяльності мозку [Elbert T., 1994; Jensen B.H., 1996; Palus M., 1999; Майоров.О.Ю. и др., 2006]. Однак, дослідження ЕЕГ методами детерміністського хаосу небагаточисельні.
В даній роботі проведені дослідження параметру, який характеризує динаміку систем мозку в часі, на основі оцінки ентропії Колмогорова-Сіная (еКС) по ЕЕГ в умовах спокійного неспання, гострого і хронічного емоційного стресу.
Для об'єктивної оцінки напруги систем регуляції й рівня емоційного стресу випробуваних тварин проводилася кількісна оцінка нейроетологічних показників у тесті “відкрите поле” [Майоров О.Ю., 1989], а також реєстрація варіабельності серцевого ритму (ВСР) [Mayorov O.Yu., Baevsky R.M., 1999; Баевский Р.М., 2004] на різних етапах формування емоційно-стресового стану. Це було необхідно для адекватної фізіологічної інтерпретації нового нелінійного параметру ЕЕГ – ентропії Колмогорова-Сіная.
Численні літературні джерела вказують на залучення нервових механізмів, що модулюють серцевий ритм. Сучасні методи аналізу варіабельності серцевого ритму дозволяють оцінити міру і механізми адаптації або збій у системах регуляції, використовуючи методи часового аналізу (статистичні і геометричні) та аналізу хвильової структури ритму серця на етапах формування емоційного стресу. У той же час, результати дослідження впливу стресу на варіабельність серцевого ритму, особливо на хвильову структуру ритму, залишаються суперечливими. У зв'язку з цим, є важливим і актуальним вивчення особливостей регуляції серцевого ритму в умовах гострого й хронічного емоційного стресу.
Зв’язок роботи з науковими програмами, планами, темами. Дисертаційна робота виконана відповідно до планів науково-дослідних робіт НДІ біології Харківського національного університету імені В.Н. Каразіна в рамках цільової програми “Дослідження механізмів накопичення епігенетичної інформації у процесі старіння та способи збільшення середньої тривалості життя” (№ держреєстрації 0103U004280) в співробітництві з НДІ охорони здоров’я дітей та підлітків АМН України.
Мета і завдання дослідження. Метою цього дослідження було вивчення стану церебральної нейродинаміки на основі оцінки ентропії Колмогорова-Сіная по ЕЕГ та кількісна оцінка напруження систем регуляції серцевого ритму в умовах гострого та хронічного емоційного стресу на основі оцінки варіабельності серцевого ритму щурів-самців лінії Вістар.
Відповідно до цілей дослідження вирішувалися наступні завдання:
Виявити значення еКС по ЕЕГ статевозрілих щурів-самців лінії Вістар характерні для вихідного стану та в умовах гострого і хронічного емоційного стресу.
Виявити регіональні і міжкульові відмінності ентропії Колмогорова-Сіная в умовах гострого і хронічного емоційного стресу.
Оцінити асиметрію еКС в умовах спокійного неспання, короткочасової іммобілізації і хронічного емоційного стресу в умовах вільної поведінки та у стані іммобілізації.
Оцінити напруження систем регуляції серцевого ритму в умовах емоційного стресу.
Визначити особливості поведінки в тесті “відкрите поле” до початку моделювання стресу та у стані хронічного стресу.
Об’єкт дослідження: церебральна нейродинаміка та стан систем регуляції серцевого ритму в умовах вільної поведінки та в умовах гострого і хронічного емоційного стресу, викликаного іммобілізацією, у статевозрілих щурів-самців лінії Вістар.
Предмет дослідження: визначення показників нелінійної нейродинаміки та показників варіабельності серцевого ритму в умовах вільної поведінки та в умовах гострого і хронічного емоційного стресу у статевозрілих щурів-самців лінії Вістар.
Методи дослідження: для моделювання стійкого емоційно-стресового стану використовувалась модель іммобілізаційного стресу. Нейроетологічні показники визначали у тесті “відкрите поле”. Реєструвались ЕЕГ і ЕКГ. Електроди для реєстрації ЕЕГ імплантували стереотаксичним методом; верифікацію положення електродів здійснювали на фронтальних гістологічних срізах мозку. Проводили нелінійний багаторозмірний аналіз ЕЕГ і аналіз варіабельності серцевого ритму в стані спокійного неспання та в умовах гострого і хронічного емоційного стресу.
Наукова новизна одержаних результатів. Встановлені діапазони значень ентропії Колмогорова-Сіная ЕЕГ в симетричних ділянках півкуль головного мозку і структурах лімбіко-ретикулярного комплексу статевозрілих щурів-самців лінії Вістар у стані спокійного неспання в умовах вільної поведінки.
Досліджена динаміка еКС в умовах гострого емоційного стресу. Виявлено збільшення еКС у фронтальній, парієтальній та окципітальній коркових ділянках лівої півкулі. У структурах лімбіко-ретикулярного комплексу найбільш виражене збільшення еКС відбувалось в обох ретикулярних ядрах покришки. При цьому відсутність рухомої активності відображується значним зниженням еКС в правій сенсомоторній корковій ділянці.
В умовах вільної поведінки після тривалої іммобілізації значно підвищується рівень еКС. Найбільш виражене збільшення еКС відбувається в правій парієтальній та в обох фронтальних коркових ділянках відносно вихідного рівня, а в лімбіко-ретикулярних структурах – в ретикулярних ядрах покришки і в лівому дорсальному гіпокампі та лівій мигдалині.
Іммобілізація на тлі хронічного емоційного стресу викликає зниження еКС у досліджуваних коркових і лімбіко-ретикулярних утвореннях головного мозку; значення еКС у цих умовах значно нижче вихідного рівня, визначеного у інтактних тварин. Тільки у правій мигдалині рівень еКС збільшується.
Оцінка Колмогорова-Сіная по ЕЕГ у стані спокійного неспання та її зміна в умовах моделювання емоційних стресів дозволяє виявити структури, які залучаються при формуванні стійкого емоційно-стресового стану.
В стані іммобілізації в умовах гострого та хронічного стресу відбувається зміщення вегетативного балансу в бік переваги симпатичної активності, зменшується варіабельність серцевого ритму, знижується загальна спектральна потужність і потужність в усіх частотних діапазонах. В умовах гострого й хронічного стресу підвищується роль центрального контуру управління у порівнянні з автономним, підсилюється стабілізуючий вплив вищих відділів мозку.
Тривала іммобілізація супроводжується стійкими змінами систем регуляції серцевого ритму, що може свідчити про емоційно-стресовий стан досліджуваних тварин.
Практичне значення одержаних результатів. Результати, одержані в цій роботі, поглиблюють фундаментальні знання церебральної нейродинаміки і напруження систем регуляції роботи серця в умовах вільної поведінки у стані спокійного неспання, в умовах гострого і хронічного емоційного стресу та можуть бути використані при розробці критеріїв оцінки функціонального стану досліджуваних систем в нейрофізіологічних, нейрофармакологічних та психофізіологічних дослідженнях.
Особистий внесок здобувача. Вибір теми дисертаційної роботи, постановка мети і задач, вибір об'єкта досліджень, методів досліджень адекватних рішенню задач та інтерпретація отриманих даних проведені разом з науковим керівником.
Автор провів патентно-інформаційний пошук і аналіз наукової літератури по даній проблемі, виконав експериментальні дослідження: стереотаксичні операції, реєстрацію ЕЕГ, ЕКГ, морфологічну верифікацію локалізації електродів у структурах мозку. Аналіз отриманих даних проведено автором самостійно за допомогою новітніх математичних та інформаційних технологій.
Апробація результатів дисертації. Основні положення дисертації висвітлювалися на міжнародній конференції “Комп’ютерна медицина’2004” (Харків, 2004), 8-th Multidisciplinary International Conference of Biological Psychiatry "Stress and Behavior" (St-Peterburg, 2004), 9-th Multidisciplinary International Conference of Biological Psychiatry "Stress and Behavior" (St-Peterburg, 2005), міжнародній конференції “Комп’ютерна медицина’2005” (Харків, 2005), 1-му З’їзді Фізіологів СНД (Сочі, 2005), другій всеукраїнській науково-технічній конференції студентів, аспірантів і молодих вчених “Актуальні питання теоретичної та прикладної фізики та біофізики. Фізика. Біофізика-2006” (Севастополь, 2006), 1-й міжнародній конференції молодих вчених “Біологія: від молекули до біосфери” (Харків, 2006), IV Всероссийской с международным участием школы-конференции по физиологии мышц и мышечной деятельности “Инновационные направления в физиологии двигательной системы и мышечной деятельности” (Москва, 2007), міжнародній конференції “Комп’ютерна медицина’2007” (Харків, 2007).
Публікації. За результатами дисертації опубліковано 11 робіт (3 статті та 8 тез).
Структура та обсяг дисертації. Матеріали викладено на 141 сторінці друкованого тексту. Дисертаційна робота складається зі вступу, розділів: літературний огляд, матеріали і методи досліджень, результати досліджень, обговорення результатів, узагальнення результатів, підсумкових висновків та переліку використаної літератури із 251 найменування. Матеріали дисертації проілюстровано 16 рисунками і 8 таблицями.
ОСНОВНИЙ ЗМІСТ РОБОТИ
Літературний огляд. У першому розділі роботи дається обсяг літератури за темою дисертації. В огляді надано сучасне уявлення про фізіологічні механізми емоційного стресу, дослідження поведінки в умовах стресу на підставі нейроетологічних показників в тесті “відкрите поле” та фізіологічні механізми регуляції серцевого ритму. Особливу увагу приділено характеристиці церебральної нейродинаміки на підставі методів нелінійної динаміки і детермінованого хаосу.
Матеріали та методи дослідження. Дослідження проведені в хронічному експерименті на 104 щурах-самцях лінії Вістар шестимісячного віку.
Для формування стійкого емоційно-стресового стану використовувалася модель іммобілізаційного стресу. Тварин прив'язували за лапи до дерев'яної підстави 26x15 см в положенні на животі, що дозволяло реєструвати ЕЕГ. Тварини знаходилися в умовах моделі протягом 5 днів по 5 годин щодня. Дослідження проводилися на перший і п'ятий день для вивчення механізмів формування гострого і хронічного емоційного стресу. У ці дні досліджувані показники реєструвалися перед початком і через 2,5 години від початку іммобілізації.
За 5 - 6 днів до реєстрації ЕЕГ тваринам стереотаксичним методом імплантували ніхромові електроди діаметром 0,1 мм у лаковій ізоляції. Операція проводилася під загальною анестезією (внутрішньочеревно 5% тіопенталу натрію в дозі 0,2 мл на 100 г ваги) з використанням стереотаксичного апарату СЕЖ-2. Стереотаксичні координати визначалися за атласом [Fivkova E. et al., 1967]. ЕЕГ реєструвалася монополярно з референтним усередненим електродом [Goldman D., 1950]. Кожна подія записувалася протягом 120 с. Використовувалися симетричні коркові і лімбіко-ретикулярні відведення: 1) коркові ділянки – фронтальні, сенсомоторні, парієтальні й окципітальні; 2) лімбіко-ретикулярні структури – мигдалеподібний комплекс, дорсальний гіпокамп (поле СА1), ретикулярна формація середнього мозку (ретикулярне ядро покришки). Реєстрація ЕЕГ і ЕКГ проводилася 16-канальним електроенцефалографом угорської фірми “Medicor”. Відведення ЕЕГ і ЕКГ у комп'ютер здійснювалося за допомогою 16-канального аналого-цифрового перетворювача (АЦП) фірми “Advantech” з частотою дискретизації 200 Гц на канал.
Для нелінійного аналізу ЕЕГ вибирали безартефактні стаціонарні ділянки ЕЕГ тривалістю 35-40 с. Керування енцефалографом і нелінійний багаторозмірний аналіз ЕЕГ здійснювали за допомогою системи комп'ютерної ЕЕГ NeuroResearcher®'2005 [Mayorov.O.Yu. еt al., 2003, Майоров О.Ю., 2005]. Ентропія Колмогорова-Сіная (еКС, біт/с) обчислювалася на підставі спектра показників Ляпунова і визначалася як усереднена по фазовому простору сума позитивних показників Ляпунова Sm. Для обчислення затримки (delay), яка істотно впливає на визначення ентропії Колмогорова-Сіная, був використаний метод оцінки розмірів осей відновленого у фазовому просторі атрактора [Майоров О.Ю. и др., 2006].
Реєстрацію й аналіз ЕКГ здійснювали за допомогою системи для комп'ютерного аналізу варіабельності серцевого ритму Cardio-Tension-Test'2005 [Mayorov O.Yu., Baevsky R.M., 1999; Майоров.О.Ю., 2005]. Вибиралися ділянки ЕКГ тривалістю одна хвилина. Для аналізу ВСР використовували загальноприйняті показники: частота серцевих скорочень (ЧСС, уд./хв), мода (Мо, с), амплітуда моди (АМо, %), сигма (у, с), індекс “напруги” (ІН), індекс “тип реакції напруги” (ТРН), варіаційний розмах (ДХ, с), нормований варіаційний розмах (ДХ/M, %), коефіцієнт варіації кардіоінтервалів (V, %), частота дихання (ЧД, дих./хв), показник адекватності процесів регуляції (ПАПР), стандартне відхилення різниць між сусідніми кардіоінтервалами (SDSD, с), ексцес (Ex), трикутний індекс ВСР, загальна потужність спектра (TSP, мс2), спектральна потужність низькочастотного компонента ВСР (LF, мс2), спектральна потужність низькочастотного компонента ВСР в нормованих одиницях (LFnorm, %), спектральна потужність високочастотного компонента ВСР (HF, мс2), спектральна потужність високочастотного компонента ВСР в нормованих одиницях (HFnorm, %), відношення потужності спектра низько- та високочастотного компонента ВСР (LF/HF) [Баевский Р.М., 1984, 2004; Майоров О.Ю., 1988]. Індекс “напруги” визначали як ІН = AMo/у, а індекс, що відображує “тип реакції напруги”, як ТРН=AMo/у*Мо [Майоров О.Ю., 1988].
Для визначення особливостей поведінки інтактних тварин і їх реакції на перебування в умовах хронічного емоційного стресу оцінювали нейроетологічні параметри в тесті “відкрите поле” [Walsh R.N. et al., 1976]. Усі тварини тестувалися у “відкритому полі” на протязі 6 хвилин двічі: перед початком моделювання емоційного стресу та в останній день досліджень перед іммобілізацією (після чотирьох днів стресування).
Для оцінки вірогідності змін усіх досліджуваних показників в умовах стресу застосовувався непараметричний критерій “U” Вілкоксона-Мана-Уітні за допомогою пакета прикладних програм “Statgraphics 5.0, Plus”.
РЕЗУЛЬТАТИ ДОСЛІДЖЕННЯ ТА ЇХ ОБГОВОРЕННЯ
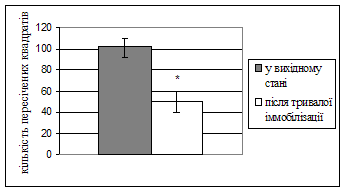
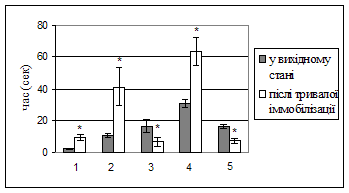
Нейроетологічні показники в тесті “відкрите поле” в нормі та в стані хронічного емоційного стресу. Тестування щурів у “відкритому полі” після перебування в умовах тривалої іммобілізації виявило зміни поведінки, які можна інтерпретувати як наслідок формування хронічного емоційно-стресового стану (рис. 1).
 А
А
 В
В
Рис. 1. Показники тесту “відкрите поле” статевозрілих щурів-самців до і після 4-х днів іммобілізації. A: 1 – період першого переміщення, 2 – час перебування в центрі площадки на початку тесту, 3 – час виходів до центру площадки, 4 – час грумінгу, 5 – вертикальна активність. B – амбуляція. * - p < 0,001.
Іммобілізаційний стрес вірогідно збільшував період першого переміщення з 2,35±0,26 с до 9,34±2,09 с (р < 0,001) після поміщення тварин у центр поля. Збільшився час перебування на центральній площадці на початку тесту з 10,59±1,23 с до 41,2±12,11.с (p = 0,001) і час грумінга – з 30,5±2,9 с до 63,41±8,64 с (p = 0,00003). Зменшилась амбуляція (горизонтальна активність) з 101,61±7,69 до 49,48±9,73 (p = 0,00015) пересічених квадратів та час вертикальної активності – з 16,22±1,39 с до 7,16±1,47 с (p.<.0,0002). Показники варіабельності серцевого ритму щурів у вихідному стані та в умовах емоційного стресу. Результати цього розділу роботи представлені в таблицях 1-4.
У вихідному стані в нормі гомеостаз основних систем організму забезпечується при мінімальній напрузі регуляторних механізмів. Таким чином, значення більшості показників варіабельності серцевого ритму (ВСР) не повинні перевищувати визначених порогів, встановлених для конкретної віково-статевої групи щурів.
Таблиця 1. Математико-статистичні показники варіабельності серцевого ритму щурів в умовах емоційного стресу
| Показник ВСР | До іммобілізації | Після 2,5 годин іммобілізації на 1-й день стресу | Після 4-х днів іммобілізації | Після 2,5 годин іммобілізації на 5-й день стресу |
| ЧСС, уд./хв | 381,45±8,75 | 440,23±5,78** | 382,60±10,42 | 434,82±8,10**,xx |
| Мо, с | 0,16±0,0036 | 0,14±0,002** | 0,16±0,0044 | 0,14±0,0027**,xx |
| AМo, % | 51,21±2,69 | 63,08±2,97** | 54,22±2,91 | 61,09±3,06 |
| s, с | 0,007±0,0005 | 0,0044±0,0003** | 0,0072±0,0006 | 0,0054±0,00048 |
| ΔХ, с | 0,04±0,0026 | 0,028±0,0023* | 0,038±0,0027 | 0,034±0,0024 |
| V, % | 4,44±0,31 | 3,13±0,21** | 4,38±0,35 | 3,86±0,32 |
| ΔX/M, % | 24,16±1,61 | 20,21±1,37 | 23,89±1,54* | 22,41±1,77 |
| ЧД, дих./хв | 72,36±3,88 | 80,87±3,94 | 63,58±4,43 | 73,88±5,26 |
| ПАПР | 327,56±22,26 | 464,99±25,83** | 347,51±23,52 | 441,53±26,00**,xx |
| SDSD, с | 0,006±0,0004 | 0,0046±0,0002** | 0,0056±0,0003 | 0,0049±0,00028 |
Примітки:
1. *,x p<0,05; **,xx p<0,01 – вірогідність розходжень;
2. *відносно вихідного стану;
3. x відносно стану хронічного стресу (після 4-х днів іммобілізації).
В умовах гострого стресу, а також у стані іммобілізації на п’ятий день стресу крива розподілу різко зміщується по осі абсцис в лівий бік, зменшується ширина гістограми (варіаційний розмах), крива стає значно вище і більш загострена вгорі – амплітуда моди (АMо), індекс “напруги” (ІН) й ексцес збільшуються. Подібні зміни вказують на посилення симпатичного тонусу в результаті збільшення емоційного стресу [Баевский.Р.М., 1984]. Кінцевий ефект регуляції проявляється у вигляді тахікардії. Підвищується активація симпатоадреналової системи, що впливає на серце одночасно через нервовий і гуморальний канали (збільшуються АMо і частота серцевих скорочень (ЧСС)). Амплітуда моди відображує стабілізуючий ефект централізації управління ритмом серця, обумовлений, в основному, ступенем активації симпатичного відділу вегетативної нервової системи [Баевский Р.М. и др., 1997].
У нашому дослідженні збільшився індекс “напруги” і показник “тип реакції напруги” (ТРН). Індекс “напруги” відображує співвідношення між активністю симпатичного та парасимпатичного відділів вегетативної нервової системи без гуморальних впливів і співвідношення центрального контуру регуляції (АМо) з автономним контуром (у). Збільшення ІН обумовлене посиленням симпатичних впливів (АМо) з одночасним зменшенням парасимпатичного тонусу (у). Показник ТРН, поряд з вищевикладеним для ІН, дозволяє оцінити рівень функціонування синусного вузла за рахунок гуморальних впливів – дає змогу оцінити ступінь централізації управління серцевим ритмом. Значення ІН та індексу ТРН дозволяють вважати, що вегетативний баланс зміщується в умовах формування емоційного стресу в бік переваги симпатичного тонусу за рахунок активації симпатичного та гальмування парасимпатичного відділів вегетативної нервової системи – АMо збільшується, σ знижується (табл. 1).
Показник адекватності процесів регуляції (ПАПР) в умовах гострого стресу й іммобілізації на тлі хронічного стресу значно збільшився, що відображує підвищений ступінь компенсаторних адаптивних процесів регуляції.
Таблиця 2
Показники тривожності щурів в умовах емоційного стресу
| Показник ВСР | ІН | ТРН |
| До іммобілізації | 9388,62±1274,58 | 61826,28±9783,50 |
| Після 2,5 годин іммобілізації на 1-й день стресу | 16763,30±1654,35** | 124927,40±13400,90** |
| Після 4-х днів іммобілізації | 9462,64±1133,20 | 62716,80±9128,40 |
| Після 2,5 годин іммобілізації на 5-й день стресу | 13803,64±1505,89 x | 101216,50±12105,20 xx |
Примітки:
1. x p<0,05; **,xx p<0,01 – вірогідність розходжень;
2. *відносно вихідного стану;
3. x відносно стану хронічного стресу (після 4-х днів іммобілізації).
Виходячи з загальноприйнятої інтерпретації математичних показників серцевого ритму і двоконтурної моделі управління ритмом [Баевский Р.М., 1984], можна стверджувати, що гострий емоційний стрес, а також іммобілізація на тлі хронічного стресу, викликає виражену симпатикотонію, посилення центральних впливів, діючих безпосередньо на нервовий канал. Встановлюється більш високий, досить стабільний на визначеному етапі емоційного стресу, рівень гуморальних впливів (кінцевий ефект регуляції), що обумовлює середню частоту серцебиття. Підвищується роль центрального контуру управління (про що свідчить збільшення ІН і ТРН), підсилюється стабілізуючий вплив вищих відділів мозку в умовах стресу. Зменшення дихальної складової спектра (HF) характеризує зниження регуляції на рівні автономного контуру. Вегетативний гомеостаз зміщується у бік переваги симпатичного тонусу в умовах як гострого, так і хронічного стресу. Зміни показників, що спостерігаються, відображують взаємодію центрального й автономного контурів, їх протидіючий вплив на ритм серця.
Таблиця 3
Геометричні показники варіабельності серцевого ритму щурів в умовах емоційного стресу
| Показник ВСР | Ex | Трикутний індекс ВСР |
| До іммобілізації | 2,94±0,13 | 2,12±0,11 |
| Після 2,5 годин іммобілізації на 1-й день стресу | 3,96±0,47** | 1,68±0,08** |
| Після 4-х днів іммобілізації | 3,00±0,13 | 2,01±0,13 |
| Після 2,5 годин іммобілізації на 5-й день стресу | 5,11±1,84 | 1,73±0,09 |
Примітка. ** - p £ 0,01.
Спостерігається зниження показників ДХ, SDSD, ДХ/M(%), V(%), що відображає зміщення симпато-вагального балансу у бік симпатичної ланки.
Нами отримане достовірне зниження загальної потужності спектральних компонентів ВСР у стані іммобілізації, що обумовлюється низхідним впливом вищих вегетативних центрів гіпоталамо-гіпофізарного рівня. Це вказує на перехід управління в режим термінової мобілізації функціональних резервів, на домінуючий вплив центральних механізмів управління на роботу автономних. Спектральна потужність знизилася в усіх частотних діапазонах.
Достовірне зменшення в стресовому стані потужності HF діапазону зі збільшенням відношення LF/HF показує, що парасимпатична активність знизилася з домінуванням симпатичної регуляції.
Наші результати узгоджуються з дослідженнями [Inagaki H. et al., 2004], в яких було встановлено, що моделювання тривожного стану у щурів (використовувалася модель спрямованого струму повітря) веде до зниження потужності HF діапазону. LF і LF/HF вірогідно не змінилися, проте, відношення LF/HF дещо збільшилося. Ці результати підтверджують, що симпатична нервова система стала переважною в автономному балансі в умовах емоційного стресу, парасимпатична активність знизилась.
Стан хронічного стресу включає системні зміни, що є наслідком тривалої іммобілізації. Дані аналізу варіабельності серцевого ритму в умовах вільної поведінки на тлі хронічного стресу відображують стійкі зміни різних ланок регуляції. Так само слід зазначити “страх очікування” повторення іммобілізації, що сам по собі є стресором [Майоров О.Ю., 1989; Inagaki H. et al., 2004]. Слід зазначити розходження в характері реакції автономної нервової системи в різних емоційних станах. Проте, найбільш характерним є збільшення значення симпато-вагального балансу в умовах стресу. Головною причиною цього є збільшення симпатичної активності і/або зниження парасимпатичної активності [Cohen H. et al., 2000; Friedman B.H. et al., 1998]. Ці результати погоджуються зі змінами в параметрах ВСР, що спостерігаються в умовах емоційного стресу в нашому дослідженні й даними літератури [Inagaki H. et al., 2004].
Таблиця 4
Показники спектрального аналізу варіабельності серцевого ритму щурів в умовах емоційного стресу
| Показник ВСР | До іммобілізації | Після 2,5 годин іммобілізації на 1-й день стресу | Після 4-х днів іммобілізації | Після 2,5 годин іммобілізації на 5-й день стресу |
| TSP, мс2 | 11,84±1,53 | 4,11±0,77** | 11,96±2,08 | 5,43±1,08xx |
| LF, мс2 | 2,44±0,42 | 1,26±0,35 | 2,38±0,64 | 1,81±0,58 |
| LFnorm | 32,62±2,13 | 33,28±3,53 | 36,64±2,61 | 38,36±3,92 |
| HF, мс2 | 4,62±0,75 | 1,78±0,23* | 4,27±0,87 | 2,14±0,30 |
| HFnorm | 67,07±2,19 | 66,72±3,53 | 62,96±2,68 | 63,15±3,79 |
| LF/HF | 0,55±0,05 | 0,64±0,10 | 0,67±0,08 | 0,79±0,18 |
Примітки:
1. *p<0,05; **,xx p<0,01 – вірогідність розходжень;
2. *відносно вихідного стану;
3. x відносно стану хронічного стресу (після 4-х днів іммобілізації).
Таким чином, аналіз матиматико-статистичних параметрів варіабельності серцевого ритму дозволив зробити об’єктивну кількісну оцінку напруження систем регуляції щурів на різних етапах формування емоційного стресу. Це в свою чергу дає можливість коректно і диференційовано інтерпретувати новий нелінійний показник динамічних систем мозку по ЕЕГ в умовах гострого та хронічного емоційного стресу.
Оцінка нейродинамічних систем мозку щурів на підставі аналізу нелінійної динаміки ентропії Колмогорова-Сіная по ЕЕГ. Проведено дослідження змін динамічних характеристик ключових коркових і лімбіко-ретикулярних структур мозку, що беруть участь у формуванні адаптивних процесів у статевозрілих щурів-самців лінії Вістар в умовах емоційного стресу за допомогою об'єктивного кількісного показника, нелінійного параметра - ентропії Колмогорова-Сіная (еКС) по ЕЕГ.
У вихідному стані в умовах вільної поведінки середні значення ентропії Колмогорова-Сіная (еКС) в усіх досліджуваних структурах знаходилися у діапазоні 3,14±0,36 – 5,67±0,89 (табл. 5, 6). У неокортикальних ділянках середні значення еКС були в діапазоні 3,14±0,36 – 4,52±0,65. Найбільші значення характерні для окципітальних (4,49±0,45 – 4,52±0,65), парієтальних (3,72±0,4 – 4,13±0,46) та сенсомоторних ділянок (3,94±0,72 – 4,19±0,63). Найменші значення в стані спокійного неспання характерні для фронтальних ділянок мозку (3,14±0,36 – 3,27±0,36).
Таблиця 5
Ентропія Колмогорова-Сіная (еКС) по ЕЕГ коркових структур щурів
у вихідному стані та в умовах іммобілізаційного стресу
| Найменування відведення | До іммобілізації | Після 2,5 годин іммобілізації на 1-й день стресу | Після 4-х днів іммобілізації | Після 2,5 годин іммобілізації на 5-й день стресу |
| Фронтальна кора (ліва) | 3,27±0,36 | 3,86±0,68 | 4,56±0,97 | 2,77±0,54 |
| Фронтальна кора (права) | 3,14±0,36 | 3,08±0,49 | 4,34±1,49 | 2,32±0,4 |
| Сенсомоторна кора (ліва) | 3,94±0,72 | 3,87±0,74 | 4,03±1,05 | 2,8±0,71 |
| Сенсомоторна кора (права) | 4,19±0,63 | 3,01±0,63** | 3,97±0,87 | 2,35±0,6* |
| Парієтальна кора (ліва) | 4,13±0,46 | 4,85±0,66 | 4,21±0,65 | 4,41±0,54 |
| Парієтальна кора (права) | 3,72±0,4 | 3,95±0,41 | 5,72±0,94* | 3,89±0,58* |
| Окципітальна кора (ліва) | 4,52±0,65 | 4,86±0,64 | 4,46±0,57 | 4,22±0,48 |
| Окципітальна кора (права) | 4,49±0,45 | 4,34±0,37 | 5,19±0,81 | 4,05±0,39 |
Примітка. *- p < 0,05,** - p £ 0,01 – вірогідність розходжень.
У лімбіко-ретикулярних структурах середні значення ентропії Колмогорова-Сіная були в діапазоні 4,9±0,51 – 5,67±0,89, що значно вище ніж в коркових утвореннях. Найбільш висока еКС виявлена в правому ретикулярному ядрі покришки (5,27±0,59) і правому гіпокампі (5,67±0,89). Найменші значення еКС були виявлені в лівому ретикулярному ядрі покришки (4,9±0,51) і в лівому мигдалеподібному комплексі (4,95±0,46).
Є всі підстави припустити, що означений рівень хаосу відіграє позитивну роль, забезпечуючи мозку “інформаційно-багатий стан” [Elbert T. et al., 1994]. Іншими словами, забезпечує готовність мозку за рахунок здатності до самоорганізації генерувати нову адаптивну активність.
Використання методу [Майоров О.Ю. и др., 2006] визначення затримки (delay) дозволило значно підвищити надійність і повторюваність результатів і, як наслідок, дало можливість одержати діапазон значень еКС по ЕЕГ досліджуваних структур мозку щурів, характерний для тварин в стані спокійного неспання та в умовах гострого і хронічного стресу.
Таблиця 6
Ентропія Колмогорова-Сіная (еКС) по ЕЕГ лімбіко-ретикулярних структур щурів у вихідному стані та в умовах іммобілізаційного стресу
| Найменування відведення | До іммобілізації | Після 2,5 годин іммобілізації на 1-й день стресу | Після 4-х днів іммобілізації | Після 2,5 годин іммобілізації на 5-й день стресу |
| Гіпокамп дорсальний (лівий) | 5,07±0,32 | 5,20±0,35 | 5,80±1,09 | 4,91±0,33 |
| Гіпокамп дорсальний (правий) | 5,67±0,89 | 5,72±1,03 | 5,59±1,02 | 4,71±0,29 |
| Мигдалеподібний комплекс (лівий) | 4,95±0,46 | 5,26±0,37 | 5,77±0,83* | 4,46±0,58 |
| Мигдалеподібний комплекс (правий) | 5,06±0,36 | 4,82±0,47 | 5,03±0,83 | 5,88±0,54* |
| Ретикулярна формація середнього мозку (ліва) | 4,90±0,51 | 5,65±0,47 | 7,19±0,82** | 5,68±0,50 |
| Ретикулярна формація середнього мозку (права) | 5,27±0,59 | 5,76±0,27 | 7,26±0,81 | 5,81±0,49 |
Примітка. *- p < 0,05,** - p £ 0,01 – вірогідність розходжень.
Слід зазначити асиметрію цього показника у вихідному стані (табл. 7, 8) – значення еКС у дорсальному гіпокампі, мигдалині і ретикулярній формації середнього мозку праворуч вище відповідно на 11,83 %, 2,22 % і 7,55 %. У коркових ділянках півкуль значення еКС, напроти, ліворуч дещо вище, ніж праворуч: у фронтальній ділянці на 3,98.%, у парієтальній ділянці на 9,93 %, тоді як у сенсомоторній ділянці значення еКС праворуч на 6,35 % вище, ніж ліворуч. Відомо, що функціональна асиметрія сприяє нормальній інтеграційній діяльності мозку.
В умовах гострого емоційного стресу, через 2,5 години від початку першої іммобілізації, спостерігається виражене збільшення еКС у лівих фронтальній, парієтальній і окципітальній ділянках (на 18,04 %, 17,41 % і 7,54 %, відповідно) (табл..5). У структурах лімбічної системи виявлене незначне збільшення еКС в обох дорсальних гіпокампах (на 2,58 % і 0,86 %) і в лівому мигдалеподібному комплексі (на 6,23 %). Збільшення еКС відбувається в обох ретикулярних ядрах покришки (на 15,22 % і 9,25 %) (табл. 6).
Помірне підвищення рівня еКС, виявлене в деяких ділянках і структурах лімбіко-ретикулярного комплексу в умовах гострого емоційного стресу, можна інтерпретувати як активацію захисних механізмів, відображення процесів формування адаптивної активності, пристосування до нових умов – іммобілізації. В стані емоційної напруги в умовах короткочасної іммобілізації підвищуються можливості самоорганізації, збільшується здатність до формування упорядкованих адаптивних дисипативних структур і, отже, здатність до адаптації. Перебування тварин у стані нерухомості привело до різкого зменшення рівня еКС у правій сенсомоторній ділянці на 28,2 % (з 4,19±0,63 до 3,01±0,63, p<0,05), що відображає бездіяльність цієї ділянки мозку в умовах іммобілізації та погоджується з результатами оцінки нелінійних параметрів ЕЕГ під час дослідження довільних рухів людини [Popivanov D. et al., 1999; Dushanova J., 2001].
В умовах короткочасної іммобілізації у щурів зниження еКС в сенсомоторній ділянці та збільшення еКС в лімбіко-ретикулярних структурах може вказувати на зменшення числа інформаційних процесів, що відбуваються у моторній корі при іммобілізації й посилення активності лімбічних структур при гострому іммобілізаційному емоційному стресі.
Слід зазначити асиметрію значень еКС в умовах короткочасової іммобілізації в усіх коркових відведеннях (10,7 % – 22,22 %), при цьому значення були вищі в лівій півкулі (табл. 7).
Таблиця 7
Асиметрія значень еКС коркових структур (у % до лівої півкулі)
| Найменування відведення | До іммобілізації | Після 2,5 годин іммобілізації на 1-й день стресу | Після 4-х днів іммобілізації | Після 2,5 годин іммобілізації на 5-й день стресу |
| Фронтальна кора (права) | -3,98 | -20,21 | -4,82 | -16,25 |
| Сенсомоторна кора (права) | 6,35 | -22,22 | -1,49 | -16,07 |
| Парієтальна кора (права) | -9,93 | -18,56 | 35,87 | -11,79 |
| Окципітальна кора (права) | -0,66 | -10,70 | 16,37 | -4,03 |
У лімбіко-ретикулярних структурах у дорсальному гіпокампі (на 10 %) і ретикулярній формації середнього мозку (на 1,95 %) значення було вище в правій півкулі і тільки в мигдалеподібному комплексі (на 8,36%) вище ліворуч, ніж праворуч (табл. 8). Достовірна латералізація реакції в умовах гострого емоційного стресу виявлена в мигдалеподібному комплексі (рис. 2), де ліворуч значення збільшилося в середньому на 6,23 %, праворуч знизилося на 4,85 % (p < 0,04).
Для дослідження стійких змін, сформованих під впливом хронічного емоційного стресу, була проведена оцінка рівня еКС після 4-х днів іммобілізації (табл. 5, 6).
Було встановлено, що хронічний емоційний стрес значно впливав на параметри нейродинамічних систем мозку щурів, що знайшло об'єктивне відображення на ЕЕГ і в рівні еКС досліджуваних областей мозку. Стан хронічного емоційного стресу супроводжується підвищеним рівнем еКС, що відображає стадію мобілізації захисних механізмів. Значне збільшення еКС у порівнянні з вихідним рівнем виявлено в лівій і правій фронтальних ділянках кори (на 39,4 % і 38,2 %, відповідно) та в правій окципітальній корі (на 15,6 %). Найбільш сильно еКС збільшується у порівнянні з вихідним рівнем в правій парієтальній ділянці (на 53,8 %). Раніше було виявлено, що парієтальна ділянка при будь-яких емоціях сильніше активується праворуч [Crowne.D.P. et al., 1987; Davidson R.J. et al., 1999]. Права сенсомоторна ділянка практично перестає реагувати на іммобілізацію – рівень еКС майже відновлюється в порівнянні з гострим стресом (він тільки на 5,5 % нижче вихідного). Це також підтверджує її бездіяльність в умовах іммобілізації. У той же час, збільшення еКС в стані хронічного емоційного стресу, викликаного тривалою іммобілізацією, характерне для лімбічних структур лівого мозку. Так, еКС лівого дорсального гіпокампа збільшилася на 14,4 % у порівнянні з вихідним рівнем (при гострому стресі тільки на 2,6 %), у лівому мигдалеподібному комплексі – на 16,6 % у порівнянні з вихідним рівнем (при гострому стресі тільки на 6,3.%). Ще більш виражені зміни відбуваються в правому та лівому ретикулярних ядрах покришки (на 46,7 % і 37,8 %, відповідно) у порівнянні з вихідним рівнем (при гострому стресі тільки на 15,3 % і 9,3 %). В роботі [Белова Т.І., 1989] було показано, що під впливом хронічного емоційного стресу відбуваються морфологічні зміни клітинних структур і підвищення проникнення гемато-енцефалічного бар’єру в області ретикулярної формації середньго мозку. Що ж стосується участі мигдалини в формуванні емоційно-стресових реакцій і тривало утримуваних станів, то на цей рахунок в літературі є багато доказів [Ведяев Ф.П. и др., 1983; Майоров О.Ю., 1988; Коплік Е.В. та ін., 1997; Phelps E.A., 2004].
Таблиця 8
Асиметрія значень еКС лімбіко-ретикулярних структур (у % до лівої півкулі)
| Найменування відведення | До іммобілізації | Після 2,5 годин іммобілізації на 1-й день стресу | Після 4-х днів іммобілізації | Після 2,5 годин іммобілізації на 5-й день стресу |
| Гіпокамп дорсальний (правий) | 11,83 | 10,00 | -3,62 | -4,07 |
| Мигдалеподібний комплекс (правий) | 2,22 | -8,36 | -12,82 | 31,84 |
| Ретикулярна формація середнього мозку (права) | 7,55 | 1,95 | 0,97 | 2,45 |
Оцінка функціональної асиметрії за середнім рівнем еКС в стані хронічного емоційного стресу виявила регіональні особливості (табл. 7, 8). У коркових утвореннях асиметрія еКС була в діапазоні від –1,48 % до +35,87 %. Найбільша асиметрія еКС спостерігалася в парієтальній ділянці (праворуч більше на 35,87 %).
У лімбіко-ретикулярних утвореннях асиметрія еКС була в діапазоні від –12,82 % до +0,97 %. Найбільш виражена асиметрія була в мигдалеподібному комплексі (праворуч менше на 12,82 %). В інших досліджуваних лімбіко-ретикулярних структурах вона була незначною (менш ніж 5,0 %).
Досліджувалась також реакція мозку на іммобілізацію тварин в стані хронічного емоційного стресу. В умовах іммобілізації на тлі хронічного стресу відбувалося різке зниження еКС, а у деяких утвореннях значення еКС були навіть нижче рівня, характерного для інтактних тварин (табл. 5, 6). Спостерігалося зменшення значень еКС практично в усіх коркових відведеннях. В лівій та правій фронтальних ділянках півкуль значення eКС знизилися на 39,25 % і 46,54 %, відповідно, в порівнянні з рівнем еКС в умовах хронічного емоційного стресу (у порівнянні з вихідним рівнем на 15,29 % і 26,11.%, відповідно). У сенсомоторній корі рівень еКС продовжував знижуватися ліворуч і праворуч на 30,52 % і 40,94 %, відповідно. У парієтальній і окципітальній корі рівень еКС значно знизився праворуч (на 31,95 % і 22 %, відповідно). У лімбіко-ретикулярних структурах зниження еКС спостерігалося в обох дорсальних гіпокампах (на 15,38 % і 15,64 %), ретикулярних ядрах покришки (на 20,99 % і 19,99 %) і лівому мигдалеподібному комплексі (на 22,69 %). Тільки в правому мигдалеподібному комплексі рівень еКС збільшився (на 16,85 %) (рис. 2). Виявлені зміни еКС в умовах іммобілізації на тлі хронічного стресу можуть вказувати на формування домінанти і системного гальмування, які виникають в життєво важливих ситуаціях.
На підставі отриманих результатів можна зробити висновок, що в стані іммобілізації на тлі хронічного емоційного стресу відбувається зниження кількості активних паралельних функціональних процесів у мозку, що відображує процес формування поведінкової домінанти. Зниження еКС може свідчити також про "інформаційно-збіднений стан", тобто відбувається зниження інформаційного резерву, що при значному або тривалому стресовому впливі може привести до "поломок" регуляції.
Оцінка функціональної асиметрії по середньому рівню еКС виявила більші значення у лівих коркових відведеннях, ніж у правих (табл. 7). Тоді як у лімбіко-ретикулярних структурах значення були вище в правій ретикулярній формації середнього мозку і правому мигдалеподібному комплексі. Тільки в гіпокампі у лівій півкулі рівень еКС був більшим, ніж у правій (табл. 8).
Достовірна латералізація реакції на іммобілізацію в умовах хронічного стресу виявлена в парієтальній корі (рис. 2), де праворуч значення знизилося на 31,95 %, а ліворуч збільшилося в середньому на 4,88 % (p=0,004).
Слід зазначити розходження в реакції еКС на іммобілізацію в умовах гострого і хронічного емоційного стресу у правій парієтальній ділянці: +6,08 і –31,95, відповідно (p.< 0,02) і лівому мигдалеподібному комплексі: +6,23 і –22,69, відповідно (p = 0,02) (рис..2), що, імовірно, пояснюється реакцією в інтактному стані та на тлі стійкого хронічного емоційно-стресового стану церебральної нейродинаміки.
Таким чином, нелінійний параметр еКС сигналу ЕЕГ є об'єктивним показником оцінки зміни динамічних характеристик ключових коркових і лімбіко-ретикулярних структур мозку, які беруть участь у формуванні адаптивних процесів в умовах гострого і хронічного емоційного стресу.
На підставі результатів власних досліджень і даних літератури можна зробити висновок, що емоційний стрес приводить до глибоких змін функціональної активності структур лімбіко-кортикальної системи головного мозку, які супроводжуються значним напруженням систем регуляції та суттєвою зміною нейроетологічних показників.
ВИСНОВКИ
Проведено дослідження динамічних систем коркових та лімбіко-ретикулярних утворень мозку щурів за допомогою нелінійного параметру ЕЕГ – ентропії Колмогорова-Сіная в умовах вільної поведінки та на різних етапах формування гострого і хронічного емоційного стресу, що дозволило встановити ефективність такого підходу для вивчення механізмів та кількісної оцінки цих станів, з одночасною оцінкою ступеня напруженості систем регуляції за допомогою аналізу варіабельності серцевого ритму.
Встановлені діапазони значень ентропії Колмогорова-Сіная по ЕЕГ в симетричних ділянках півкуль головного мозку та структурах лімбіко-ретикулярного комплексу для інтактних статевозрілих щурів-самців лінії Вістар в умовах вільної поведінки. Ентропія Колмогорова-Сіная ЕЕГ у цих тварин має регіональні та міжпівкульові розходження. Найбільш високі показники еКС характерні для правої сенсомоторної кори та для обох окципітальних ділянок півкуль. У структурах лімбічної системи найбільш високий рівень еКС виявлений у правих дорсальному гіпокампі, мигдалеподібному комплексі і ретикулярному ядрі покришки. При цьому в коркових ділянках значення еКС вище ліворуч, а у лімбіко-ретикулярних – праворуч.
В умовах гострого емоційного стресу, викликаного короткочасною іммобілізацією, виявлено виражене збільшення еКС у фронтальній, парієтальній та окципітальній коркових ділянках лівої півкулі. У структурах лімбіко-ретикулярного комплексу найбільш виражене збільшення еКС відбувалось в обох ретикулярних ядрах покришки. При цьому відсутність рухомої активності відображується значним зниженням еКС у правій сенсомоторній корі.
Стан хронічного емоційного стресу, викликаний тривалою іммобілізацією, та за даними аналізу ВСР і тесту “відкрите поле”, супроводжується значним підвищенням рівня еКС, що відображує збільшення кількості паралельних інформаційних процесів в динамічних системах мозку, та за даними аналізу ВСР і нейроетологічних показників – підвищений рівень тривожності. У порівнянні з вихідним рівнем найбільш сильно еКС збільшується в правій парієтальній ділянці, обох фронтальних ділянках, ретикулярних ядрах покришки, лівому гіпокампі і лівому мигдалеподібному комплексі.
На фоні хронічного емоційного стресу виявлено зниження величини еКС у досліджуваних коркових і лімбіко-ретикулярних структурах головного мозку, що обумовлено формуванням домінуючих функціональних систем і системного гальмування, які супроводжуються зменшенням паралельних інформаційних процесів в домінуючих нейродинамічних системах, що відображає включення механізмів адаптації та стійкості. Тільки в правому мигдалеподібному комплексі рівень еКС збільшується, відображуючи активацію апарату негативних емоцій.
Рівень ентропії Колмогорова-Сіная по ЕЕГ може служити індикатором залучення тих чи інших ділянок мозку в процес формування гострого та хронічного емоційного стресу.
Статистичний та спектральний аналіз варіабельності серцевого ритму одночасно з оцінкою нелінійної динаміки ЕЕГ дозволяє отримати об’єктивну кількісну оцінку стану організму на різних етапах формування емоційного стресу, що сприяє адекватній інтерпретації нелінійного параметра нейродинамічних систем – ентропії Колмогорова-Сіная. Виявлено типові характеристики серцевого ритму для різних етапів формування емоційного стресу. В умовах іммобілізації в стані гострого стресу та на тлі змінених під впливом хронічного емоційного стресу властивостей нейродинамічних систем виявлено зсув вегетативного балансу у бік переваги симпатичної активності, зниження варіабельності серцевого ритму, загальної спектральної потужності та потужності в усіх частотних діапазонах. В умовах гострого та хронічного стресу підвищується роль центрального контуру керування в порівнянні з процесами саморегуляції, підсилюється стабілізуючий вплив вищих відділів мозку.
Оцінка нейроетологічних показників щурів після тривалої іммобілізації виявила зміни притаманні стану хронічного емоційного стресу: збільшилися період першого переміщення після того, як тварин помістили в центр поля, час перебування на центральній площадці на початку тесту, тривалість грумінга, зменшилися горизонтальна та вертикальна активність. Кількісна об’єктивна оцінка поведінки в тесті “відкрите поле” є ефективною для діагностики стану хронічного емоційного стресу, що сприяє фізіологічній інтерпретації параметрів детермінованого хаосу нейродинамічних систем по ЕЕГ.
СПИСОК ОПУБЛІКОВАНИХ ПРАЦЬ ЗА ТЕМОЮ ДИСЕРТАЦІЇ
1. Майоров О.Ю., Вязовская О.В. Влияние острого и хронического эмоционального стресса на уровень энтропии Колмогорова-Синая ЭЭГ // Експериментальна і клінічна медицина. – 2006. – № 4. – С. 32–37. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
2. Вязовская О.В., Майоров О.Ю. Состояние церебральной нейродинамики в условиях иммобилизационного стресса у крыс на основе оценки энтропии Колмогорова по ЭЭГ // Вісник Харківського національного університету імені В.Н. Каразіна. Серія біологія. – 2006. – Вип. 4, № 748. – С. 139–146. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
3. Майоров О.Ю., Вязовская О.В. Оценка напряжения систем регуляции работы сердца в условиях моделирования эмоционального стресса у крыс // Експериментальна і клінічна медицина. – 2007. – № 1. – С. 53–58. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
4. Применение методов нелинейного анализа (детерминистского хаоса) ЭЭГ для оценки состояния нервной деятельности / Майоров О.Ю., Глухов А.Б., Тимченко Л.Н., Следюк Д.В., Козидубова С.М., Прогнимак А.Б., Вязовская О.В. // Комп’ютерна медицина’2004: клінічна інформатика і телемедицина: міжнародна науково-практична конференція. Харків, 27-29 травня 2004 р.; Вісник Харківського національного університету імені В.Н. Каразіна. Медицина. – 2004. – Вип. 8, № 617. – С. 120–121. (Дисертант брала участь в обговоренні та узагальненні отриманих результатів).
5. Vyazovska O.V. The effects of chronic immobilization on neuroetological parameters in the “open field test” in rats // Stress and Behavior: 9-th Multidisciplinary International Conference of Biological Psychiatry. St-Petersburg, May 16-19, 2005; Psychopharmacology & Biological Narcology. – 2005. – V. 5, № 2. – P. 907.
6. Майоров О.Ю., Вязовская О.В. Применение методов нелинейного анализа ЭЭГ (детерминистского хаоса) для оценки состояния церебральной нейродинамики в условиях хронического иммобилизационного стресса у крыс // Компьютерная Медицина’2005: информационные технологии для клинической практики: научно-практический симпозиум. Харьков, 23-25 июня 2005 г.; Клиническая информатика и телемедицина. – 2005. – Т. 2, № 1. – C. 149. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
7. Майоров О.Ю., Вязовская О.В. Спектральный анализ вариабельности ритма сердца крыс в условиях иммобилизационного стресcа // Біологія: від молекули до біосфери: 1 міжнародна конференція молодих вчених. Харків, 21-23 листопада 2006 р.; Тези доповідей. – Харків, 2006. – C. 65. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
8. Майоров О.Ю., Вязовская О.В. Динамика энтропии Колмогорова по ЭЭГ в условиях острого и хронического эмоционального стресса // Актуальные вопросы теоретической и прикладной физики и биофизики. Физика. Биофизика-2006: Вторая всеукраинская научно-техническая конференция студентов, аспирантов и молодых ученых. Севастополь, 17-22 апреля 2006 г.; Материалы конференции. – Севастополь, 2006. – C. 112–114. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
9. Майоров О.Ю., Вязовская О.В. Вариабельность сердечного ритма в условиях острого стресса у крыс // Біологія: від молекули до біосфери: 1 міжнародна конференція молодих вчених. Харків, 21-23 листопада 2006 р.; Тези доповідей. – Харків, 2006. – C. 63-64. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
10. Майоров О.Ю., Вязовская О.В. Снижение уровня энтропии Колмогорова-Синая по ЭЭГ сенсомоторной коры крыс в условиях иммобилизации // Инновационные направления в физиологии двигательной системы и мышечной деятельности: IV Всероссийская с международным участием школа-конференция по физиологии мышц и мышечной деятельности. Москва, 31 января-3 февраля 2007 г.; Материалы конференции. – Москва, 2007. – С. 40-41. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
11. Майоров О.Ю., Вязовская О.В. Оценка вариабельности ритма сердца крыс в условиях острого и хронического иммобилизационного стресса // Инновационные направления в физиологии двигательной системы и мышечной деятельности: IV Всероссийская с международным участием школа-конференция по физиологии мышц и мышечной деятельности. Москва, 31 января-3 февраля 2007 г.; Материалы конференции. – Москва, 2007. – С. 37-39. (Дисертант виконувала експериментальну частину, аналізувала та узагальнювала отримані результати).
АНОТАЦІЯ
В’язовська О.В. “Вплив емоційного стресу на церебральну нейродинаміку та варіабельність серцевого ритму щурів”. – Рукопис.
Дисертація на здобуття наукового ступеня кандидата біологічних наук за спеціальністю 03.00.13 – фізіологія людини і тварин, Харківський національний університет імені В.Н. Каразіна, Харків, 2008.
Встановлені діапазони значень ентропії Колмогорова-Сіная (еКС) ЕЕГ у симетричних коркових ділянках півкуль головного мозку та структурах лімбіко-ретикулярного комплексу щурів лінії Вістар у стані спокійного неспання в умовах вільної поведінки та в умовах гострого і хронічного емоційного стресу. В умовах гострого емоційного стресу виявлено виражене збільшення еКС у фронтальній, парієтальній та окципітальній коркових ділянках лівої півкулі. У структурах лімбіко-ретикулярного комплексу найбільш виражене збільшення еКС відбувалось в обох ретикулярних ядрах покришки. При цьому відсутність рухомої активності відображується значним зниженням еКС в правій сенсомоторній корковій ділянці. Виявлено стійке підвищення еКС в умовах вільної поведінки після тривалої іммобілізації та зниження рівня еКС в умовах іммобілізації на тлі хронічного емоційного стресу. В стані іммобілізації в умовах гострого та хронічного стресу спостерігалось зменшення варіабельності серцевого ритму, зниження загальної спектральної потужності та потужності в усіх частотних діапазонах. Тривала іммобілізація супроводжувалась стійкими змінами систем регуляції серцевого ритму, імовірно, характерними для хронічного емоційного стресу.
Ключові слова: емоційний стрес, іммобілізація, ЕЕГ, нелінійний аналіз, ентропія Колмогорова-Сіная, ЕКГ, варіабельність серцевого ритму.
АННОТАЦИЯ
Вязовская О.В. “Влияние эмоционального стресса на церебральную нейродинамику и вариабельность сердечного ритма крыс”. – Рукопись.
Диссертация на соискание ученой степени кандидата биологических наук по специальности 03.00.13 - физиология человека и животных, Харьковский национальный университет имени В.Н. Каразина, Харьков, 2008.
В данной работе исследовали состояние церебральной нейродинамики и оценивали напряжение систем регуляции сердечного ритма крыс линии Вистар в исходном состоянии и в условиях острого и хронического эмоционального стресса.
Установлены диапазоны значений энтропии Колмогорова-Синая (эКС) по ЭЭГ в симметричных областях полушарий головного мозга и структурах лимбико-ретикулярного комплекса половозрелых крыс-самцов линии Вистар в состоянии спокойного бодрствования в условиях свободного поведения.
Проведено исследование изменений динамических характеристик ключевых корковых и лимбико-ретикулярных структур мозга, которые принимают участие в формировании адаптивных процессов у половозрелых крыс-самцов линии Вистар в условиях иммобилизационного эмоционального стресса.
Исследована реакция этого показателя на острый иммобилизационный стресс. Через 2,5 часа от начала первой иммобилизации наблюдалось выраженное увеличение эКС во фронтальной, париетальной и затылочной областях левого полушария. В лимбико-ретикулярных областях наиболее выраженное увеличение эКС происходило в обоих ретикулярных ядрах покрышки. При этом отсутствие двигательной активности отражалось значительным снижением эКС в правой сенсомоторной коре. Таким образом, в условиях кратковременной иммобилизации снижение эКС в сенсомоторной области и увеличение эКС в лимбико-ретикулярных структурах мозга у крыс может указывать на уменьшение числа параллельно протекающих информационных процессов в моторной коре при иммобилизации и усиление активности лимбических структур при остром иммобилизационном эмоциональном стрессе.
Выявлены характерные для хронического эмоционального стресса диапазоны значений энтропии Колмогорова-Синая в корковых и лимбико-ретикулярных структурах мозга крыс. Длительная иммобилизация сопровождалась значительным повышением уровня эКС в условиях свободного поведения, что отражает стойкие изменения параметров нейродинамических систем мозга крыс. В данном исследовании выявлено значительное увеличение энтропии Колмогорова-Синая в условиях хронического эмоционального стресса в ключевых структурах лимбической системы (миндалевидном комплексе), в ретикулярной формации среднего мозга (левом ретикулярном ядре покрышки) и в корковых областях полушарий (фронтальной и париетальной).
На фоне измененных под влиянием хронического эмоционального стресса свойств нейродинамических систем в условиях иммобилизации у крыс происходит резкое снижение эКС практически во всех исследуемых структурах. Это может свидетельствовать об активации механизмов самоограничения, характерных для нелинейных динамических систем мозга, о формировании доминирующих функциональных систем и о системном торможении.
Таким образом, нелинейный параметр – энтропия Колмогорова-Синая сигнала ЭЭГ является объективным количественным показателем изменения динамических характеристик ключевых корковых и лимбико-ретикулярных структур мозга, которые принимают участие в формировании адаптивных процессов в условиях острого и хронического эмоционального стресса.
В условиях иммобилизации в состоянии острого и хронического стресса выявлено смещение вегетативного баланса в сторону преобладания симпатической активности, снижение вариабельности сердечного ритма, общей спектральной мощности и мощности во всех частотных диапазонах, усиление централизации управления сердечным ритмом. Длительная иммобилизация сопровождается стойкими изменениями регуляции сердечного ритма, по-видимому, характерными для хронического эмоционального стресса.
Тестирование исследуемых крыс в “открытом поле” после пребывания в условиях длительной иммобилизации выявило изменения поведения, характерные для развития хронического эмоционально-стрессового состояния — увеличился период первого перемещения после того, как животное поместили в центр поля, время нахождения на центральной площадке в начале теста, продолжительность груминга, уменьшились амбулация и вертикальная активность.
Давая общую оценку воздействиям эмоционального стресса, можно сказать, что все они приводят к глубоким сдвигам функциональной активности структур лимбико-кортикальной системы головного мозга, которые сопровождаются отчетливыми этологическими и кардиоваскулярными коррелятами.
Ключевые слова: эмоциональный стресс, иммобилизация, ЭЭГ, нелинейный анализ, энтропия Колмогорова-Синая, ЭКГ, вариабельность сердечного ритма.
SUMMARY
Vyazovska O.V. The influence of the emotional stress on the cerebral neurodynamics and heart rate variability in rats. - Manuscript.
Thesis for a scientific degree of candidate of biological sciences by the speciality 03.00.13 - physiology of human and animals. - V.N. Karazin's Kharkіv National University, Kharkіv, 2008.
The ranges of Kolmogorov-Sinay entropy (KSe) values of the EEG signal in the symmetric cortical areas of the cerebral hemispheres and the limbic-reticular complex structures for freely behaving adult male Wistar rats during quiet wakefulness, in conditions of acute and long emotional stress were established. An increased KSe was found in the frontal, parietal and occipital cortical areas of the left hemisphere and in the midbrain reticular formation (in the both reticular nuclei of tegmentum) in conditions of acute emotional stress. An absence of locomotor activity was reflected in the significant decrease of KSe in the right sensorimotor cortex. The stable increase in KSe in the conditions of free behavior after long immobilization in the cortical and limbic-reticular brain structures was observed. Under immobilization conditions on the background of chronic emotional stress there was response in the form of the reduction of KSe. A shift of the vegetative balance towards prevalence of sympathetic activity, a decrease in heart rate variability, a decrease in total spectral power and the power in all frequency ranges in response to the immobilization under acute and chronic stress condition were observed.
Key words: emotional stress, immobilization, EEG, nonlinear analysis, Kolmogorov-Sinay entropy, ECG, heart rate variability.
Підписано до друку 28.12.07. Формат 60х90/16
Папір офсетний. Друк ризографія.
Ум. друк. арк. 0,9. Тир. 100 прим. Зам. № 307-07.
Надруковано СПД ФО Бровін О.В. Св-во 2708608999.
м. Харків, майдан Свободи, 7. Т. (057) 758-01-08, (8066) 822-71-30
СТИЛЬ______
______ИЗДАТ
типография
0 комментариев