Вступ
Розділ 1. Загальна характеристика мохоподібних та їх різноманітність на Україні
1.1 Дослідження Мохоподібних на Україні
1.2 Умови зростання та географічне походження мохоподібних
Розділ 2. Характеристика відділу Мохоподібні
2.1 Особливості відділу Мохоподібні
2.2 Особливості основних класів відділу Мохоподібні
Розділ 3. Роль мохоподібних в житті людини та природи та охорона
Висновок
Список використаних джерел
Вступ
Мохоподібні - велика група зовнішньо надзвичайно різноманітних рослин. Представники цього відділу - найдавніші сучасні вищі рослини. Вони є найпримітивнішими серед спорових рослин і викликають певний науковий і методичний інтерес. Серед вищих рослин за кількістю видів мохоподібні займають друге місце після квіткових. В усьому світі зростає від 2200 до 27000 видів мохоподібних (за різними джерелами) [2,4], в тому числі біля 8500 печіночників (280 родів), біля 14500 (18000 видів) листкостеблових мохів і біля 300 видів (3 чи 6 родів) антоцеротових. В Україні зустрічається близько 580 видів. На поліссі зростають 368 видів мохів, в Лісостепу знайдено їх майже вдвічі менше, а в Степу й ще менше, лише Карпати можуть зрівнятися в цьому з Поліссям [19].
Мохоподібні своєрідні і неповторні за своєю організацією та способом життя, тому й заслуговують на певну увагу в шкільному курсі біології. Актуальним питанням сучасної методики біології є шляхи підвищення розвитку розумових та творчих здібностей школярів, підвищення пізнавальної активності учнів, вивчення ефективності інноваційних підходів навчання і виховання, що дають змогу не тільки дати учням основи знань, але й навчити їх обробляти, аналізувати, синтезувати потік сучасної інформації.
Цю групу рослин ми пропонуємо для вивчення з використанням різних підходів, а саме:
1) Активного впровадження незвичайних, нетрадиційних ситуацій на уроці.
2) Створення на уроці моментів активного включення учнів в процес пізнання природи, закономірностей її розвитку.
3) Використання в більшій мірі матеріалу, що доводять втілення біологічної теорії в практику.
Мета даної роботи: систематизувати відомості про відділі Мохоподібні, їх різноманітність, значення та принципи охорони.
Об'єктом дослідження виступають мохоподібні та їх поширення на території України.
Предмет – використання мохоподібних людиною .
Методи дослідження: аналітичний огляд літератури.
Для досягнення мети і доказу або заперечення гіпотези нами були поставлені такі завдання:
1. Провести аналіз відділу Мохоподібні
2. Узагальнити відомості про біологічні дослідження Мохоподібних.
3. Проаналізувати поширення окремих видів відділу.
4. З’ясувати значення мохоподібних для людини.
Розділ 1. Загальна характеристика мохоподібних та їх різноманітність на Україні 1.1 Дослідження Мохоподібних на Україні
Біологічні дослідження на території України беруть початок, в основному в середині XIX ст. вони розвивались повільно, інколи залишались незакінченими.
В них приймали участь і небріологи, які займались вивченням мохоподібних поступово, переходячи потім до інших груп рослин чи до інших питань. Роботи по біології проводились майже виключно в університетах - Харківському (Т.А. Алексенко), Київському (А. Покровський і інші), Новоросійському (А.А.Сапегин), а також в Львові [4]. Результати вивчення бріофлори України висвітлені в малочисленних роботах і опубліковані переважно в виданнях Київського, Харківського і Новоросійського товариства природовипробувачів, в "Известиях" Петербурзького ботсаду і інших джерелах. Відомості про мохи представлені в них в вигляді окремих списків чи разом з списками інших груп рослин. Ці дослідження торкнулися окремих місцевостей України [2].
Деякі данні про мохи районів Кам'янець-Подільського і Радомишля приводить Г.Бельке в 1858, 1866 рр., І.Плутенко опублікував в 1871р. матеріали до флори мохів і лишайників Полтавської губернії, які містять список із 36 видів мохів. В.В.Совінскім в "Материалах для флорьі водорослей и отчасти мхов некоторьгх уездов Киевской и Полтавской губернии" (1875) описано 17 видів мохів. АДокровський вперше для околиць Києва вказує 137 видів мохів (1892 р.). В двох роботах М.А.Алексенко представлені списки мохів для північної частини Харківської (1897) і для Чернігівської (1898-1899) губернії [2]. На початку XX ст. К.Шафнагелем згадується ряд видів мохів із району Житомира і районів Кам'янець-Подільської губернії (1908). Відомості до бріофлори південно-сходу України приводить Н.А.Мосолов в вигляді списку, який містить 3- види печіночників і 27 видів мохів. І.Ширяєв в описах сфагнових боліт в околицях Харкова (1909) дає перші відомості про сфагнових мохів району, потім - при описі гіпнових і сфагнових боліт в Лебединському уїзді (1910 р.) А.Матушевський повідомляє про мохи Шепетівських лісів (1913) р.) Н.Н.Чікілевський розглядає екологію моху Polytrichum commune на пісках в південно-західній частині Чернігівщини (1909). В.В.Дотуровський в 1916 р. для поліської частини Волинської губернії приведено 111 видів мохоподібних.
В ряді публікацій представлені матеріали по флорі мохоподібних Криму. За даними Н.Зеленського, відомо було 38 видів мохоподібних, в тому числі 6 видів печінкових мохів: 13 видів були вказані вперше (1896) Е.В.Цикендрат приводить 1 вид печіночників і 13 видів мохів (1900);
О.А. і Б.А.Федченко - 18 видів мохоподібних (1905). До того часу для Криму були відомі 45 видів. Особливий інтерес представляє робота А.А.Сапегіна "Мхи горного Крьіма" (1910) написана по матеріалах зборів автора і колекціям А.Н.Криштофовича, Ф.М.Каменського і інших. В ній наведено 195 видів мохів, описано 4 нових види [4].
В роботі вперше в бріології виражено географічне направлення і закладені основи наукової бріогеографії.
Бріологічні дослідження в Карпатах були початі також з середини XIX ст. і проводились в основному польськими ботаніками, а також угорськими і чехословацькими вченими. Перші відомості про мох регіону зустрічаються у роботах професора Львівського університету Я. Лабажевського. По його матеріалах опубліковано ряд робіт. Значно поповнюються відомості з другої половини 19 ст. В 1860 р. з'являється повідомлення угорського ботаніка Ф. Гашлинського про два види печіночників: Riccia fluitans і Ricciocarpus natars із околиць Ужгорода.Е. Гюккель в роботі 1868 р. приводить список видів мохоподібних в основному для Цівнічно-Західного Прикарпаття.
В роботі Е.Волощака, присвяченій флорі Покутських Карпат (1888), є і список мохоподібних А.Гееба вказує декілька видів для східних Карпат, два із них вперше для регіону. Ф.Ліліенфельдівна в 1911р. приводить повідомлення про 25 видів печіночників із району Косова і 20 видів в основному з хр. Черногора [3].
Бріологічні дослідження на території України в XX ст. продовжились в таких напрямках:
1) інвентаризація гербарних фондів;
2) опис рідкісних і зникаючих видів та їх охорона;
3) дослідження екологічних особливостей мохоподібних: ролі абіотичних факторів у житті мохів; адаптації до них; антропогенного впливу на мохоподібні;
4) дослідження мохоподібних заповідних об'єктів України;
5) мохоподібні як індикатори та монітори забруднення середовища. [12]
1.2 Умови зростання та географічне походження мохоподібнихПредставники мохоподібних були знайдені на всіх континентах і зростають вони майже всюди, хоча в різних географічних областях різняться систематичним складом та кількістю [1].
Кріофіти, які живуть у водоймі, поділяються на дві великі групи: вільно плаваючі на поверхні та повністю занурені. Перших порівняно небагато. Це головним чином не чіпкові. Других набагато більш - десятки видів.
Говорячи про водні мохоподібні, необхідно зазначити, що всі вони зростають лише у прісних водоймах. В морській воді вони майже відсутні [11].
Крім водних є ще такі мохоподібні, які ведуть напівводяний спосіб життя. До них належать багато болотних видів, які зростають на берегах річок, на камінні в їх руслах. Коливання рівня води призводять до того, що такі види то занурюються у воду, то повністю на камінні.
Мохоподібні, які зростають на камінні мають дві форми росту - маленькі кустики з прямих пагін та гілок і пагінів з перпендикулярними гілками.
Це не паразит, він використовує гумус, який накопичується в тріщинах кори дерев, вологу, яка збігає по стовбуру під час дощу.
Два фактори обмежують розповсюдження епіфітів - температура і вологість. Епіфіт, який знаходиться на стовбурі дерева в декількох метрів від землі, як правило, не може бути схований на зиму шаром снігу, який захищає рослини від холоду. Це різко зменшує можливість зростання епіфітів в холодних областях. Епіфіти майже всю необхідну їм вологу отримують із атмосфери, а не з субстрату, який майже не містить вологи. Тому водозабезпечення для них по ступеню вологості стоїть на першому місці. Ще один субстрат, на якому знаходяться мохоподібні - листки. Звичайно, листки завжди зелених рослин. Більшість зростаючих на листях мохоподібних - печіночні мхи, їх називають епіфіпами [3,11]
Але найбільш цікавим субстратом із всіх, що використовують кріофіти, є раковини молюсків, цей вид - фісіденса, зростає ще на іншому субстраті, нерухомому - прісноводні губки.
Ще один приклад субстрату, на якому зростають представники родини Splachnaceae. Представники його ростуть на субстратах тваринного походження. Це експерименти самих різних тварин, як диких, так і домашніх. Всі ці субстрати в умовах теплого або жаркого клімату дуже швидко розкладаються.
Крім того, кріофіти можуть жити ще в дуплах дерев та кустарників, на дерев'яних дахах домів та інше.
Але одна група мхів (їх близько 20 видів) може бути названа безсубстратною. Це ті виді, які можуть відриватися від субстрату і вільно переміщуватись вітром. Ці види подушкоподібні формою. Вони головним чином зустрічаються на відкритих місцях. Відірвавшись від субстрату, ця подушка, яка здатна міняти орієнтацію в просторі, отримує можливість рости во всі боки і з часом стає кулястою. Тому безсубстратні мохи частіше називають мохоподібні кульки. Але ці мохоподібні кульки живі. Всередині нього є невеликий камінець, разом з яким він колись відірвався від субстрату. Звичайно розміри мохоподібних кульок 6-9 см в діаметрі. Максимальний розмір 15 см.
Географічне розповсюдження мохоподібних має ті ж самі закономірності, що і у інших вищих рослин. Для них характерні великі ареали. В теоретичному плані можна запропонувати 3 шляхи створення диз'юнкцій:
1. Види в кожному ізольованому районі виникають знову, незалежно один від одного.
2. Ізольованість створюється розповсюдженням зародків мохоподібних (головним чином спор) за допомогою повітряних течій.
3. Розділений на серію ізольованих частин ареалу, представляє собою залишок колись єдиного, цілісного ареалу.
Перший шлях: багаторазові виникнення виду.
Виходячи з того, що на сьогоднішній день людство знає об еволюції та механізмах видоутворення, цей шлях можна відразу відкинути.
Другий шлях: розповсюдження зародків мохоподібних. Необхідно розглянути два варіанти, коли вид зростає на суходолі і вид зростає у водоймі. В обох випадках розповсюдження буде відбуватись в напрямку діючих вітрів. Зародки можуть бути підняти на велику висоту віднесені далеко на ділянку, яка не контактує з цим ареалом. Але якщо розглянути розповсюдження зародків не як одноразовий акт, а як процес, який відбувається постійно впродовж усього життя виду, то зрозуміло, що таке розповсюдження не може привести до формування ізольованих районів ареалів.
Третій шлях, який розглядає розірвані ареали як залишки колись єдиного ареалу. Вид колись займав велику площину, але згодом під впливом несприятливих умов на великій частині її - вимер, залишившись на двох або декількох ділянках, де умови змінились найменше. Якщо ці ділянки далеко одна від одної, то виникає розірваний, диз’юнктивний ареал [5].
Таким чином, можна зробити висновок, що головний шлях утворення розірваних ареалів - це подрібнення колись єдиного ареалу, інші шляхи формування таких ареалів мають лише другорядне значення.
Розділ 2. Характеристика відділу Мохоподібні 2.1 Особливості відділу Мохоподібні
Мохоподібні - невеликі наземні, рідше водяні рослини, досить простої (для вищих рослин) будови, з чітко виявленою зміною поколінь або ядерних фаз. На відміну від решти вищих рослин з домінуванням у циклі розвитку гаплоїдного, або статевого покоління (гаметофіта) над диплоїдним, або нестатевим (спорофітом). Гаметофіт являє собою слань (талом) або більш-менш вертикальне стебельце - каулідій з дрібними листками - філідіями і ризоїдами, але без справжніх коренів. Провідна система мохоподібних досить простої будови: без судин і навіть без трахеїд. На гаметофіті розвиваються статеві органи - чоловічі (антеридії) і жіночі (архегонії). Запліднення відбувається у вологому середовищі за допомогою рухливих сперматозоїдів [23].
Спорофіт (спорогон) дуже редукований. Він складається із спороносної коробочки і ніжки з гаусторією, якою присмоктується до гаметофіта, і веде напівпаразитичний спосіб життя. У коробочці з клітин археспорія після редукційного поділу утворюються спори, якими мохоподібні розмножуються. Із спори розвивається статеве покоління, або гаметофіт.
В Україні - близько 580 видів. На Поліссі зростають 368 видів мохів, в Лісостепу знайдено їх майже вдвічі менше, а в Степу й ще менше, лише Карпати можуть зрівнятися в цьому з Поліссям. [19]
Відділ Мохоподібні, або Бріофіти (Bryophyta). Клас: Антоцеротовидні, або Антоцеротопсиди. (Anthocerotopsia) Порядок: Антоцеротові (Anthocerotales) Родина Антоцеротові (Anthocerotaceae)
Вид: Антоцерос крапчастий (Anthoceros punctatus) Вид: Феоцерос гладенький (Phaeoceros Iaevis) Клас: Печіночники, або Маршанціопсиди (Marchantiopsida) Підклас: Маршанцієві, або Маршанціїди (Marchantiidae) Порядок: Маршанцієві (Marchantiales) Родина: Річчієві (Ricciaceae)
Вид: Річчія водяна (Riccia fluitans) Родина: Маршанцієві (Marchantiaceae)
Вид: Маршанція поліморфна (Marchantia polymorpha) Підклас: Юнгерманієві, або Юнгерманіїди (Jungermanniidae) Порядок: Метугерманієві (Metzgeriales) Родина: Пелієві (РеШасеае)
Вид: Пелія налисткова (Pellia epiphylla) Порядок: Юнгерманієві (Jungermanniales) Родина: Птилідієві (Ptilidiaceae)Вид: Птилідій красивий (Ptilidium pulcherrimum) Родина: Радулові (Radulaceae)
Вид: Радула сплющена (Radula complanata) Родина: Фруланієві (Frullaniaceae)
Вид: Фруланія розширена (Frullania dilatata) Клас: Листкостеблові мохи, або Мохи, або Бріопсиди (Bryopsida) Підклас: Сфагнові, або Сфагніди (Sphagnidae) Порядок: Сфагнові мохи (Sphagnales) Родина: Сфагнові (Sphagnaceae)
Вид: Сфагнум болотній (Sphagnum palaustre) Підклас: Андреєві, або Андреїди (Andreaeidae) Порядок: Андреєві мохи (Andreaeales) Родина: Андреєві (Andreaeaceae)
Вид: Андрея скельна (Andrea rupestris) Підклас: Брієві, або Бріїди (Bryidae) Порядок: Політрихові (Polytrichales) Родина: Політрихові (Polytrichaceae)
Вид: Політрихум, або Зозулин льон звичайний (Ро commune)
Вид: Зозулин льон ялівцевий (P. juniperium) Вид: Зозулин льон волосистий (P. piliferum) Порядок: Дикранові (Dicranales) Родина: Дикранові (Dicranaceae)
Вид: Дикранум зморшкуватий (Dicranum rugosum) Вид: Дикранум метлевидний (D. scoparium) Порядок: Фунарієві (Funariales) Родина: Фунарієві (Funariaceae)
Вид: Фунарія вологомірна (Funaria hygrometrica) Порядок: Брієві (Bryales) Родина: Брієві (Вгуасеае)Рід: Бріум (Bryum) Родина: Мнієві (Мпіасеае) Рід: Мніум (Мпішп) Родина: Аулакомнієві (Aulacomniaceae)
Вид: Аулакомніум болотний (Aulacomnium palustre) Порядок: Гіпнобрієві (Hypnobryales)
Родина: Амблістегієві (Amblystegiaceae)
Вид: Дрепанокладус викривлений (Drepanocladus aduncus) Родина: Гіпнові (Нурпасеае) Рід: Гіпнум (Hypnum)
Вид: Птіліум страусове перо (Ptilium crista-castrensis) Родина: Гілокомієві (Hylocomiaceae)
Вид: Гілокоміум блискучий Hylocomium splendens) [23]
2.2 Особливості основних класів відділу МохоподібніКлас Антоцеротовидні, або Антоцеротопсиди (Anthocerotopsida)
Сланеві рослини мають просто побудований пластинчастий талом, дихотомічно розгалужений (рис. 1). Клітини талома майже всі однакові, тобто не диференційовані на окремі тканини і органи. Вони містять по одному пластинчастому хлоропласту з піреноїдом. Гаметофіт двостатевий, з антеридіями і архегоніями, зануреними в тіло талома; ризоїди прості. Спорофіт складається з довгого стручкоподібного спорогона, що розкривається двома поздовжніми щілинами, і бульбоподібного присоска. Над присоском міститься на меристематичної тканини, клітини якої, постійно поділяючись, зумовлюють ріст спорогона в довжину. Всередині спорогона є довга неплідна колонка, оточена спорогенною тканиною, з клітин якої утворюються спори з елатерами. Спори довгий час лишаються в тетрадах.
В епідермісі спорогона є типові продихи з двома замикаючими клітинами.
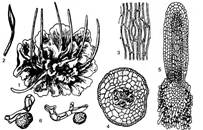
Рис. 1. Феоцерос гладенький (Phaeoceros laevis): 1 — загальний вигляд; 2 — розкритий спорогон; 3 — продих; 4,5 — поперечний і поздовжній розріз спорогона; 6 — спори з елатерами
Клас включає один порядок — Антоцеротові (Anthocerotales), який має одну родину — Антоцеротові (Anthocerotaceae), до складу якої входять п'ять родів і понад 300 видів, поширених майже по всій земній кулі, особливо в тропіках. У флорі України є лише два види: антоцерос крапчастий (Anthoceros punctatus) та феоцерос гладенький (Phaeoceros laevis). Вони ростуть на вогких глинястих ґрунтах, у канавах, по схилах і біля доріг. Антоцеротові мають ряд спільних з водоростями рис будови. Це сланева будова тіла, пластинчасті хлоропласти з піреноїдами. Однак відрізняються від них багатоклітинними гаметангіями, властивими лише для вищих рослин. На думку деяких ботаніків поряд з родиною Антоцеротові наводиться родина Нототиласові (Notothylaceae).
Клас Печіночники, або Маршанціопсиди (Marchantiopsida, або Hepaticopsida )
Сланеві або листкостеблові рослини, в яких гаметофіт майже зйнжди має дорзовентральну будову, верхня (спинна) і нижня (черевна) поверхні побудовані по-різному. Ризоїди прості, одноклітинні, нерозгалужені. Листки (у листкостеблових форм), різноманітні за формою, розміщені на стеблі в два або три ряди, без середньої жилки, здебільшого одношарові. На поверхні слані або на стебельці розвиваються антеридії і архегонії на одній або на різних рослинах; вони в одних випадках заглиблені в тканину талома, в інших — міситься на його поверхні чи на стебельці або підносяться над таломом на особливих підставках. У коробочці спорогона, крім спор, часто утворюються ще особливі пружинки (елатери), які сприяють розсіюванню спор. Коробочка розтріскується здебільшого поздовжніми щілинами на чотири і більше стулок. Стадія протонеми під час проростання спор слабко виражена.
Поширені печіночники майже по всій земній кулі, від тропіків до полярних областей. Найбільша різноманітність їх спостерігається в регіонах з вологотропічним кліматом. Тут у дощових тропічних лісах вони ростуть здебільшого як епіфіти на стовбурах, гілках і листках дерев або на вологих скелях, камінні. В областях з помірним і холодним кліматом печіночники є здебільшого наземними рослинами, що ростуть переважно на сирих і пологих грунтах, у тінистих лісах, рідше в сухих місцях; епіфітів небагато.
У викопному стані печіночники достовірно відомі, починаючи а верхнього карбону. В сучасній флорі світу їх налічується близько 8500 видів, що входять до складу більш як 235 родів і п'яти-семи порядків. У флорі України, за даними Д.К. Зерова, є 165 видів печіночників. Д.К. Зеров запропонував поділити клас Печіночники на два підкласи: Маршанцієві печіночники (Marchantiidae) і Юнгерманієві печіночники (Jungermannidae).

Рис. 2. Печіночники: 1 — річчіокарп {Ricciocarpus natans); 2 — річчія (Riccia fluitans)

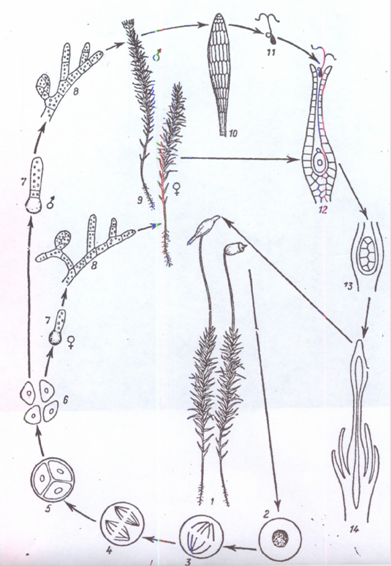
Рис. 3. Цикл розвитку маршанції {Marchantia polymorpha): 1,2 — фрагмент талома з чоловічими і жіночими підставками; 3 — чоловіча підставка в розрізі з антеридіями; 4 — антеридій і сперматозоїди; 5, 6 — жіноча підставка в розрізі з архегоніями, опорогонами і перихецієм; 7 — закритий спорогон; 8 — розкритий спорогон; 9 — опори з елатерами; 10 — проростаюча спора
Підклас Юнгерманієві, або Юнгерманіїди (Jungermanniidae)
Сланеві або листкостеблові рослини досить простої організації, здебільшого з дорзовентральним (рідше радіально-симетричним) типом будови гаметофіта і слабкою (або майже відсутньою) внутрішньою диференціацією тканин. Гаметофіт двостатевий або одностатевий, сланкий, дихотомічно розгалужений (таломного типу) або листкостебловий, з трирядним або дворядним листкорозміщенням. Антеридії сланевих форм виникають на дорзальній поверхні талома, а в листкостеблових — у пазухах листків-філідіїв. Особливих підставок вони не мають. Сперматозоїди численні, дводжгутикові. Архегонії також містяться або безпосередньо на таломах, або на стебельцях чи гілочках, причому розміщені вони або на верхівці стебла, або за точкою росту. Підставок також немає. У листкостеблових і в деяких таломних форм архегонії мають обгортку з листків, яка називається періантієм.
Спорофіт складається з коробочки, що має здебільшого довгу ніжку з добре вираженим присоском біля її основи. Стінки коробочки продихів не мають і складаються з двох - чотирьох і більше шарів клітин. Розкривається вона здебільшого чотирма стулками "бо внаслідок руйнування верхньої частини; колонки всередині немає. Спори і елатери (пружинки) виникають із спорогенної тканини, що є в коробочці. Елатери мають спіральні потовщення, розміщені здебільшого радіально.
Порядок Метцгерієві (Metzgenales)
Містить сланеві форми, рідше рослини розчленовані на стеблову частину і листковидні лопаті. Архегонії та антеридії в спеціальних заглибинах або оточені обгорткою. Родина Пелієві (РеІІіасеае) в Україні представлена трьома видами, з яких дуже поширеною є пелія налисткова (РеІІіа epiphylla), що зустрічається часто на вогких затінених місцях, в ярах, балках, по берегах річок тощо, утворюючи суцільні дернини темно-зеленого кольору (рис. 4). Найбільше поширена пелія в північних районах України і в Карпатах; загальний її ареал охоплює майже всю північну півкулю. Слань у неї плоска, розгалужена на стьожкоподібні галузки до 1 см завширшки; на кінцях слані є точки росту. Ризоїди на нижній поверхні талома численні, прості. Внутрішня будова його також проста; він складається з тонкостінних клітин паренхіми, в якій ледве відокремлено верхній і нижній епідерміс. Хлорофілові зерна у великій кількості містяться у верхньому епідермісі і прилеглих до нього шарах паренхіми.
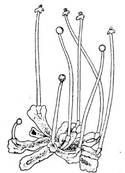
Рис. 4. Пелія налисткова (РеІІіа epiphylla)
Антеридії і архегонії розміщені на верхній поверхні талома. Антеридії численні, розташовані по середній жилці, містяться вони в особливих антеридіальних камерах, кожний окремо. Камери здебільшого червонуватого кольору. Антеридії мають вигляд кулястих тіл з одношаровою стінкою, наповнених дрібними сперма генними клітинами, з яких виникають численні дводжгутикові сперматозоїди. Архегонії містяться трохи нижче точки росту; вони виникають групами і занурені в особливі заглиблення кишенькоподібної форми. Кожний архегоній має черевце і шийку; у черевці міститься яйцеклітина і черевцева канальцева клітина, у шийці — шийкові канальцеві клітини. Під час дощу або роси антеридії лопаються, сперматозоїди звільняються, проникають крізь шийку архегонію до черевця, де один з них зливається з яйцеклітиною, запліднюючи її. Зигота починає швидко ділитися, через короткий час виникає спорофіт (спорогон). Він складається з ніжки з присоском і кулястої коробочки. Деякий час (до початку весни) спорогон заглиблений у слань, має відносно коротку ніжку і одягнений видовженим ковпачком, або каліптрою, яка утворилася з розрослих стінок черевця архегонію. Навесні ніжка спорогона швидко росте і дуже видовжується, коробочка прориває ковпачок і високо підноситься над сланню. Стінки коробочки згодом тріскаються, вона розкривається чотирма стулками, і з неї висипаються спори. Вони одноклітинні, з двома оболонками, з яких внутрішня (ендоспорій) тонша, а зовнішня (екзоспорій) товстіша. Потрапивши в сприятливі умови, спора починає проростати, даючи початок новому талому (гаметофіту) пелії. Спочатку талом має циліндричну форму, але згодом стає плоским, набуваючи вигляду, властивого цій рослині.
Таким чином, цикл розвитку в пелії (як і в інших мохоподібних) полягає в зміні поколінь — статевого і нестатевого. Статеве покоління (гаметофіт) є домінуючим, як і в інших мохів.
Порядок Юнгерманієві (Jungermanniales)
Найчисленніший серед печіночників, містить близько 5000 видів, поширених майже по всій земній кулі. До його складу входять 200 родів і 40 родин. За Т. Реймерсом, їх поділяють на анакрогінні юнгерманієві та акрогінні юнгерманієві. Характерною рисою перших є те, що архегонії, а також спорогон (спорофіти) завжди виникають не на верхівці стебла або в точці росту талома, а трохи нижче їх, тому ріст талома або стебла не припиняється. У акрогінних юнгерманієвих, до яких належать тільки листкостеблові форми, архегонії і спорогони завжди верхівкові, тобто виникають і розвиваються завжди на верхівці стебла або гілочок. У системі печіночників Д.К. Зерова і багатьох Інших вчених немає поділу юнгерманієвих на акрогінні та анакрогінні.
Родина Птилідієві (Ptilidiaceae)
Охоплює рослини з перисторозгалуженими стеблами. Наземні, або епіфіти з дво — чотирироздільними листками, по краю довгозубчастими або війчастими і подібними до листків амфігастріями. Звичайним в Україні є представник оліготипного роду птилідій красивий, поширений на Поліссі і в Карпатах (рис. 5). Зустрічається на корі дерев, пеньках, стовбурах, часом на скелях.
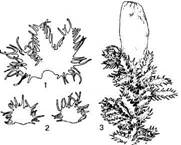
Рис. 5. Птилідій красивий (Ptilidium pulcherri-тит): 1 — листок, 2 — амфігастрії, 3 — верхня частина рослини
Родина Радулові (Radulaceae) Представлена єдиним родом радула (Radula) з численними, переважно тропічними видами (250). Рослини утворюють плоскі, здебільшого жовто-зелені дернинки. Стебло перистогіллясте, з дволопатевими листками. Верхня лопать листка велика, округла, нижня — дрібна ромбічна або трапецієподібна, притиснута до верхньої.

Рис. 6. Радула сплющена {Radula complanata); a — на верхівці однієї з гілочок видно періантій
Радула сплющена (Radula complanata) як епіфіт на корі багатьох листяних дерев і на скелях зустрічається майже в усіх районах України від Гірського Криму до Карпат і Полісся. її загальний ареал охоплює майже всю північну півкулю. Тіло окремої рослини (гаметофіта) складається з тонкого стебельця (до 3-4 см завдовжки), густо вкритого двома рядами широких округлих листків, які тісно прилягають один до одного (рис. 6). Листки плоскі або опуклі, з рівним кілем; кожний листок складається з двох лопатей: верхньої (більшої, широкої і округлої), і нижньої (майже в 4 рази меншої за верхню, неправильної ромбічної або прямокутної форми, притиснутої до верхньої). Від нижньої лопаті листка (від її основи) відходять пучки ризоїдів, якими рослина прикріплюється до субстрату. Радула — дводомна рослина. Архегонії звичайної будови, виникають групами на кінці стебла або гілочок. Спорогон верхівковий, складається з яйцеподібної коробочки і короткої ніжки з присоском; присосок здутий і глибоко проникає у верхівку стебла.

Рис. 7. Фруланія розширена (Frullania dilatata): 1 — частина рослини (вигляд з нижнього боку); 2 — верхівка стебла з періантіем і спорогоном
Родина Фруланієві (Frullaniaceae)
Містить види з дрібними, притиснутими до субстрату дернинками, перисторозгалуженими. Листки дволопатеві: верхня лопать овальна, нижня — менша, шоломовидна, ковпачковидна або утворена-ланцетним листочком. З трьох родів найбільший — фруланія (Frullania), який включає понад 700 видів, переважно тропічних. Одним із звичайних в Україні печіночників є ф. розширена (F. dilatata) (рис. 7). Епіфіт на корі дерев, росте і на скелях. Зустрічається часто — від Полісся і Карпат до Південного берега Криму.
Клас Листкостеблові Мохи, або Мохи, або Бріопсиди (Bryopsida, або Musci)
Тіло (гаметофіт) цих мохів завжди розчленоване на стебло і листки, але з відносно простою внутрішньою диференціацією тканин. Стебло — каулідій, має радіальну будову (рис. 8). Воно здебільшого округле, рідше трикутне, сплющене або багатокутне. Листки-філідії сидячі, розміщені на стеблі тісною спіраллю. Антеридії і архегонії утворюються здебільшого групами на верхівці стебла або на кінцях гілочок. Спорогон (спорофіт) має стопу (гаусторію), ніжку і коробочку (спорангій), здебільшого з кришечкою. Коробочка завжди має всередині особливу колонку. Спори без елатер.
Листкостеблові мохи поширені як у тропіках, так і в позатропічних регіонах з помірним і холодним кліматом. Вони поширюються далеко на північ і високо в гори. Нерідко мохи суцільним килимом укривають великі площі в лісах і на болотах. Деякі види ростуть на стовбурах і гілках дерев, на пнях, скелях тощо. У прісних водоймах оселяються рідко, а в морях, солоних озерах їх немає зовсім.
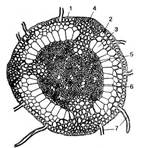
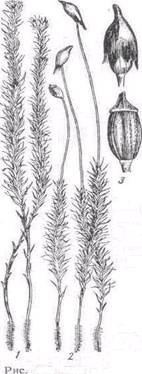
Рис. 8. Поперечний розріз нижньої частини стебла політрихума (Pofytrichum): 1 — епідерміс; 2 — паренхіма; 3 — ендодерма; 4 — гіподермальний тяж; 5 — центральний циліндр; 6 — перицикл; 7 — ризоїд
Листкостеблових мохів відомо понад 14 000 видів, що входять до складу 700 родів, які об'єднуються в три підкласи: Сфагнові, або Сфагніди, Андреєві, або Андреїди, Брієві, або Бріїди.
Підклас Сфагнові, або Сфагніди (Sphagnidae)
Порядок Сфагнові мохи (Sphagnales) характеризується рядом морфологічних, анатомічних і біологічних ознак, якими вони помітно відрізняються від інших мохоподібних, зокрема від зелених мохів. Із спори сфагнового моху виростає спочатку коротка нитчаста протонема, яка незабаром розвивається в пластинчасту (таломну) одношарову протонему з багатоклітинними ризоїдами. На таломній протонемі виникають бруньки, з яких розвиваються стебла (рис. 9).
Стебла сфагнових мохів галузисті, невисокі, без ризоїдів, ростуть увесь час верхівкою, а знизу відмирають, накопичуючи торф. У верхній частині стебла короткі гілочки зібрані в щільну верхівкову голівку. Нижче по стеблу містяться довші гілочки, які розташовані пучками (по кілька штук), більш-менш горизонтально, поверхами. Найнижчі гілочки звісають вниз уздовж стебла.
Внутрішня будова стебла сфагнуму відносно проста. В центрі його міститься серцевина, що складається з досить великих тонкостінних паренхімних клітин. її оточує кілька шарів вузеньких видовжених прозенхімних клітин з потовщеними стінками, здебільшого забарвленими в бурий або червоний колір. Це так званий деревний циліндр — склеродерма. Зовні від циліндра розташований багатошаровий епідерміс, який у сфагнів часто називається ще водоносною корою — гіалодермою. Він складений з досить великих прозорих і порожніх (тобто мертвих) клітин без протопластів. У стінках цих клітин є отвори (пори), а в деяких видів стінки ще мають характерні спіральні та кільчасті потовщення. Клітини епідермісу завжди виповнені водою і розчиненими в ній поживними речовинами. Вони фактично виконують функцію провідної тканини. Характерно, що у водяних сфагнів епідерміс менш розвинутий, ніж у наземних.

Рис. 9. Сфагнові мохи (Sphagnum): 1 — протонема з молодою рослиною; 2 — верхня частина моху зі спорогоном; 3 — архегоній; 4 — гілочка з антеридіями; 5 — антеридій; 6 — спорогон у розрізі; 7 — спорогон збоку
Листки сфагнових мохів складаються з одного шару клітин і на відміну від зелених мохів не мають середньої жилки. Проте клітини сформованих листків моху неоднакові; спостерігається явище так званого диморфізму клітин. Одні з них живі, вузькі, довгі, ніби червоподібні, виповнені хлорофіловими зернами; це асимілюючі хлорофілоносні клітини. Вони затиснуті між широкими більш або менш ромбоподібними клітинами, які позбавлені хлоропластів і є мертвими; це водоносні клітини. Оболонки їх здебільшого мають спіральні та кільчасті потовщення і продірявлені порами, кількість яких у різних видів мохів різна. Оболонки водоносних клітин листка і стебла здатні вбирати воду і розчинені в ній поживні речовини. Ця властивість, а також наявність в оболонках пор забезпечує сфагнуму швидке всмоктуванню води і накопичення її. Встановлено, що сфагнуми можуть вбирати і накопичувати води в 20-30 разів більше за свою масу.
Органи статевого розмноження сфагнових мохів (антеридії і архегонії) містяться на одній рослині (двостатевий гаметофіт) або на різних рослинах (одностатевий гаметофіт). Вони виникають звичайно на коротких бічних гілочках, розміщених біля самої верхівки стебла. Однак антеридії та архегонії ніколи не утворюються на одній гілочці. Гілочки, на яких містяться антеридії, трохи товстіші за вегетативні; листки їх інтенсивно забарвлені у жовтуватий або буруватий колір.
Спорофіт, що виникає із заплідненої яйцеклітини, складається з майже сидячої кулястої коробочки, що тримається на товстій гаусторії. Після запліднення гілочка, на якій міститься архегоній, витягується і виносить на собі коробочку. Отже, коробочка має несправжню ніжку, яка називається псевдоподієм. Усередині коробочки є колонка, яка не доходить до її верхівки. Зверху над колонкою міститься куполоподібний споровий мішок, в якому утворюється багато спор округло-тетраедричної форми. В епідермісі коробочки (до повного її достигання) є хлоропласти і недорозвинуті (нефункціонуючі) продихи із замикаючими клітинами, без хлорофілу. Зверху коробочки є кришечка, а також кільце; перистом відсутній.
У складі порядку сфагнових мохів є тільки одна родина — Сфагнові (Sphagnaceae) з одним родом сфагнум (Sphagnum), що налічує близько 350 видів, поширених майже в усіх частинах світу, особливо в північній півкулі. Ростуть сфагнові мохи переважно на так званих верхових, або сфагнових, болотах, у заболочених лісах, переважно хвойних, а також високо в горах. Оселяючись і поширюючись у лісах, вони разом із зеленими мохами сприяють заболоченню їх, особливо в тайзі; заболочування тундри також відбувається за їхньої участі. Сфагнові мохи найбільш поширені в середній і північній частинах лісової зони, в лісотундрі і тундрі, тайзі Сибіру і Далекого Сходу, на Кавказі. В Україні (29 видів) вони поширені на Поліссі і в Карпатах, де ростуть у хвойних лісах, на торфових (верхових) болотах і в субальпійському поясі серед скель та на вогких гірських луках. По долинах річок, особливо по болотах борових терас, сфагнові мохи заходять далеко на південь України у степову зону. У заболочених хвойних (соснових) лісах Полісся поширені такі види, як сфагнум дібровний (Sphagnum nemoreum), с. компактний (S. compactum). На мохових сфагнових болотах ростуть с загострений (S. cuspidatum), с. обманливий (S. fallax), с. бурий (S. fuscum) тощо. На низинних болотах у вільшняках трапляються с. притуплений (S. obtusum), с. бахромчастий (S. fimbriatum), с. болотний (S. palustre), c. центральний (S. centrale) тощо.
У районах поширення сфагнових боліт ці мохи відіграють важливу роль в утворенні ландшафту і мають велике практичне значення. Сфагнові торфовища, особливо верхових боліт, мало мінералізовані (вміст золи — 1-6%), і торф висококалорійний. Брикетований торф з цих боліт широко використовується як паливо в побуті і промисловості. На торфовому паливі працюють теплові електростанції. Торф використовують також як будівельний матеріал та як хімічну сировину. Висушений сфагновий мох використовують у сільському господарстві як підстилку для тварин.
Підклас Андреєві, або Андреїди (Andreaeidae)
Андреєві мохи становлять невелику групу рослин, яка займає ніби проміжне місце між сфагновими і зеленими мохами. У порядок Андреєві мохи (Andreaeales) входить родина Андреєві мохи (Andreaeaceae) з двома родами. З них один — неуролома (Neuroloma) — монотипний, поширений на о. Вогняна Земля (Південна Америка) і в Антарктиці, другий — андрея (Andreaea) налічує понад 100 видів, які властиві для Арктики і гірських територій середніх широт обох півкуль.
Види роду Andreaea утворюють на скелях і каменях невеликі бурі або темно-бурі щільні дернинки. їхні стебла вилчасто-галузисті, без центрального тяжа, густо вкриті вузькими листками. Стебла прикріплені до субстрату ризоїдами, звичайно товстими, циліндричними або пластинчастими. Листки одношарові, з жилкою або без неї. Антеридії та архегонії містяться на верхівках гілочок, будова їх нагадує будову статевих органів зелених мохів.
Спорогон без ніжки, складається з коробочки і стопи. Коробочка видовжено-яйцеподібна, без продихів, розкривається поздовжніми щілинами на чотири-вісім стулок, з'єднаних на верхівці. Всередині спори утворюється багатоклітинна кулька — протонема. Проростання спор відбувається тоді, коли вони висипаються з коробочки. Багатоклітинна протонема після виходу з оболонки спори розростається, утворюючи дуже розгалужені короткі тонкі нитки. Частина цих ниток перетворюється на зелені пластинки, а їхні крайові тонкі кінцеві нитки стають ризоїдами. Прямостоячі нитки дають циліндричні відростки і листковидні пластинки. В цілому протонема андреї — дуже розгалужена багатоклітинна пластинка різноманітної форми. Така протонема допомагає більшості представників підкласу стати піонерами заростання скель і каміння. У флорі України (в Карпатах) на скелях і серед кам'яних брил зустрічаються два види. Більш поширена а. скельна (A. rupestris), що зустрічається в Карпатах (рис. 10). Це невелика рослина, 1-2 см заввишки, яка утворює щільні дернинки бурого або навіть темно-бурого кольору. Зрідка трапляється а. альпійська (А. аіpestris), яка має дернинки до 4 см заввишки. Значення андреєвих мохів у природі і практиці дуже незначне, навіть у районах, де вони досить поширені.
Гаметофіт, а.бо статеве покоління, брієвих мохів складається з протонеми і листкостеблового пагона, або гаметофора. Протонема має здебільшого нитчасту будову, звичайно дуже розгалужена, зелена; розвивається вона зі спори (первинна протонема) або з будь-якої частини гаметофіта (вторинна протонема). На протонемі утворюються особливі бруньки, з яких розвиваються листкостеблові пагони (гаметофори). З появою їх протонема звичайно відмирає; тільки в деяких мохів вона залишається протягом усього життя рослини.

Рис. 10. Андрея скельна (Andreaea rupestris): 1 — загальний вигляд; 2 — протонемаПідклас Брієві, або Бріїди (Bryidae)
Стебла брієвих мохів або не розгалужені зовсім, або галузяться моноподіально, рідше дихотомічно. Вони прямостоячі або лежачі (сланкі), досить простої внутрішньої будови, тобто слабко диференційовані на епідерму, кору і основну тканину. У самому центрі стебла іноді міститься центральний тяж із тонкостінних клітин.
Листок являє собою одношарову пластинку різної форми (округлої, яйцеподібної, лінійної, ланцетної тощо) із серединною жилкою, що має звичайно кілька шарів; іноді її немає. Жилка часом буває така широка, що займає значну частину листка. До стебла листок прикріплюється своєю широкою основою, як правило, впоперек.
Антеридії і архегонії виникають на верхівках стебел або на кінцях бічних відгалужень, на одній або на різних рослинах. У першому випадку гаметофіт двостатевий, у другому — одностатевий. Антеридії та архегонії розміщені звичайно групами і виникають з епідермальних клітин. Антеридії мають форму видовжених міхурців з короткими ніжками. Між ними часто розвиваються нитко- або булавоподібні волоски, так звані парафізи.
Архегонії мають звичайну для мохоподібних будову, але на відміну від печіночників у них шийки довші, а черевце має не одношарову, а багатошарову стінку. Після запліднення стінка черевця дуже розростається, утворюючи так звану каліптру, яка ніби ковпачком закриває коробочку спорогона. Якщо архегоній міститься на верхівці стебла і спорогон, що виникає з яйцеклітини архегонія після її запліднення, відповідно є верхівковим, то такі мохи називають верхоплідними, або акрокарпними. Якщо архегоній і спорогон містяться на верхівці бічного вкороченого пагона, то такі мохи називають бокоплідними або плеврокарпними. Такий поділ є зручним для визначення зелених мохів: систематичного значення йому тепер майже не надають. .
Спорогон (спорофіт), який розвивається із заплідненої яйцеклітини (запліднення відбувається під час дощу або великої роси), є диплоїдом, тобто ядра його клітин миють подвійну кількість хромосом. Ніжка спорогона буває різної довжини і складається з суцільної тканини. Верхній кінець ніжки здебільшого розширений В апофізу, яка є водночас основою коробочки.
Коробочка округла, овальна або циліндрична, прямостояча, відхилена або звисла. Вона складається зі спороносної частини, або урни, і кришечки. В центрі урни завжди є колонка, що піднімається Від апофізи. Стінки урни здебільшого багатошарові, в їхньому епідермісі часто є продихи, особливо в нижній частині. Кришечка конічна, опукла, плоска тощо. Зверху вона накрита ковпачком, котрий є залишком архегонія. Між кришечкою і краєм урни є пояс клітин з потовщеними стінками, який називається кільцем. При достиганні спорогона кришечка разом із ковпачком відокремлюється від урни і Відпадає, а спори висипаються. Кільце в одних мохів відпадає разом з кришечкою, а в інших залишається на урні. Розсіювання спор із коробочки в більшості зелених мохів регулюється так званим перистомом — одним або двома рядами зубців, розташованих по краю урни. Якщо зубці розміщені в один ряд, перистом називається простим, а якщо в два концентричні ряди, — подвійним.
Потрапивши в сприятливі умови, спора проростає, утворюючи нитчасту протонему. На ній розвиваються особливі бруньки, котрі дають початок листкостебловим рослинам моху. У деяких мохів протонема залишається протягом усього життя. Це так звані протонематичні мохи. Крім розмноження спорами, зелені мохи здатні до вегетативного розмноження за допомогою виводкових бруньок, виводкових тілець, виводкових пагонів тощо.
Брієві мохи — один з найбільших підкласів серед мохоподібних; він налічує понад 13 500 видів, об'єднаних у 657 родів, 80 родин. Поширені ці мохи в усіх частинах світу — від тропіків до холодних полярних регіонів. Особливо велика різноманітність мохів спостерігається в гірських тропічних регіонах з вологим кліматом. У пустелі мохів або зовсім немає, або вони зустрічаються дуже рідко. За сприятливих умов брієві мохи ростуть дернинами, купинами або подушками на грунті, скелях, стовбурах дерев, а деякі навіть у прісних водоймах. У лісах, на луках брієві мохи нерідко утворюють суцільні покриви, іноді на великих площах. У лісових рівнинних районах України, Гірському Криму, Карпатах і Степу налічується понад 500 видів брієвих мохів. У Сибіру брієві мохи досить широко представлені багатьма порядками, родинами і видами, кількість яких ще остаточно не встановлена. Цей найбільший із листкостеблових мохів підклас поділяють останнім часом не менше як на 13 порядків, з яких розглянемо лише п'ять.
Розділ 3. Роль мохоподібних в житті людини та природи та охорона
Мохоподібні - могутні сорбенти. Вони здатні поглинати не лише значну кількість води, а й інші хімічні сполуки. Так, при дослідженні вмісту накопичених елементів у водних мохів, зібраних у різних частинах басейну річки, можна виявити місце залягання покладів корисних копалин.
Сорбентні властивості мохоподібних використовуються у промисловості для очищення стічних вод на гірничодобувних підприємствах, у цьому разі в ролі фільтра. Очищену воду можна використовувати повторно і значно зменшуються втрати цінної сировини.
Мохоподібні як рослини-біоіндикатори використовуються в зв'язку з їх адсорбційними властивостями та надзвичайною чутливістю до забруднення. В дуже забруднених районах мохи відсутні, в менш забруднених - з'являються лише стійкі толерантні види. Бріофіти здатні концентрувати у своєму тілі сполуки різних елементів, причому накопичують їх значно більше, ніж інші групи рослин. За допомогою спектрального аналізу можна швидко встановити ступінь забруднення атмосфери, а проводячи такі дослідження протягом 10 років, можна простежити за змінами в атмосфері не лише певного міста, а й регіону в цілому.
Сорбентні властивості моху дають змогу використовувати їх як теплоізоляційний, пакувальний матеріал.
Використовуються мохи і в медицині. Види родів листяних мохів (брій, мній, філонотіс, політрих) - застосовувалися у вигляді свіжих паст при опіках та вивихах. Особливо широко (майже 40 видів) мохи використовувалися в китайській медицині. Одним з найцінніших "медичних мохів" у деяких країнах вважається зозулин льон звичайний, який є жарознижувальним, дезінфікуючим, сечогінним; здатний розчиняти камені в нирках і жовчному міхурі. Такими видами як фунарія вологомірна, брій сріблястий, клімацій деревоподібний, лікують очі.
Застосовують бріофіти із естетичною метою: для декорування приміщень, створення мініатюрних садів, альпійських гірок.
Все це - приклади позитивного використання бріофітів. Але мохи також негативно впливають на довкілля. Вони сприяють заболочуванню лісів, луків, що знижує господарську цінність останніх. Поселяючись на дахах, вони призводять до руйнування покрівлі. Деякі види мохів спричиняють у людини алергічні реакції [14].
Виявляється, що окремі види мохів, які донедавна нікого, крім фахівців-ботаників не цікавили, тепер потрібні геологам, хімікам та іншим
Використання цієї групи рослин почалось ще у 17ст. На території Англії була виявлена корзина, яка була сплетена із Polytrichum commune. Вік знахідки приблизно 1900років. Вважається, що це самий перший доказ використання мохоподібних в господарстві.[11]Найбільш широко використовують різноманітні види сфагніт. Характеризуй використання сфагнових, неможливо не згадати торф [1].
Утворюється він на болотах і уявляє собою спресування, не повністю розклавшихся в умовах ускладненої аерації залишків болотних рослин. Усі рослини, які зростають на болотах є торфоутворюючими. Значення їх в утворенні торфа приблизно теж саме, що і в складенні рослинності болота. Тому визначають три види боліт - низинне, перехідне і верхове. Низинний торф утворюють залишки листополіпних мохів із підкласа брієвих (звичайно їх називають зеленими мохами). Перехідний утворюється із залишків сфагнових мхів. Верховий — це перш за все залишки сфагнових, а серед них на першому місці стоїть Sphagnum fuscum [13,14].
Використання торфу в народному господарстві дуже різноманітне. Перш за все, це томливо. Але торф - це не лише паливо. Це також добриво, особливо важливо для підзолистого ґрунту. Для добрива торф використовують у чистому вигляді, а також в компостах [14,16].
Торф є також доброю підстилкою для усіх сільськогосподарських тварин (також птахів). По цілому ряду характеристик торф'яна підстилка є набагато краще ніж усі інші. Вона характеризується дуже високою вологостійкістю. Наприклад, маса води, яку може поглянути і утримати солома, перевищує її власну масу десь в 2,4 рази. А торф'яна підстилка може поглинути і утримати води стільки, що її маса перевищує масу торфу від 4 до 10 разів. В окремих випадках від 20 і навіть 30 разів.
Торф'яна підстилка має також властивість поглинати гази у великих кількостях, сильними теплоізоляційними властивостями. Вона має також антисептичні та інсектицидні властивості. Це захищає тварин від хвороб, не дає змоги розвиватися паразитам.
Торф є важливим джерелом, сировиною для отримання ряду цінних речовин. З нього отримують етиловий та метиловий спирти, оцтову кислоту, нітрати, нафталін; торф входить до складу активованого вугілля, продукти його переробки - в склад різних маслин. Переробки торфу кислотами дозволяє отримати з нього речовини, які використовують в якості кормових для сільськогосподарських тварин.
Крім того торф використовують в медицині. З нього після спеціальної обробки готують лікувальні грязі. Використовують торфотерапію при різних захворюваннях [4].
Сфагнові мохи здатні поглинати вологу в 30-35 разів більше ніж їх власна маса. Ця фактична гігроскопічність сфагнових робить їх гарним перев'язочним матеріалом, який в багатьох значеннях краще за вату. Так, сфагнові поглинають вологу приблизно в 3-4 рази більше, ніж вата тієї ж маси, і в 3 рази швидше. Крім того, вони краще утримують вологу і поглинають більш рівномірно. Це зменшує кількість перев'язок однієї і тієї ж рани. Сфагнові пов'язки м'яке та більш прохолодні ніж ватні і тому менш подразнюють хворе місце. Сфагнові мають сильну бактерицидну дію.
Ця дія полягає в зменшенні мікрофлори відкритих ран, особливо гнійних та інфікованих. Тому сфагнові - не лише перев’язочний матеріал, але і лікарський препарат.
Дуже широко використовуються і інші бліофіти в лікарських цілях. Наприклад, Marchantia polymorpha I Conocephalum conicum для лікування захворювань печінки. Раніше була відомою „доктрина знаків". В зовнішній схожості якоїсь рослини бачиться знак, що саме ця рослина повинна бути використана при захворюванні того органа, на який вона схожа. Слань печіночника Conocephalum conicum по формі нагадує печінку людини. Саме ця схожість і є причиною такої назви для рослини - печіночникові.
Маршанція як лікарська рослина у XVIII ст. Вважалась єдиним засобом проти усіх захворювань печінки.
Зозулин льон (Polytrichum commune) використовувався в якості засобу, який покращував ріст волосся і надавав їм кращого кольору.
Можна сказати, що антисептичну активність виявлено майже у всіх видів мохоподібних. Виявлено, що деякі мохоподібні здатні пригнічувати рістдеяких видів мікроорганізмів (іноді більше десяти). Наприклад, Marchantia paleacea, він здатен пригнічувати ріст дев'яти різних мікроорганізмів. Серед мікроорганізмів, ріст яких пригнічується екстрактами із мохоподібних, є золотистий стафілокок, кишечна паличка, сальмонела, кандіда, сарцина жовта, холерний вібріон, стрептокок [8].
Хімічна природа речовини, забезпечуючи антибіотичну активність мохоподібних, не зовсім виявлена. Вважають, що можуть діяти дві групи речовин: органічні кислоти або полі феноли або фенолоподібні речовини. Було виявлено, що маршанція використовувалась не лише в якості засобу проти захворювання печінки, але і як протитуберкульозний засіб. Для лікування захворювань печінки використовувався і печіночник lonocephalum conicum, в якості клітинної речовини - листопагінний мох Pseudoscleropodium purum. Відвир із зозулиного льону використовувався в якості речовини, яка допомагає розчиняти каміння в нирках і в жовчному міхурі, при сильних кровотечах (як кровозупиняючий засіб). В Китаї цей вид взагалі вважався одним із найкращих „медичних" мохів. Він широко використовувався там в якості ліків від температури, а також дезінфікуючого засобу. Індіанці Аляски використовували бокоплідні мохи для лікування опіків, ран.
Використовують мохи для очистки стічних вод гірничих заводів. При проходженні стічних вод через мох він може поглинати іони елементів, які добуваються. У води стануть чистими, таким чином втрата цінної речовини зменшиться [15,16]. Велике значення має використання мохоподібних для індикації умов середи, виявлення ступеня її забруднення. Накопичення в собі різних елементів, мохоподібні не лише показують ступень забруднення атмосфери. Вони також є чудовими індикаторами наявності або відсутності їх в субстраті. Це відкриває широкі можливості використання їх в біогеохімічних дослідах.
Використовують біогеохімічний метод пошуку копалин, геологи Якутії вже виявили декілька районів, які містять багато цінних речовин, в тому числі олова, вольфраму, золота. Ще один шлях використання кріофітів - використання мохів в якості пакувального матеріалу. Живі мохи дуже широко використовуються в різних країнах для транспортування саджанців, живих рослин, деяких використовують для культивування орхідей. Крім того мох використовують для транспортування крихких предметів, фаянсового посуду. Широко використовуються мохи для набиття подушок. З одного боку мохи це - чудовий матеріал, з іншого боку - вважалось, що вони викликають сон. Крім того, мох ще використовують як теплоізоляційні прокладки при будівництві дерев'яних будівель [7].
В Скандинавії мох (зозулин льон) використовувався в якості матраців та ковдр.
Але крім цього мохоподібні приносять і шкоду для людини. Наприклад, розростання мохів на лугах знижує їх хазяйське значення, робить малопридатними, може привести до заболочення. Мохи знищують дерев'яні дахи домів, вони забезпечують накопичення вологи, це призводить до більш швидкого знищення деревини.
Висновок
Мохоподібні — невеликі наземні, рідше водяні рослини, досить простої (для вищих) будови, з чітко виявленою зміною поколінь, або ядерних фаз, і на відміну від решти вищих рослин з домінуванням у циклі розвитку гаплоїдного, або статевого, покоління (гаметофіта) над диплоїдним, або нестатевим (спорофітом). Гаметофіт являє собою слань (талом) або більш-менш вертикальне стебельце — каулідій з дрібними листками-філідіями і ризоїдами, але без справжніх коренів. Провідна система мохоподібних досить простої будови: без судин і навіть без трахеїд. На гаметофіті розвиваються статеві органи — чоловічі (антеридії) і жіночі (архегонії). Запліднення відбувається у вологому середовищі за допомогою рухливих сперматозоїдів. Спорофіт (у мохоподібних він називається спорогоном) дуже редукований. Він складається із спороносної коробочки і ніжки з гаусторією, якою присмоктується до гаметоцЫТа, і веде напівпаразитичний спосіб життя. У коробочці з клітин археспорія після редукційного поділу утворюються спори, якими мохоподібні розмножуються. Із спори розвивається статеве покоління, або гаметофіт.
Усі мохоподібні поділяють на три класи: Антоцеротовидні, або Антоцеротопсиди; Печіночники, або Маршанціопсиди, і Листко-стеблові мохи, або Мохи, або Бріопсиди.
Деякі видів мохів вважаються рідкісними і потребують охорони.
Список використаних джерел
1. Алейко В.И., Дисько И.А., Вьгпаев Г.В., Козловская Н.В. По станицам Красной книги. - Мн: Бел СЭ. 1987. – 248 с.
2. Андрієнко Т.Л., Горностай ВЛ., Графін М.Ф. Зелені скарби Чернігівщини. – Чернігів, 2004. – 84 с.
3. Бачурин А.Ф., Партыка Л.Я. Печёночные мхи Украины. - K.: I979.
4. Бардунов Л.В. Древнейшие на суше. - Сибирское отделение, 1984 – 150 с.
5. Вірменко В.М., Панченко С.М. Мохоподібні // Чорноморський ботанічний журнал. - 2005.-№2. – С. 92-109
6. Возхинския В.Б., Трасе Х.Х., Шляхов Р.Н. Водоросли, лишайники и мохообразные СССр . -М: Мысль, 1978. - 365с.
7. Горностай В.І., Графін М.В., Дадашева Т.Г. Вищі спорові судинні рослини. – Чернігів, 2005. – С. 27, 28,39,68.
8. Ивченко СИ. Занимательно о ботанике. - М.: Молодая гвардия, 1991. – С.4-8.
9. Карпенко Ю.О.При ліс. – Чернігів, 2005 - с. 57.
10. Комарницкий Н.А., Кудряшов Л.В., Уралов А.А. Систематика растений. - М: Просвящение, 1962. - С. 210 -249
11. Остапова В.Г. Лесные болота. - М., 1982 – 127 с.
12. Рейв Н.П., Питер. Современная ботаніка: 1 т. / Под ред. Шверт Р. - М.: Мир, 1990. - С. 30.
13. Слюсарів А.О., Самсонов О. В. Біологія - 3-тє вид. О - К.: Вища шк.. 2001. – 622 с.
14. Тахтаждян А.Л. Мир растений. - М.: 1991. - С. 109
15. Федоров А.А. Жизнь растений. - М.: Просвящение, 1974. - С. 56-97
16. Хржановский В.Г., Исаин В. Н. Приктический курс ботаніки. - М: Высшая школа, 1963. – 299 с.
Похожие работы
... іл 4. Мохоподібні лісових ценозів околиць сіл розсошенці та копили В результаті наших досліджень встановлено, що в лісових масивах околиць сіл Розсошенці та Копили виявлено 19 видів мохоподібних. Вони належать до 8 родин, до 12 родів і 15 видів. Таблиця 1. Видовий склад зібраних мохоподібних РОДИНА Polytrichaceae Dicranaceae Funariaceae Bryaceae Mniaceae Brachytheciасeae ...

... нові відомості про динаміку чисельності видів, темпи відновлення або скорочення ареалів, життєвість тощо. Друге, нинішнє видання Червоної книги підготовлене у двох томах: “Тваринний світ” і “Рослинний світ”. “Червона книга України. Рослинний світ” складається з 5 розділів, що включають статті про 541 вид (підвид, різновидність, форма) рослин і грибів: судинні рослини (439 видів), мохоподібні ...
... 'єктами права лісокористування можуть бути державні, колективні, громадські підприємства, установи, організації, в тому числі релігійні, громадяни України та їх об'єднання, іноземні громадяни та юридичні особи. Об'єктами права лісокористування є лісові ресурси та земельні ділянки лісового фонду. Змістом права лісокористування є сукупність повноважень лісокористувачів або сукупність визначених ...
... ї ваги рекреаційної сфери у зростанні національного доходу країни. 3. РЕСУРСОЗБЕРЕЖЕННЯ ЯК ФАКТОР ПІДВИЩЕННЯ ЕФЕКТИВНОСТІ ВИРОБНИЦТВА Ресурсозбереження — це прогресивний напрям використання природно-ресурсного потенціалу, що забезпечує економію природних ресурсів та зростання виробництва продукції при тій самій кількості використаної сировини, палива, основних і допоміжних матеріалів. ...
0 комментариев