МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РФ
ФГОУ ВПО
ОРЛОВСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ
Кафедра микробиологии и вирусологии
Реферат
«Мутация вирусов, характеристика мутагенов»
Орел – 2008
План
Введение
Мутация у вирусов
Частота мутаций и механизмы их возникновения
Модификации, вызываемые хозяином
Изменчивость вирусов при пассажах
Изменчивость вирусов при пассажах на животных
Изменчивость вирусов при пассажах на куриных эмбрионах
Изменчивость вирусов при пассажах на культуре клеток
Изменчивость вирусов, возникающая в процессе пассажей при пониженных и повышенных температурах
Химические мутагены
Физические мутагены
Заключение
Список литературы
Введение
Повышение сохранности и продуктивности сельскохозяйственных животных невозможно без дальнейшего совершенствования ветеринарного обслуживания животноводства. Среди ветеринарных дисциплин важное место принадлежит вирусологии. Современный ветеринарный врач должен знать не только клинико – патологическую сторону болезни, но и иметь четкое представление о вирусах, их свойствах, методах лабораторной диагностики и особенностях постинфекционного и поствакцинального иммунитета.
Вирусы – облигатные внутриклеточные паразиты животных, растений, насекомых, бактерий, грибов, простейших и других живых существ.
Вирусы изменяют свой свойства как в естественных условиях размножения, так и в эксперименте. В основе наследственного изменение свойств вирусов могут лежать два процесса: 1) мутация, т. е. изменение последовательности нуклеотидов в определенном участке генома вируса, ведущее к фенотипически выраженному изменению свойства; 2) рекомбинация, т. е. обмен генетическим материалом между двумя близкими, но отличающимися по наследственным свойствам вирусами.
Мутация у вирусов
Мутация – изменчивость, связанная с изменением самих генов. Она может иметь прерывистый, скачкообразный характер и приводить к стойким изменением наследственных свойств вирусов. Все мутации вирусов делятся на две группы:
· спонтанные;
· индуцированные;
По протяженности их делят на точечные и аберрационные (изменения, затрагивающие значительный участок генома). Точечные мутации обусловлены заменой одного нуклеотида (для РНК-содержащих вирусов). Такие мутации могут иногда ревертировать с восстановлением исходной структуры генома.
Однако мутационные изменения способны захватывать и более крупные участки молекул нуклеиновых кислот, т. е. несколько нуклеотидов. В этом случае тоже могут происходить выпадения, вставки и перемещения ( транслокация ) целых участков и даже повороты участков на 180° ( так называемые инверсии ), смещения рамки считывания – более крупные перестройки в структуре нуклеиновых кислот, а следовательно, и нарушения генетической информации.
Но не всегда точечные мутации приводят к изменению фенотипа. Имеется ряд причин, в силу которых такие мутации могут не проявляться. Одна из них – вырожденность генетического кода. Код белкового синтеза вырожден, т. е. некоторые аминокислоты могут кодироваться несколькими триплетами (кодонами). Например, аминокислота лейцин может кодироваться шестью триплетами. Вот почему, если в молекуле РНК вследствие каких-то воздействий произошла замена триплета ЦУУ на ЦУЦ, ЦУА на ЦУГ, то в молекуле синтезируемого белка все равно включится аминокислота лейцин. Поэтому ни структура белка, ни его биологические свойства не нарушатся.
Природа пользуется своеобразным языком синонимов и , заменяя один кодон другим, вкладывает в них одно и тоже понятие ( аминокислоту ), сохраняя, таким образом, в синтезируемом белке его естественную структуру и функцию.
Другое дело, когда какая-то аминокислота кодируется всего одним триплетом, например, синтез триптофана кодируется только одним триплетом УГГ и замены, т. е. синонима, не имеется. В этом случае в белок включается какая-нибудь иная аминокислота, что может привести к появлению мутантного признака.
Аберрация у фагов обусловлена делециями (выпадением) различного числа нуклеотидов, от одной пары до последовательности, которая обуславливает одну или несколько функций вируса. Как спонтанные, так и индуцированные мутации делят также на прямые и обратные.
Мутации могут иметь разные последствия. В одних случаях они ведут к изменению фенотипических проявлений в нормальных условиях. Например, увеличивается или уменьшается размер бляшек под агаровым покрытием; увеличивается или ослабляется нейровирулентность для определенного вида животных; вирус становится более чувствительным к действию химиотерапевтического агента и т. п.
В других случаев мутация является летальной, так как вследствие ее нарушается синтез или функция жизненно важного вирусспецифического белка, например вирусной полимеразы.
В некоторых случаях мутации являются условно летальными, так как вирусспецифический белок сохраняет свои функции в определенных для него условиях и теряет эту способность в неразрешающих (непермиссивных) условиях. Типичным примером таких мутаций являются температурно-чувствительные - ts-мутации, при которых вирус теряет способность размножаться при повышенных температурах (39 - 42°С), сохраняя эту способность при обычных температурах выращивания (36 – 37°С).
Морфологические или структурные мутации могут касаться размера вириона, первичной структуры вирусных белков, изменения генов, детерминирующих ранние и поздние вирусспецифические ферменты, обеспечивающие репродукцию вируса.
По своему механизму мутации могут быть тоже разными. В одних случаях происходит делеция, т. е. выпадение одного или нескольких нуклеотидов, в других – происходит встраивание одного или нескольких нуклеотидов, а в некоторых случаях – замена одного нуклеотида другим. [1]
Мутации могут быть прямыми и обратными. Прямые мутации меняют фенотип, а обратные (реверсии) – его восстанавливают. Возможны истинные реверсии, когда обратная мутация происходит вместе первичного повреждения, и псевдореверсии, если мутация происходит в другом участке дефектного гена (интрагенная супрессия мутации) или в другом гене (экстрагенная супрессия мутации). Реверсия не является редким событием, так как ревертанты обычно более приспособлены к данной клеточной системе. Поэтому при получении мутантов с заданными свойствами, например вакцинных штаммов, приходится считаться с возможной их реверсией к дикому типу.
Вирусы отличаются от остальных представителей живого мира не только своими малыми размерами, избирательной способностью размножаться в живых клетках, особенностями строения наследственного вещества, но и значительной изменчивостью. Изменения могут касаться величины, формы, патогенности, антигенной структуры, тканевого тропизма, устойчивости к физико – химическим воздействиям и других свойств вирусов. Значение причин, механизмов и характера изменения имеет большое значение при получении необходимых вакцинных штаммов вирусов, а также для разработки эффективных мер борьбы с вирусными эпизоотиями, в процессе которых, как известно, свойства вирусов могут существенно изменят одной из причин сравнительно высокой способности вирусов изменять свои свойства является то, что наследственное вещество этих микроорганизмов менее защищено от воздействия внешней среды.
Мутация вирусов может возникать в результате химических изменений цистронов или нарушения последовательности их расположения в структуре молекулы вирусной нуклеиновой кислоты.
В зависимости от условий различают естественную изменчивость вирусов, наблюдаемую в обычных условиях размножения, и искусственную, получаемую в процессе многочисленных специальных пассажей или путем воздействия на вирусы особых физических или химических факторов (мутагенов). [2]
В естественных условиях изменчивость проявляется не у всех вирусов одинаково. Наиболее ярко этот признак выражен у вируса гриппа. Значительной изменчивости подвержен вирус ящера. Об этом свидетельствует наличие большого количества вариантов у разных типов этих вирусов, и существенные изменения его антигенных свойств в конце почти каждой эпизоотии.
Частота мутаций и механизмы их возникновения
Мутации бактериофагов изучались очень интенсивно не только с целью генетического анализа, но и для получения информации о свойствах самих фагов. Частота появления тех или иных мутантов в фаговом потомстве варьирует в весьма широких пределах: например, некоторые мутанты образуются с частотой не выше 10 , тогда как другие возникают с частотой 10 и выше. Неблагоприятный эффект высокой частоты мутаций обычно компенсируется действием отбора. Например, мутант фага может быть вытеснен диким типом, который дает больший выход фага.
Высокая частота вспонтанного возникновения обычно характерна для таких мутаций, которые могут происходить во многих сайтах одного локуса. В тех случаях, когда нормальный признак соответствует функциональной форме гена, а мутантный появляется в результате какого-то изменения в любой точке данного локуса, частота прямых мутаций будет выше, чем частота обратных, так как обратные мутаций должны приводить к восстановлению нормального состояния. Иногда ревертанты оказываются на самом деле псевдоревертантами: это происходит либо в результате изменений в каком-либо другом гене (супресорные мутации), либо вследствие изменений в том же гене, которые обусловливают иную, но также активную форму продукта.
У зрелых фаговых частиц частота вспонтанных мутаций очень низка, но их можно идуцировать, воздействуя какими-либо мутагенными факторами, например рентгеновскими или ультрафиолетовыми лучами, азотистой кислотой, гидроксиламином или алкилирующими агентами. Азотистая кислота дезаминирует основания нуклеотидов, а этилметилсульфонат их этилирует. Гидроксиламин превращает щитозин в урацил. При зарожении модифицированными фагами вследствие ошибок, происходящих при репликации химически измененной нуклеиновой кислоты, возникают мутации, и потомство фага, высвобождающееся из одной бактерии, содеожит как нормальные, так и мутантные частицы. Однако, как и следует ожидать при обработке мутагеном фага, содержащего одноцепочечную ДНК, образуется чистый клон мутанта.
Изучение мутационного процесса, происходящего во время размножения фага, имеет большое прямое отношение к анализу развития фага. Сначала рассмотрим спонтанный мутационный процесс. В бактериальной клетке, в которой произошла мутащия фага, образуется как нормальный, так и мутантный фаг. Число мутантных фаговых частиц, содержащихся в популяции фага, выходящей из данной единичной бактериальной клетки, очевидно, определяется характером репродукщии фага, ибо новые гены могут образоваться лишь путем репликации предсуществовавших. Если вероятность данной мутации при каждой репликации одинакова, то число возникших мутантов зависит от механизма репликации. Например, если каждая новая копия гена образуется независимо от остальных, то распределение мутантных копий в потомстве фага из различных инфицированных бактерий будет случайным. Если же, напротив, каждая из образующихся копий будет в свою очередь репродуцироваться, то мутантные копии будут встречаться группами, или клонами, состоящими из мутантных «сибсов».
Для проверки этих предложений были проведены эксперименты. Бактериальные клетки заражали фагом Т2 и подсчитывали бляшки, образуемые мутантными частицами, - r-мутанты (вызывающие быстрый лизис). Затем находили распределение по частоте клонов, содержащих 1, 2, 3, . . .5,… мутантов, и сравнивали найденное распределение с рассчитанным, которое было основано на допущении, что воспроизведение фаговых генов – экспоненциальный процесс, состоящий из последовательных дупликаций, а вероятность мутирования гена при каждой репликации – величина постоянная. Если эти допущения справедливы, то следует ожидать, что частота y , с которой образуются клоны, содержащие
х(=2 ) мутантов, будет следующей:
где m – вероятность мутации, N – конечное число копий гена, а n – число генераций фаговых генов, полученных в данном опыте. Поскольку на практике геномы фага реплицируются несинхронно, удобнее анализировать распределение мутантов, исходя из частоты Yx образования клонов, содержащих x или более мутантов:
Распределение, полученное в эксперименте очень хорошо совпадает с теоретическим, особенно если учитывать влияние, которое оказывает на результаты опыта ограниченная величина каждого выхода фага - сборки зрелых фаговых частиц из фонда реплицирующихся элементов. Из полученных данных следует, что репликация генов фага – экспоненциальный процесс, аналогичный размножению любого организма, которое происходит путем повторных делений. Конечно, эта аналогия носит чисто формальный характер и является просто следствием полу консервативного механизма репликации фаговой ДНК, т. е. результатом образования из каждой двухцепочечной молекулы ДНК двух новых двухцепочечных молекул. [3]
Интересная ситуация была обнаружена при анализе распределения мутантов, образовавшихся в результате обработки фага Т4 мутагеном этилметилсульфонатом. Оказалось, что клоны с 1, 2, 4, 8 мутантами встречались примерно с одинаковой частотой. Известно, что действие этого мутагена на фаговую ДНК состоит в этилировании гуанина. Поэтому полученные данные были интерпретированы в том смысле, что для каждого этилгуанина, входящего в состав ДНК, существует некоторая постоянная вероятность замены на неправильное основание. Следовательно, некая мутация может возникнуть с равной вероятностью в любой генерации фага и все копии, образующиеся после этого события, будут мутантами. Результаты этого опыта показали, что цепи ДНК исходных частиц фага Т4 неоднократно копировались внутри одной зараженной бактериальной клетки.
Модификации, вызываемые хозяином
В дополнение к мутациям бактериофаги подвергаются негенетическим изменениям, в которых основная роль принадлежит клетке-хозяину. Это явление получило название модификаций, вызываемых хозяином (Luria, Human, 1952; Arber, 1965, 1974). Важность этих модификаций для молекулярной биологии состоит в том, что они продемонстрировали способность внутриклеточной среды вызывать такие изменения в химическом строении генетического материала, с помощью которых можно идентифицировать клеточные линии, синтезирующие ДНК. Подобные явления были впервые открыты на фаговой ДНК, однако они верны и для любой ДНК бактериальной клетки. Есть также наблюдения, согласно которым этот феномен справедлив и для эукариотических клеток. В особых случаях могут возникнуть более сложные ситуации. Двустороннее ограничение фага двумя хозяевами иногда наблюдается, но оно не является обязательным.
Фаг, который отвергается клетками, способен адсорбироваться на них и инъецировать свою ДНК. Однако часть последней быстро разрушается и репликации не происходит. Деградация ДНК обусловлена специфическими эндонуклеазами (рестриктазы, или R-нуклеазы), которые способны узнавать особые участки ДНК и расщеплять их, если они не были модифицированы под влиянием М-ферментов. После этого происходит расщепление ДНК экзонуклеазами до отдельных нуклеотидов. Бактериальный штамм может иметь одну или несколько R-нуклеаз и одновременно М-ферменты, которые предохраняют собственную ДНК клетки. Предложена удобная номенклатура этих ферментов. Согласно ряду данных, участки узнавания R-нуклеаз не всегда совпадают с участками расщепления ДНК; возможно, фермент способен мигрировать вдоль цепи, прежде чем он найдет участок, где ДНК подвергнется расщеплению. Модификации обычно заключаются в метилировании аденина или цитозина в специфических участках ДНК. Донором метильных групп при этом служит S-аденозилметионин. Участки, которые способны распознаваться R- и М-ферментами, часто являются палиндромами:
5…pGpTpPupPypApCp…3
3…pCpApPypPupTpGp…5
где Pu и Py обозначают соответственно пурины и пиримидины. Метилирования даже одной цепи вполне достаточно для защиты последовательностей от действия рестриктазы. М- и R-активности входят в состав ферментного комплекса, построенного из нескольких субъединиц. Последние кодируются системой из трех генов. Одна из субъединиц ответственна за распознавание нуклеотидной последовательности.
Функциональная роль модификаций, вызываемых хозяином, неясна. Они способны защитить данный штамм бактерий от массивного разрушения фагами, растущими на различных бактериях. В более общем виде роль модификаций можно определить как защиту от попадания неприемлемой чужеродной ДНК в бактериальную клетку и последующего ее «приживления». Бактерия А, которая отвергает фаг, размножающийся на штамме В, отвергает также и ДНК бактерии В, если ее вводить с помощью конъюгации или трансдукции.
Изменчивость вирусов при пассажах
Изменчивость вирусов при пассажах на животных.
Основоположником этого метода изменения наследственности вирусов был Пастер (1882 год), впервые получивший живую антирабическую вакцину путем серийных пассажей через мозг кролика дикого (вирулентного) штамма вируса бешенства. Было продемонстрировано, что изменчивость вирулентности вируса в процессе пассажей происходит не сразу, а поэтапно, путем последовательных наследственных изменений. Так, при внутримозговом заражении кролика культурой вируса десятого пассажа инкубационный период колебался от 10 до 14 дней, после 21 пассажа он сократился до 7 – 8 дней, а штамм, выделенный после 90-го пассажа, обладал строго фиксированным инкубационным периодом (6 – 7 дней) для кролика и был авирулентным для человека. Селекционированный аттенуированный вариант был использован для иммунизации людей против бешенства. Аттенуированный штамм при подкожном введении утратил патогенность для собак и кроликов, не проникал в слюнные железы, более активно размножался в мозгу кролика, вызывая при этом менее выраженные патоморфологические изменения в ЦНС; как исключение вызывал образование телец Бабеш – Негри и, наконец, быстро инактивировался глицерином. Этими эследованиями был открыт один из методов экспериментального получения живых вакцин против вирусных болезней.
В ряде исследований показана изменчивость отдельных свойств вируса осповакцины при пассажах на животных (кроликах, телятах и других животных). Успешные опыты были проведены с вирусом желтой лихорадки, показавшие возможность значительного усиления нейровирулентности для мышей при пассажах через мозг, с одновременной утратой вирулентности для обезьян при подкожном и внутрибрюшинном введении. В процессе чередующихся подкожных и внутрибрюшинных пассажей был селекционирован авирулентный вариант для мышей, сохранивший иммуногенные свойства. После 260 пассажей селекционированный мутаген использовали для приготовления живой вакцины.
Изменчивость вирулентности и других признаков при пассажах на животных наблюдали у вирусов ящура, чумы свиней, истиной чумы птиц, болезни Ньюкасла, гриппа и др.
Изменчивость вирусов при пассажах на куриных эмбрионах.
Метод пассажей на куриных эмбрионах получил широкое распространение при изучении изменчивости многих вирусов. При пассажах вируса осповакцины отмечали усиление патогенности для куриного эмбриона и для кролика при подкожном введении. Был выведен мутант по ряду генетических признаков, сходных с вирусом натуральной оспы.
Вирус бешенства после 30 пассажей на куриных эмбрионах приобрел способность проникать во все ткани и органы зародыша, а после 75 пассажей утратил вирулентность для кролика при заражении в мозг.
При пассажах в куриных эмбрионах вируса ящура наблюдали снижение вирулентности для крупного рогатого скота, а после 120 пассажа вирус утратил не только патогенные, но и иммуногенные свойства. Пассажами в куриных эмбрионах был получен высокоиммуногенный штамм Н вируса ньюкаслской болезни, нашедший широкое применение для приготовления живой вакцины. При пассажах на утиных эмбрионах было выделено несколько вакцинных штаммов этого вируса.
Однако при длительных пассажах в куриных эмбрионах не всегда отмечали снижение вирулентности (вирусы ньюкаслской болезни, инфекционного аборта лошадей, лошадиного энцефаломиелита, западнонильского и венесуэльского энцефалитов, оспы птиц, классической чумы птиц и др.) Так, из 16 штаммов вируса чумы крупного рогатого скота только один штамм утратил вирулентность, сохранив иммуногенные свойства. Изменчивость вирусов при пассажах в куриных эмбрионах зависит от ряда условий: свойств штамма вируса, возраста эмбрионов, способа их заражения, чередования пассажей на восприимчивом животном и эмбрионе и других факторов.
Изменчивость вирусов при пассажах на культуре клеток.
Культура клеток – одна из наиболее применяемых методов для изменения вирулентности и других свойств вирусов. Широкое использование этого способа дало возможность получать наследственно измененные варианты вирусов и селекционировать их из генетически неоднородной вирусной популяции. Кроме первично трипсинизированных тканей, с этой целью широко использовали перевиваемые линии клеток. Причем отдельные клеточные линии могут быть высокочувствительными к определенным вирусам, но тем не менее в них вирус не накапливается в высоких титрах. Другие же клеточные линии, обладают пониженной чувствительности к вирусам, обеспечивает накопление их высоких концентрациях.
Используя перевиваемую или первичные культуры ткани, были выделены аттенуированные штаммы вируса классической чумы птиц. Вирулентность вируса японского инцефалита была ослаблена пассированием в культуре клеток почки Хомика с последующими пассажами на мышах–сосунках. Выделенный вариант вируса утратил вирулентность для мышей при внутримозговом заражении и не передавался комарами.
За последние годы появилось много обнадеживающих сообщений о получении стабильных вирулентных штаммов вируса ящура при пассаже на различных типах культуры тканей. После многократных пассажей в культуре тканей куриного эмбриона вируса желтой лихорадки наблюдали утрату нееротропных и висцетропных свойств для обезьян с сохранением иммуногенности. Этот вариант (штамм 17 Д) успешно используется для приготовления живой вакцины.[4]
При пассажах вируса полиомиелита в культуре почечной ткани обезьян был выделен ряд мутантов, не вызывающих у обезьян параличей при введении в головной и спиной мозг. Авирулентные штаммы были получены при быстро следующих друг за другом пассажах, благодаря чему создаются благоприятные условия для отбора вариантов, обладающих наибольшей активностью размножения. Сочетая отбор с многократными пассажами, были выделены мутанты с наследственно ослабленной нейровирулентностью для обезьян. Для снижения вирулентности необходимо было проделать 20-40т пассажей.
Изменчивость вирусов, возникающая в процессе пассажей при повышенных и пониженных температурах.
Убедительные данные об ослаблении вирулентности в процессе пассажей при пониженной температуре полученных в опытах с вирусом ящура. Вирус пассировали на первичных культурах почки телят при 37°, 28° и 22° . Вирулентность проверяли на свиньях и мышах сосунках. Как выяснилось, все авирулентные для свиней варианты не размножались в культуре клеток при 40°. В то же время вирулентные и авирулентные штаммы одинаково активно размножались при 28°, а при 22° активно репродуцировались только авирулентные варианты. Однако не у всех авирулентных для свиней штаммов оптимум температуры размножения был ниже 37°. Отдельные варианты более активно размножались при 37° и не размножались при 22°. Таким образом, не во всех случаях ослабление вирулентности для свиней и мышей сопровождалось изменением активности репродукции при пониженных температурах, что не дает основания считать основание абсолютной взаимосвязи у ящура между вирулентностью и активностью репродукции при пониженной и пониженной температурах.
В ряде наблюдений была так же показана возможность получение авирулентных вариантов у других вирусов в процессе пассажа их при пониженных температурах (гриппа, чумы свиней, полиомиелита, кори, японского энцефалита и др.). Так, при пассажах вирусах гриппа на куриных эмбрионах при пониженных (32-25°С) температурах были селекционированые мутанты, более активно размножающие при 28° и 32°, чем при 36°. У них закономерно наблюдалось ослабление или утрата вирулентности и токсических свойств. И, наоборот, мутанты селекционированные в процессе пассажей при повышенных температурах, более активно размножались при 39° и 41°, чем исходные штаммы. Варианты утратили свойства размножаться при 25-28°С.
Сходные результаты были получены в опытах с другими видами вирусов. На основании проведенных исследований можно сделать выводы, что пассажи вирусов при пониженных температурах, как правило, приводили к появлению вариантов с ослабленной вирулентностью. Снижение или полная утрата ее в ряде случаев коррелировали с утратой способности размножаться при повышенных температурах.
Причина изменчивости вирусов при пассажах еще мало выяснены. Согласно селекционной теории, культура вируса генетически не однородна и в процессе пассажей на животных или культуре клеток происходит отбор и накопление вариантов для которого данное условие культивирование более благоприятно. Другая возможная причина возникновения вариантов при пассажах - рекомбинация вирусов.[5]
Изменчивость вирусов при пассажах принципиально не отличается от мутации, возникающих при воздействии физическими и химическими мутогенами, так как в обоих случаях изменения признаков у вируса связаны с изменениями в его генотипе. Различия между ними несущественные, главным образом количественные. Одна из особенностей изменчивости при пассажах состоит в том, что для изменения генетических признаков, особенно таких сложных полигинных признаков, как патогенность или репродуктивная активность, требуется ряд мутаций, возникающих в серии пассажей. Такого типа многоступенчатые мутации являются отражением процесса перехода количественных изменений в качественные. Следует при этом отметить, что изменения свойств у вирусов при пассажах возникают раньше, чем мы их обнаруживаем. С момента появления первых изменений до выявления их проходит, какой то период количественных изменений, которые на каком то этапе пассажей переходит в качественные, существенно изменяющие наследственные признаки вируса.
Химические мутагены
Предложено три классификации химических мутагенов:
Рехборна, Фриза, Раппопорта.
Фриз предложил разделить мутагены на две основные группы:
1) мутагены, реагирующие с нуклеиновой кислотой только во время ее репликации;
2) мутагены, вступающие в реакцию с покоящейся молекулой нуклеиновой кислоты, но требующие для формирования мутащий последующих ее репликаций.
В основе молекулярных изменений вирусной нуклеиновой кислоты, приводящих к мутации, лежат два основных процесса: замена основания или вставка основания. Различают два типа замены оснований: простую (транзиция) – на место одного пуринового основания встает другое или одно пиримидиновое основание заменяется другим; сложную (трансверсия) – вместо пуринового основания появляется пиримидиновое или пиримидиновое основание заменяется пуриновым. Вставка основания – ведет более к глубоким изменениям генетического кода, чем простая замена оснований. В то же время основой изменения генетического признака, имеющего одно и то же фенотипическое выражение, могут быть мутационные повреждения различных генов.
Кроме простых замен, алкилирующие агенты способны индуцировать сложные замены – пурин на пиримидин. Мутагенное действие этих соединений было показано с вирусами ньюкаслской болезни и клещевого энцефалита.
Гидроксиламин индуцирует мутации по типу образования простых замен оснований в нуклеиновой кислоте, направление которых зависит от типа нуклеиновой кислоты, которую содержит вирус. С помощью гидроксиламина были индуцированы мутации у вирусов герпеса, ньюкаслской болезни, полиомиелита.
В последнее время был синтезированный и изучен один из аналогов гидроксиламина – оксиметилгидроксиламин (ОМГА), реагирующий только с цитозином, но не с урацилом РНК, а следовательно, обладающий более высокой специфичностью и одной направленностью мутагенного действия.
Для вирусов человека и животных мутагеном является и формальдегид, с помощью которого были индуцированы мутанты у вируса полиомиелита и вируса западного энцефаломиелиталошадей при воздействии на очищенную РНК и внутриклеточный вирус. Механизм мутагенного действия формальдегида недостаточно изучен.
Механизм действия азотистой кислоты (НNО2) как мутагена на нуклеиновые кислоты заключается в дезаминировании органических оснований, т. е. отщепление от их молекул аминогруппы (NH2).
Физические мутагены
Мутагенное действие ультрафиолетового излучения. Действие УФ лучей как мутагенов состоит в том, что они взаимодействуют с молекулами нуклеиновых кислот и поглощаются ими, особенно лучи с длинной волны 260 – 280 нм. Попадая в молекулу нуклеиновой кислоты, они поглощаются входящими в ее состав органическими основаниями. Оказалось, что тимин, урацил, цитозин более чувствительны к ультрафиолетовым лучам, чем аденин и гуанин. При облучении УФ-лучами две соседние молекулы тиминов соединяются друг с другом в пары, образуя так называемые тиминовые димеры.
Под влиянием УФ-облучения получен мелкобляшечный мутант вируса западного лошадиного энцефаломиелита, обладающий стабильным S-фенотипом в культуре клеток ФКЭ. Установлена принципиальная возможность получения мутаций при воздействии УФ-лучей на репродуцирующийся вирус и его нуклеиновую кислоту, в которой происходят структурные нарушения РНК: компонент ее – урацил – образует диаметр и гидраты.
Заключение
Не все мутации, образующиеся под влиянием мутагенов, одинаково стабильны. Мутанты, полученные при действии повышенной температуры, кислой среды, ультрафиолетовых лучей и ультразвуковых волн, давали около 20% реверсий, при воздействии профлавина все мутанты оказались полностью стабильными. Эти различия в стабильности связаны с неодинаковым молекулярным механизмом действия использованных мутагенов. Повышенная температура, кислая среда, ультрафиолетовые лучи вызывают главным образом локальные изменения вирусной нуклеиновой кислоты, ведущие к замене отдельных оснований. При мутагенном действии профламина, а также частично азотистой кислоты причиной мутаций являются выпадения или вставки оснований. При получении вакцинных вирусных штаммов путем воздействия на вирус мутагенами целесообразно использовать мутагены, вызывающие более глубокие изменения генетического кода – типа выпадений или вставок, так как такие мутанты обладают стабильностью наследственных свойств.
Эволюция вирусов в природе идет в различных направлениях, т. е. изменение патогенности вируса, спектра патогенности, антигенные, иммуногенные свойства вирусов и т. д. Эволюция разных видов имеет свои особенности. Одно из важных изменений, это изменение круга хозяев, приспособление к бактериям, грибам, насекомым, растениям.
Список литературы:
1. Сюрин В. Н. Ветеринарная вирусология. – 2-е изд., перераб. и доп. – М.: Агропромиздат, 1991. 431 с., с ил.
2. Горьский Б. В. И др. Основы общей ветеринарной вирусологии. 2-е изд., М., «Колос», 1978., 192 с., с ил.
3. Гендон Ю. З. Общая и частная вирусология. – М.: Мир, 1981. – 680 с., с ил.
4. Земсков М. В. И др. Основы общей микробиологии, вирусологии и иммунологии. – 2-е изд., испр. и доп. М., «Колос», 1977., 312 с., с ил.
5. Под редакцией Сюрина В. Н. Руководство по ветеринарной вирусологии. – М., «Колос», 1966., 686с.
Похожие работы
... Как бы не менялся фенотип организма в ответ на воздействия внешней среды – его изменения не могут привести к изменению генов, которые этот организм передаст следующему поколению. 1.4 Роль хромосомных и геномных мутаций в эволюции Все перечисленные выше характеристики верны для всех типов мутаций – генных, хромосомных и геномных. Однако, такие геномные и хромосомные мутации как полиплоидия ( ...

... по численности и плотности популяции животных хозяйств. Все это приводит к возникновению неизвестных ранее возбудителей, изменение свойств и путей циркуляций известных ранее вирусов, а также к значительным изменениям восприимчивости и сопротивляемости человеческих популяций. Влияние загрязнения внешней среды. Современный этап развития общества связан с интенсивным загрязнением ...
... растений. Примером тому может служить новый сорт пеларгонии Velvet Rose, полученный через каллусную культуру. Для ускорения селекционного процесса в культуре клеток используются химические и физические мутагены. Обработка ткани раувольфии змеиной азотистым ипритом в концентрации 2,5 * 10-3 М привела к повышению уровня аберраций хромосом в первом пассаже до 32%, вызвала сдвиг популяции в сторону ...
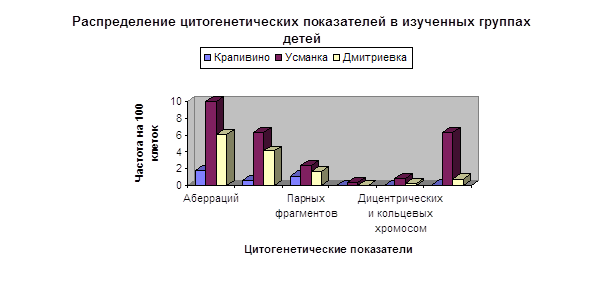
... Глава II. Материалы и методы 2.1. Характеристика обследованных групп. В соответствии с поставленными задачами проведено цитогенетическое обследование в группах девочек-подростков, проживающих в с. Усманка и с. Дмитриевка Чебулинского района Кемеровской области. В качестве группы сравнения была использована группа девочек близкого возраста, проживающих в п. Крапивинский Кемеровской области - ...
0 комментариев