Введение
Дарвинизм — это материалистическая теория эволюции органического мира Земли, основанная на воззрениях Ч. Дарвина. Фундаментом для создания теории эволюции Ч. Дарвину послужили наблюдения во время кругосветного путешествия на корабле «Бигл», исследования и обобщение достижений современной ему биологии и селекционной практики.
В 1859г. была напечатана книга Ч. Дарвина «Происхождение видов путём естественного отбора, или Сохранение благоприятствуемых пород в борьбе за жизнь». В этой книге Ч. Дарвин показал, что преобразование пород домашних животных и культурных растений происходит на основе незначительных изменений в признаках отдельных организмов. Возникающие у животных и растений наследственные изменения попадают под действие естественного отбора, так что в борьбе за существование выживают формы, наиболее приспособленные к данным условиям среды. Тем самым Дарвин материалистически объяснил целесообразность организации живых существ. Таким образом, он впервые в истории биологии построил теорию эволюции, руководствуясь, в частности, данными, полученными хозяйственной практикой.
Дарвин исходил из существования двух основных типов изменчивости: определённой (представляющей собой приспособительные реакции организмов на воздействие факторов внешней среды) и неопределённой (также возникающей под влиянием внешних факторов, но не имеющей приспособительного характера). Материалом эволюции может служить только неопределённая (наследственная) изменчивость, основанная, на мутациях и их комбинациях, возникающих в результате скрещивания. Главным движущим фактором эволюции служит естественный отбор.
Заслуга Дарвина в том, что он вскрыл движущие силы органической эволюции. Дальнейшее развитие биологии углубило и дополнило его представления, послужившие основой современного дарвинизма. Развитие дарвинизма стимулировало прогресс многих областей биологии. Во всех биологических дисциплинах ведущее место занимает теперь исторический метод исследования. В результате эволюционной трактовки факты, добытые наукой, в свою очередь, способствовали дальнейшей разработке проблем дарвинизма.
Современный дарвинизм служит важнейшей теоретической основой биологии, сельскохозяйственной и медицинской практики: только дарвинистический подход даёт возможность эффективного преобразования пород домашних животных и сортов культурных растений, выведения новых, более продуктивных штаммов микроорганизмов; дарвинизм создаёт основу для представления о биосфере как о сложнейшей развивающейся системе и в перспективе даёт возможность управления эволюционным процессом.
Предпосылки эволюции:
изменчивость и наследственность
Изменчивость и наследственность как свойство живого представляют собой предпосылки эволюции жизни. Главной целью их познания должно быть: 1) выяснение роли этих явлений в органической эволюции и 2) доказательство невозможности сведения эволюции к этим предпосылкам. Решение упомянутых задач возможно только при более близком знакомстве с изменчивостью и наследственностью.
Формы изменчивости, основные понятия и термины
Из матричного принципа строения и работы генетического кода с неизбежностью следуют фундаментальные явления живой природы — наследственность (передача основных генетических структур от поколения к поколению) и изменчивость (разнородность, разнокачественность, наличие различий между особями). Наследственность определяется самим принципом передачи наследственного кода посредством процесса, сходного с получением оттисков, копий с одной и той же печати (матрицы). Изменчивость определяется, во-первых, не абсолютной стабильностью молекул, образующих цепочку генов (в результате разного рода причин в этих молекулах время от времени происходят изменения — мутации, ведущие к изменению кода наследственности), и, во-вторых, влиянием внешних условий, определяющих конкретное проявление того или иного генетического зачатка в данных конкретных условиях. Можно сказать поэтому, что природа наблюдаемой изменчивости всегда двойственна, противоречива. Эта двойственность долгое время ставила исследователей в тупик, была причиной многих дискуссий в биологии и теории эволюции, начиная с работ Ж. Ламарка и Ч. Дарвина. Ч. Дарвин различал несколько форм изменчивости. В процессе дальнейших исследований, подтвердивших дарвиновские представления, была выработана новая терминология, которая должна быть, сопоставлена с дарвиновской (см. табл.).
| Терминология Ч.Дарвина | Современная терминология |
| Вся наблюдаемая изменчивость: 1. Наследственная, неопределенная, индивидуальная изменчивость 2. Ненаследственная, определенная, массовая изменчивость 3. Соотносительная (коррелятивная) изменчивость 4. Изменчивость, возникающая вследствие скрещивания | Фенотипическая изменчивость: 1.Изменчивость генотипическая (мутации) 2.Изменчивость паратипическая (модификации) 3.Коррелятивные изменения 4.Комбинативная изменчивость |
С новой терминологией тесно связаны некоторые понятия, которые значительно облегчают понимание процессов изменчивости и наследственности:
1) Генотип и фенотип. Термины эти были предложены Иоганнсеном (1903). Фенотип — это вся совокупность признаков и свойств любого индивидуума — является результатом взаимодействия между генотипом и средой. Под генотипом понимают взаимно связанную систему единиц наследственности (генов), наследственную программу развития.
Как правило, различные генотипы обусловливают развитие различных фенотипов. Изменение генотипа влечет за собой и изменение фенотипа.
2) Фенотипическая изменчивость. В результате простых наблюдений не всегда ![]()
![]()
![]() можно сказать, какой характер изменчивость носит, т.е. определяется ли она изменениями генотипа (наследственно обусловленная изменчивость, или мутация) или она ненаследственна (модификация). Изменения проявляются в конкретных морфофизиологических, видимых или фенотипических изменениях: цвета, запаха, вкуса, формы, пропорций, размера, числа частей и т. д. Следовательно, в условиях простого наблюдения можно говорить лишь в самой общей форме о фенотипической изменчивости. И только эксперимент (в частности, скрещивание) может показать, какова доля наследственной (генотипической) и ненаследственной, определяемой влиянием конкретных факторов в развитии данной особи (паратипической) изменчивости в общей фенотипической изменчивости.
можно сказать, какой характер изменчивость носит, т.е. определяется ли она изменениями генотипа (наследственно обусловленная изменчивость, или мутация) или она ненаследственна (модификация). Изменения проявляются в конкретных морфофизиологических, видимых или фенотипических изменениях: цвета, запаха, вкуса, формы, пропорций, размера, числа частей и т. д. Следовательно, в условиях простого наблюдения можно говорить лишь в самой общей форме о фенотипической изменчивости. И только эксперимент (в частности, скрещивание) может показать, какова доля наследственной (генотипической) и ненаследственной, определяемой влиянием конкретных факторов в развитии данной особи (паратипической) изменчивости в общей фенотипической изменчивости.
Любой наблюдаемый признак есть видимый результат реализации наследственной программы развития в данных условиях. Поэтому признаки сами по себе, строго говоря, ненаследственны. Признаки зависимы, с одной стороны, от генетических особенностей организма, а с другой — от условий жизни. Это доказано совершенно твердо на огромном количестве фактов.
Например, известно, что породные признаки культурных животных проявляются только в условиях соответственного кормления и общего благоприятствующего содержания. При плохом кормлении и содержании не проявятся и типичные наружные признаки породы (рис.1), ее экстерьер.

Рис.1. Пример ненаследственных различий.
Два поросенка одного помета стали несходными
вследствие различного кормления.
Весь спектр возможных изменений данного генотипа при разных условиях развития получил название нормы реакции. Таким образом, можно сказать, что наследуется не признак, а норма реакции генотипа.
Ненаследственные (паратипические модификационные) фенотипические изменения есть реакция конкретного генотипа на разные условия среды. В разных условиях среды один и тот же генотип будет выражен различными фенотипами.
Наследственные изменения – мутации
Мутации — это наследственные изменения генотипа. Это изменение нормы реагирования выражается обычно в появлении новых признаков и свойств в изменении нормы реагирования на условия среды. По своей природе мутации могут быть весьма различны, хотя во всех случаях являются дискретными изменениями общего кода наследственной информации. Различают:
ü генные мутации — единицами изменения являются гены или локусы в хромосомах;
ü хромосомные мутации — единицами изменения являются структурные перестройки отдельных хромосом (инверсии, дупликации, делеции, транслокации);
ü геномные мутации — единицами изменения является число хромосом;
ü внеядерные мутации — касаются изменения некоторых константных и более или менее автономных внеядерных структур клетки (митохондрий, пластид и т. п.).
Эволюционная характеристика мутаций
Важнейшими с эволюционной точки зрения характеристиками мутаций являются частота возникновения и встречаемость их в природных популяциях, а также влияние мутаций на те или иные признаки организма.
Частота возникновения отдельных спонтанных мутаций выражается в числе гамет одного поколения, содержащих определенную мутацию по отношению к общему числу гамет. Эти частоты, точно определенные для довольно значительного числа растений, животных и микроорганизмов, оказываются весьма близкими по величине и составляют 10-5-10-7 (т. е. 1 из 100000-10000000 несет вновь возникшую мутацию в определенном локусе). Частота мутаций неодинакова для разных генов, у отдельных лабильных генов растений она достигает 10-2. Общая частота мутаций, которая складывается из частот мутаций отдельных генов, также оказывается близкой у разных организмов: от нескольких процентов (одноклеточные водоросли, низшие грибы, бактерии) до 25% (дрозофила) всех гамет одного поколения несут ту или иную мутацию.
При воздействии облучением, определенными химическими веществами, температурой частота мутаций резко повышается и может достигать очень значительных величин. На частоту возникновения мутаций оказывает влияние физиологическое состояние организма.
Спектр мутантных признаков, или признаков, затрагиваемых мутациями, оказывается очень широким. Нет признаков и свойств, которые в той или иной степени не затрагивались бы мутациями. Наследственной изменчивости подвержены все морфологические, физиологические, биохимические, этологические признаки и свойства. Эти вариации выражаются как в качественных различиях, так и количественно, т.е. по средним значениям варьирующих признаков. Мутации могут происходить как в сторону увеличения, так и в сторону уменьшения выраженности определенного признака или свойства. Мутации бывают или очень резко выражены (вплоть до летальности), или представлены в виде незначительных отклонений от исходной формы (так называемые «малые» мутации). В качестве примера «большой» мутации на рис.2 изображена дрозофила с мутацией tetraptera; эта мутация привела к возникновению признака, характерного для другого отряда (четыре крыла вместо обычных для мух двух крыльцев). Во многих работах показано, что мутации затрагивают такие существенные биологические признаки, как общая жизнеспособность, способность к скрещиванию, плодовитость и скорость роста и т.п. То обстоятельство, что мутации могут затрагивать все без исключения признаки и свойства организмов, является их важнейшим эволюционным свойством: эти наследственные изменения и являются материалом для эволюции.
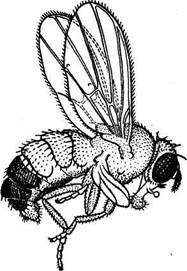
Рис.2. Мутация tetraptera у Drosophila melanogaster.
Видна развитая вторая пара крыльев.
Другим важнейшим эволюционным свойством мутаций является их встречаемость в природных популяциях. Изучение большого числа природных популяций разных видов, особенно активно развернувшееся в 30—40-е годы нашего столетия, подтвердило вывод С. С. Четверикова о насыщенности всех популяций разнообразными мутациями. В разных популяциях частоты разных мутантных генов оказывались весьма различными. Практически нет двух популяций, которые имели бы совершенно одинаковые частоты встречаемости и спектры мутантных признаков. При этом близко расположенные, соседние популяции могли отличаться друг от друга столь же значительно, как и далеко расположенные популяции.
Характерные особенности мутаций, высокая и постоянная частота возникновения, затрагивание мутациями любых, в том числе и биологически важных, признаков, насыщенность природных популяций мутациями свидетельствуют о том, что мутации как элементарные единицы наследственной изменчивости могут рассматриваться в качестве элементарного эволюционного материала. Лучшим доказательством этого вывода служит обнаружение в природе генетических различий по отдельным мутациям между расами, подвидами и близкими видами.
Генетические различия между близкими группами
Существование генетических (основанных на немногих мутациях) различий по отдельным признакам между близкими популяциями в природе является убедительным доказательством того, что именно мутации являются элементарным материалом.
По изучению генетического состава природных популяций проведено довольно много работ на разных группах растений (львиный зев, фиалки, пикульник и др.), беспозвоночных (дрозофилы, непарный шелкопряд, божьи коровки и др.) и позвоночных (мышевидных хомячков, домовых мышей, крыс, полевок и др.). Во всех без исключения случаях, когда удавалось провести достаточно точный генетический анализ признаков, по которым различаются сравниваемые популяции, подвиды или близкие виды, было найдено расщепление изученных наследственных признаков.
Во всех случаях таксоны отличались друг от друга лишь комбинациями генных, хромосомных или геномных мутаций. Это дает основание полагать, что именно мутации как элементарные наследственные изменения действительно являются элементарным эволюционным материалом.
Ненаследственные изменения – модификации
Термин модификация был предложен датским генетиком В. Иоганнсеном. В широком смысле слова под модификациями следует понимать ненаследственные изменения, возникшие под влиянием факторов абиотической и биотической среды в пределах реализации одной нормы реакции. К первым принадлежат температура, влажность, свет, химические свойства воды и почвы, механически действующие факторы (давление, ветер и т. д.); ко вторым — пища, а также прямое и косвенное воздействие других организмов. Все эти факторы вызывают ненаследственные фенотипические изменения более или менее глубокого характера
В естественной обстановке на организм влияют, разумеется, не отдельные факторы, а их совокупность. Однако некоторые факторы среды имеют ведущее значение. Так, можно утверждать, что главнейшее модифицирующее значение имеют температура, влажность и свет, а для водных организмов — солевой состав воды. Конечно, не надо забывать, что соответственное значение факторов среды определяется генотипическими свойствами конкретного организма, его нормой реакций.
Температура определяет очень многообразные изменения. Так, под влиянием различной температуры цветки китайской примулы приобретают различную окраску. При 30-35°С развиваются белые цветки, а при 15-20°С — красные цветки.
Подобные явления наблюдаются и у животных. Так, у бабочек-крапивниц повышение температуры вызывает в окраске крыльев усиление красных и желтых тонов. Окраска кожи млекопитающих и их волосяного покрова также в ряде случаев связана с температурным воздействием. Так, удаление шерсти на отдельных участках кожи у кроликов в одной из наследственных групп (линий) при содержании этих животных на холоде вызывало развитие (на выбритых местах кожи) черного пигмента с последующим ростом черных волос. Известно, что волосяной покров млекопитающих под влиянием пониженной температуры достигает более пышного развития. Температура влияет также на развитие форм тела животных. Так, воспитание новорожденных мышей в тепле приводит к более слабому развитию шерсти, к удлинению ушей и хвоста.
Под влиянием фактора влажности удивительные превращения наблюдаются у растений. Подводные листья стрелолиста имеют вытянуто-лентовидную форму, надводные на том же экземпляре — типичную стреловидную. У животных фактор влажности также вызывает явственные изменения. Прежде всего влажность влияет на окраску. Сухость вызывает у лягушек посветление, повышение влажности стимулирует потемнение кожных покровов. Под влиянием влажности после каждой линьки у многих птиц наблюдается потемнение рисунка пера.
Свет также вызывает глубокие изменения, в особенности у растений, изменяя форму и размеры стебля и листьев, а также обусловливая анатомические изменения органов. При недостаточном освещении у дикого латука изменяется форма стебля, диаметр которого становится меньше; появляется полегаемость; листьев мало, они свисают, форма их изменена; листовые пластинки тонкие, у них редуцированы и изменены некоторые ткани и т. д.
Влияние химизма среды также имеет модифицирующее значение. Для растений основное значение имеет минеральное питание, изменение состава которого вызывает у них преобразование формы. Для развития, например, высших растений необходимо присутствие следующих зольных элементов: К, Са, Mg, S, Р и Fe. Отсутствие одного из перечисленных элементов изменяет форму развития.
Формообразующее воздействие химизма питания на животный организм также велико. При неправильном кормлении животные не достигают полного развития, а поэтому типичные признаки вида или породы остаются невыраженными.
Организмы изменяются также под косвенным и прямым воздействием других организмов. Сосна, выросшая на свободе приобретает широкую крону, напоминающую крону дуба тогда как дуб, выросший в густом лесу, приобретает мачтовый ствол.
При изучении модификаций уже Ч. Дарвином была вскрыта весьма характерная черта. Модификации всегда строго закономерны и специфичны. Один и тот же фактор вызывает у разных организмов различные модификаций в соответствии с различиями их генотипов (их норм реакции).
Так, повышение температуры вызывает у прыткой ящерицы (Lacerta agilis) посветление кожных покровов, у стенной ящерицы (Lacerta muralis) — потемнение. У антилопы-гну (Африка) в условиях украинской зимы развивается зимний волосяной покров; у оленебыка (Африка) в тех же условиях сохраняется летний наряд. Реакция на один и тот же фактор оказывается различной в зависимости от различия генотипических особенностей данных форм. При этом в условиях, нормальных для данного вида, все эти модификации оказываются явно приспособительными (адаптивными).
Другой особенностью модификационной изменчивости является тот факт, что модификационные изменения одного и того же организма оказываются различными на разных стадиях развития и при разных физиологических состояниях.
Третья особенность модификаций — их ненаследственность. Поколение одной и той же формы в разных условиях дает разные модификации.
Ч. Дарвин указывал, что в разных точках семенной коробочки условия различны и что каждое отдельное семя развивается в индивидуальных условиях. В каждом семени проявятся индивидуальные модификации.
Ненаследственность индивидуальных модификаций доказал В. Иоганнсен в своих наблюдениях над чистыми линиями фасоли. Чистой линией называют серию поколений, полученных от одного конкретного самоопыляющегося растения. Такая чистая линия некоторое время остается генотипически относительно однородной.
В. Иоганнсен показал, что, несмотря на наследственную однородность материала одной чистой линии, семена потомства ее различны по величине, массе и другим признакам и что эти различия являются последствием индивидуальной дифференцировки в несколько иных условиях развития каждого семени.
Ненаследственную изменчивость нельзя сводить только к индивидуальным модификациям. Особи любого вида связаны видовой общностью. Поэтому каждая особь, каждый отдельный генотип, каждая норма реакции должна быть единством отдельного и общего.
Например, кожа травяной лягушки под влиянием пониженной температуры темнеет. Мех лисицы после прохладного лета зимой становится более пушистым и полноволосым и т.д. Во всех этих случаях налицо модификационные изменения, носящие однотипно-общий, групповой характер. На фоне этой общей модификации разыгрывается процесс модификационной индивидуализации, которая имеет то же направление; например, при низкой температуре все травяные лягушки темнеют, но каждая — в различной степени.
Таким образом, следует различать индивидуальные модификации, массовые, или групповые, модификации. Наличие их показывает, что определенная форма модификационной изменчивости исторически обусловлена и является конкретным видовым свойством.
Корреляции
Корреляции — вторичные изменения, возникающие в ходе онтогенетического развития под влиянием какого-либо первичного изменения. Фенотипически корреляции выражаются в соотносительном изменении функций и структуры органа или его части, в зависимости от изменений в функции и структуре другого органа или его части. Корреляции являются, следовательно, соотносительными функциональными изменениями органов или их частей.
Учение о корреляциях было введено Ч. Дарвином. Эволюция видов идет через процесс их приспособления к меняющимся условиям среды и что расхождение (дивергенция) видов идет по руслу их приспособительной дифференцировки.
Казалось бы, в практике исследования виды должны четко различаться по приспособительным признакам. Однако в действительности этого сплошь и рядом нет. Напротив, в очень 6oльшом числе случаев виды значительно явственнее различаются признакам, приспособительное значение которых не ясно. В таком случае становится непонятным, каким образом возникают эти не явно приспособительные признаки различий между видами. Ведь они, казалось бы, не могли быть накоплены отбором, как отбор накапливает полезные, приспособительные признаки.
Ч. Дарвин для объяснения этого кажущегося противоречия привлек понятие о корреляциях. Он указал, что для систематики ценность признаков, не существенных в смысле их приспособительного значения, зависит преимущественно от их корреляций с другими, малозаметными, практически недостаточными для определения, но приспособительными признаками. Несущественные в адаптивном отношении признаки возникают, следовательно, не под прямым влиянием отбора, а косвенно, т. е. в силу коррелятивной зависимости от других фенотипически нечетких, но приспособительных признаков. Отсюда следует, что нужно различать ведущие приспособительные изменения и изменения коррелятивные, зависимые. Если возникло приспособительное изменение, то оно влечет за собой возникновение ряда зависимых коррелятивных признаков.
Вопрос о коррелятивной изменчивости важен, т.к. если какие-либо органы изменяются в приспособительном направлении, то вместе с ними изменяются и другие, «без всякой видимой пользы от изменения».
Следовательно, явления корреляции объясняют возникновение и сохранение неявно адаптивных особенностей в потомстве. Другой важной стороной для Ч. Дарвина была проблема целостности организма. Изменение в одной части связано с изменениями во всех или многих других частях организма.
Классификация корреляций
Материал, собранный Ч. Дарвином, показал многообразие коррелятивных связей и значительный теоретический и практический интерес явления. В последарвиновскую эпоху проблема корреляций разрабатывалась многими авторами второй половины XIX и в XX в. Однако особенно важны исследования академика И. И. Шмальгаузена.
Различают следующие типы корреляции:
1) Морфогенетические корреляции приурочены преимущественно к эмбриональной фазе онтогенеза. Именно на примерах этих корреляций четко выявляется природа коррелятивных зависимостей. С первых стадий развития (дробления) яйца и последующего-органогенеза морфогенетические или формообразовательные корреляции играют ведущую роль в эмбриогенезе.
Морфогенетические коррелятивные изменения происходят вследствие возникновения первичных изменений, которые влекут за собой соответственные зависимые вторичные изменения.
2) Эргонтические корреляции по преимуществу относятся к постэмбриональному периоду онтогенеза, но особенно характерны для ювенильного периода. Их значение: окончательная моделировка индуцируемых частей. Эргонтические (рабочие) корреляции возникают в результате контакта между соответственными частями организма. Особенно отчетливо выявляются эргонтические корреляции во взаимоотношениях между работающей мышцей и подлежащей костной опорой. Чем сильнее развита мышца, тем резче развиваются гребни на участках кости, к которым мышца прикрепляется. Поэтому по развитию костных гребней можно судить о степени развития мышцы, служащей эргонтическим индуктором.
Модификации, мутации и корреляции создают, таким образом, огромное многообразие индивидуумов, причем потенциальные возможности этого многообразия далеко не исчерпываются. Многообразие возрастает также под влиянием комбинативной изменчивости.
Причины изменчивости
В возникновении изменений организма ведущую роль играют внешние причины. По отношению к модификациям это положение не требует особых объяснений, так как модификации являются реакциями организма на внешние воздействия, определяемыми нормой реакции.
В отношении корреляций ясно, что зависимые изменения, протекающие в каком-либо органе или его части, являются реакцией на внешние по отношению к этому органу или его части воздействия; всякое первичное изменение в одном органе индуцирует изменения в другом. В системе одного и того же организма вторичного коррелятивного изменения не произойдет, если не было первичного. Следовательно, корреляции могут рассматриваться как особый тип модификационной изменчивости, определяемой внутренней средой самого организма. Только в случае изменения самого генотипа, а следовательно, и нормы реакции, другими словами, в случае мутации, характер модификаций и формы корреляций изменяются. Проблема причин изменчивости упирается в вопрос о причинах наследственных изменений (мутационных).
В чем же причина возникновения наследственной изменчивости мутации разных типов? Причины заключаются в физико-химических особенностях строения молекул ДНК, с одной стороны, и влиянии различных внешних факторов (мутагенов) — с другой. Первая причина возникает из того простого факта, что молекула ДНК состоит в конечном итоге из атомов. Атомы состоят из электронов и других элементарных частиц, находящихся в постоянном движении. Время от времени совершенно спонтанно и без участия каких-либо внешних сил могут происходить переходы электронов с орбиты на орбиту и т. п. Эти ничтожные изменения в микромире в конце концов должны как-то сказаться и на строении молекул, слагающих ДНК.
Спонтанный мутационный процесс определяет, наверное, половина мутаций, в основном на генном уровне. Внешние по отношению к коду наследственной информации факторы определяют появление мутаций на всех уровнях: генном, хромосомном, генотипном и внеядерном.
Мутации могут возникать под воздействием внешних факторов, воспринимаемых организмом. Однако характер возникающего изменения определяется не внешним фактором, а изменениями генотипа организма. Вот почему один и тот же фактор может вызвать самые различные мутации. Внешний фактор не определяет направления изменчивости, она остается единичной, объективно случайной, неопределенной.
Изменчивость и наследственность
как предпосылки эволюции
Рассмотрение изменчивости и наследственности имеет огромное принципиальное значение. Можно ли объяснить эволюционный процесс только изменчивостью и наследственностью? Каковы данные для разрешения этого вопроса?
Мутации как элементарный материал эволюции
Мутации определяют характер модификаций. Если произошла мутация, то изменяется норма реакции, а стало быть, и модификации. Мутации могут влиять и на корреляции. Если возник резкий мутационный сдвиг, который существенно изменил норму реакции клеток, то это может повести и к изменениям в коррелятивных зависимостях. Мутации отзовутся и на комбинативной изменчивости.
Каково же эволюционное значение мутаций? Ответ на этот вопрос определяется следующими данными. Если возникла какая-либо мутация, которая значительно изменила норму реакции, то такая мутация может нарушить или даже разрушить установившиеся коррелятивные связи, а вместе с ними и приспособленную организацию развивающегося животного или растения. Тогда, естественно, снизится жизнеспособность или даже она будет вообще утрачена на том или ином этапе индивидуального развития организма. Именно поэтому мутации очень часто имеют смертельное (летальное) значение, как это, например, известно и отношении многих мутаций у дрозофил. Таким образом, мутации относительно вредны в каждый данный момент эволюции. Они нарушают установившийся ход онтогенеза, разрушают целостность организма, т.е. тесную взаимозависимость и взаимообусловленность, скоррелированность частей.
Особенно вредны крупные мутации, которые вызывают значительные изменения в норме реагирования, а вследствие этого и в онтогенезе организмов. Однако крупные мутации не так часты. Наиболее обычны мелкие мутации, которые лишь незначительно изменяют норму реакции, а потому и не влияют отрицательно на онтогенез. Именно поэтому мелкие мутации могут свободно накапливаться в популяции вида. Особенно это относится к таким мутациям, которые совершенно не изменяют нормы реагирования и потому фенотипически трудно или совсем не регистрируются. Подобные мутации, накапливаясь с значительной быстротой, могут дать такой же конечный эффект, как и эффект крупных мутаций; через суммирование они могут привести к нарушениям в онтогенезе, т. е. приобрести относительно вредное значение.
Итак, мутации сами по себе не имеют положительного значения. Если данный вид как жизненная форма хорошо приспособлен к среде, то бесконтрольный мутационный процесс может иметь только одно следствие: он разрушает или, во всяком случае, более или менее нарушает приспособленность вида, снижая его жизнеспособность. Это один из весьма частых эффектов мутаций. Поэтому в большинстве случаев мутанты, например у дрозофилы, полученные в лабораторных условиях, быстро погибают в естественной обстановке. Эти данные уточняют уже давно известное положение, что мутации возникают независимо от того, полезны они или вредны.
На первый взгляд кажется парадоксальным, что мутации, т.е. наследственные изменения, являющиеся, по Дарвину, фактором эволюции, относительно вредны. В самом деле, вид развивается через эволюцию приспособительных изменений. Каким же образом относительно вредные мутации могут служить материалов для приспособлений? Очевидно, если, с одной стороны, мутации дают материал для приспособительной эволюции, а с другой стороны, они относительно вредны, то в таком случае должен существовать какой-то естественный процесс, который превращал бы относительно вредные мутации в полезные изменения, т. е. приспособления. Без наличия такого процесса характеристика мутаций делает совершенно невозможным объяснение эволюции только при помощи теории наследственной изменчивости. Другими словами, очевидно, что эволюция не может быть сведена к наследственной изменчивости. Таков весьма важный вывод из рассмотрения роли мутаций. Несмотря на это, мутации имеют большое эволюционное значение, хотя они и являются лишь элементарным материалом эволюции.
Роль комбинативной изменчивости
Большинство организмов размножаются при посредстве гамет, разноименных по полу и индивидуальности. Так, например, хотя среди растений есть много самоопылителей, но, как оказывается, только немногие являются облигатно (обязательно) самоопыляющимися формами.
У всех остальных самоопылителей почти всегда наблюдается факультативное (временное) перекрестное опыление. Поэтому можно утверждать, что в растительном мире в целом господствует перекрестное опыление. У животных соответственно также господствует не самооплодотворение, а размножение при участи двух полов. Таким образом, фактически при господстве скрещивания неизбежно образование генетических комбинаций. Уже отсюда становится ясной огромная роль комбинаций и комбинативной изменчивости. Еще Ч. Дарвином был установлен факт благотворного воздействия скрещивания на жизнеспособность потомства. Одна из причин этого явления состоит в том, что мутации, как правило, вливаются в популяцию вида в комбинации с их аллеломорфными и другими наследственными качествами. Поэтому сплошь и рядом наблюдается, что относительно вредный эффект мутаций при таком комбинировании их с другими качествами снижается или нейтрализуется. Комбинирование делает потомство более многообразным как генетически, так и фенотипически. И далее, комбинирование стимулирует новые проявления наследственной изменчивости (комбинативной). Сюда относятся и мутации, возникшие на той же почве комбинирования. Основное значение имеет тот факт, что при комбинировании снижается и даже нейтрализуется относительно вредный эффект мутаций.
Итак, значение комбинативной изменчивости таково: 1) стимулирует возникновение новых наследственных изменений; 2) повышает генетическое и фенотипическое многообразие потомства; 3) повышает жизнеспособность потомства и 4) нейтрализует вредное влияние мутаций.
По всем этим причинам комбинации со включенными в них через скрещивание мутациями становятся у двуполых, перекрестно размножающихся форм непосредственным материалом для эволюции. Однако комбинации, так же как и мутации, сами по себе не могут обеспечить эволюционного процесса. Ведь комбинации возникают в природе относительно свободно, в большом количестве. При этом возникают самые различные комбинации, вовсе не обязательно «удачные», в том числе не обязательно жизнеспособные. Напротив, возникают и вредные комбинации, ведущие организм к гибели. В чем же положительная роль комбинаций? Основное значение комбинаций заключается в том, что под влиянием комбинирования при скрещивании комбинативные формы становятся многообразными, биологически неравными. Эта биологически неравная масса индивидуумов данного вида становится тем фактически неисчерпываемым материалом, за счет которого и развертывается внутривидовой эволюционный процесс.
Роль корреляций
Корреляции возникают в ходе индивидуального развития организма. Между отдельными частями и органами устанавливаются взаимозависимые связи. Благодаря наличию корреляций организм становится целостным. Части его нормально функционируют только в их связях друг с другом. Организм в ходе индивидуального развития формируется не как механический «агрегат» частей, которые можно разобрать и сложить, а как целостность. Поэтому и значение корреляций в эволюции прежде всего заключается в том, что при их наличии может эволюировать только целостный организм со всеми его морфофизиологическими «достоинствами» и «недостатками». Положение это было развито И. И. Шмальгаузеном.
Корреляции определяют «пригодность» данной мутации. Поэтому корреляции, как и модификации, становятся «проводниками в жизнь» конкретных мутаций, являясь инструментом, который (конечно, через борьбу за существование) определяет их жизненность.
Итоговые замечания и выводы
При обсуждении роли различных форм изменчивости и значения наследственности почти не затрагивалась дарвиновская теория борьбы за существование и естественного отбора, т. е. была предпринята попытка обсудить вопрос об эволюционном значении мутаций, комбинативной изменчивости, корреляций и модификаций независимо от этой теории. Однако обзор эволюционного значения этих форм изменчивости вне связей с теорией отбора неизбежно неудовлетворителен. Вывод: мутации дают элементарный материал для эволюции; комбинации дают непосредственный материал для эволюции; при обсуждении эволюционной роли модификаций и рассмотрении значения корреляций уже вообще невозможно обойтись без кратких слов относительно естественного отбора. Это происходит потому, что полностью эволюционное значение форм изменчивости и наследственности можно понять только на основе и в свете теории естественного отбора. Доказательства этого:
1) Мутации не обеспечивают эволюционного процесса. Это вытекает из следующего:
- мутации относительно вредны, в то время как эволюционное развитие видов осуществляется через развитие приспособлений, т. е. полезных изменений, которые, следовательно, неравнозначны мутациям. Отсюда ясно, что мутационной теорией объяснить приспособительный характер эволюционного процесса нельзя.
- мутационный процесс протекает в каждом поколении, в то время как вид изменяется и превращается в другой вид на протяжении многих поколений. Значит, нельзя свести эволюционный процесс к мутационному процессу. Не мутации определяют скорость эволюционного процесса.
2) Комбинации не обеспечивают эволюционного процесса. Комбинации возникают стихийно, они в пределах видовых популяций относительно свободны. Поэтому через комбинирование могут возникнуть разные формы — приспособленные и неприспособленные. Однако популяции вида всегда состоят из более или менее приспособленных форм. Значит, не сама комбинация, а что-то другое создает приспособительную эволюцию.
3) Модификации и корреляции не обеспечивают эволюционного процесса. Во-первых, модификации ненаследственны. Поэтому они не могут сами обеспечить закрепление новых особенностей. Во-вторых, модификации могут быть различными — вредными, полезными или индифферентными. Однако в популяции вида, населяющей конкретную стацию (местообитание), модификации носят, как правило, адаптивный характер. Значит, должен существовать особый фактор, который сохраняет только приспособительные модификации. И в этом случае очевидно, что модификации не объясняют приспособительного характера эволюции. То же самое можно сказать и о корреляциях. И они сами не обеспечивают гармоничность формы.
4) Наследственность не объясняет стойкости видовой формы. Как различные формы изменчивости не объясняют поступательного эволюционного процесса, так и фактор наследственности недостаточен для объяснения стойкости видовой формы, стойких признаков конкретных организмов. Значит, кроме генетической преемственности, должна существовать какая-то другая форма преемственности. Эта филогенетическая преемственность основана на том, что признак становится очень стойким не потому, что с ним стойко связано неизменное наследственное основание, а потому, что этот признак надолго сохраняет свое полезное значение. Ясно, что стойкость признака на протяжении длинной истории вида или группы видов определяется естественным отбором, а не предполагаемым постоянством генов.
Вывод
![]() Различные формы изменчивости не могут объяснить поступательный эволюционный процесс, всегда носящий приспособительный характер. Наследственность хотя и может объяснить сходство особей в ряду поколений с родоначальной родительской парой, но недостаточна для объяснения стойкости видовой формы на протяжении жизни вида, измеряемой геологическим временем. Тем более только наследственность не может объяснить стойкости отдельных признаков, существующих у конкретных организмов на протяжении многих миллионов лет.
Различные формы изменчивости не могут объяснить поступательный эволюционный процесс, всегда носящий приспособительный характер. Наследственность хотя и может объяснить сходство особей в ряду поколений с родоначальной родительской парой, но недостаточна для объяснения стойкости видовой формы на протяжении жизни вида, измеряемой геологическим временем. Тем более только наследственность не может объяснить стойкости отдельных признаков, существующих у конкретных организмов на протяжении многих миллионов лет.
Все это показывает, что объяснить эволюционный процесс действием только изменчивости и наследственности практически невозможно. И изменчивость, и наследственность являются необходимыми и обязательными предпосылками процесса эволюции. Поэтому естественно привлечение для объяснения эволюции какого-то другого фактора. Таким фактором оказывается естественный отбор.
Список литературы
1) Веселов Е.А. Дарвинизм. М.: Государственное учебно-педагогическое издательство Министерства просвещения РСФСР, 1960г., 504с.
2) Гурев Г.А. Дарвинизм и его значение. М.: Государственное учебно-педагогическое издательство Министерства просвещения РСФСР, 1959г., 340с.
3) Дарвин Ч. Происхождение видов. М.: Сельхозгиз, 1952г., 484с.
4) Медников Б.М. Дарвинизм в XX веке. М.: Советская Россия, 1975г., 224с.
5) Парамонов А.А. Дарвинизм. М.: Просвещение, 1978г., 336с.
6) Тимирязев К.А. Краткий очерк теории Дарвина. М.: Сельхозгиз, 1953г., 160с.
7) Шмальгаузен И.И. Проблемы дарвинизма. Ленинград: Наука, 1969г., 496с.
Похожие работы
... при их слиянии зиготе хранится наследственная информация о физических, биохимических и физиологических свойствах, с которыми появляется новый человек. Материальной основой наследственности служат нуклеиновые кислоты, а именно ДНК. Но каким же образом генетическая информация передается от родителей к потомству? Как известно, новые клетки появляются в результате деления исходных материнских. Для ...
... , видов и т. д. в результате дивергенции Роль в эволюции Приспособление организмов к условиям среды Материал для естественного отбора Форма изменчивости Определенная (групповая) Неопределенная (индивидуальная), комбинативная Подчиненность закономерности Статистическая закономерность вариационных рядов Закон гомологических рядов наследственной изменчивости ...
... жизни имеет генетический характер, ибо наследственность и изменчивость поддерживают стабильность свойств и способность организмов к изменчивости. Однако генетическая непрерывность жизни связана не только с наследственностью и ее изменчивостью. Она связана также со средой, в которой живут и развиваются организмы. Все организмы живут и размножаются в среде, условия которой не безразличны для них. ...
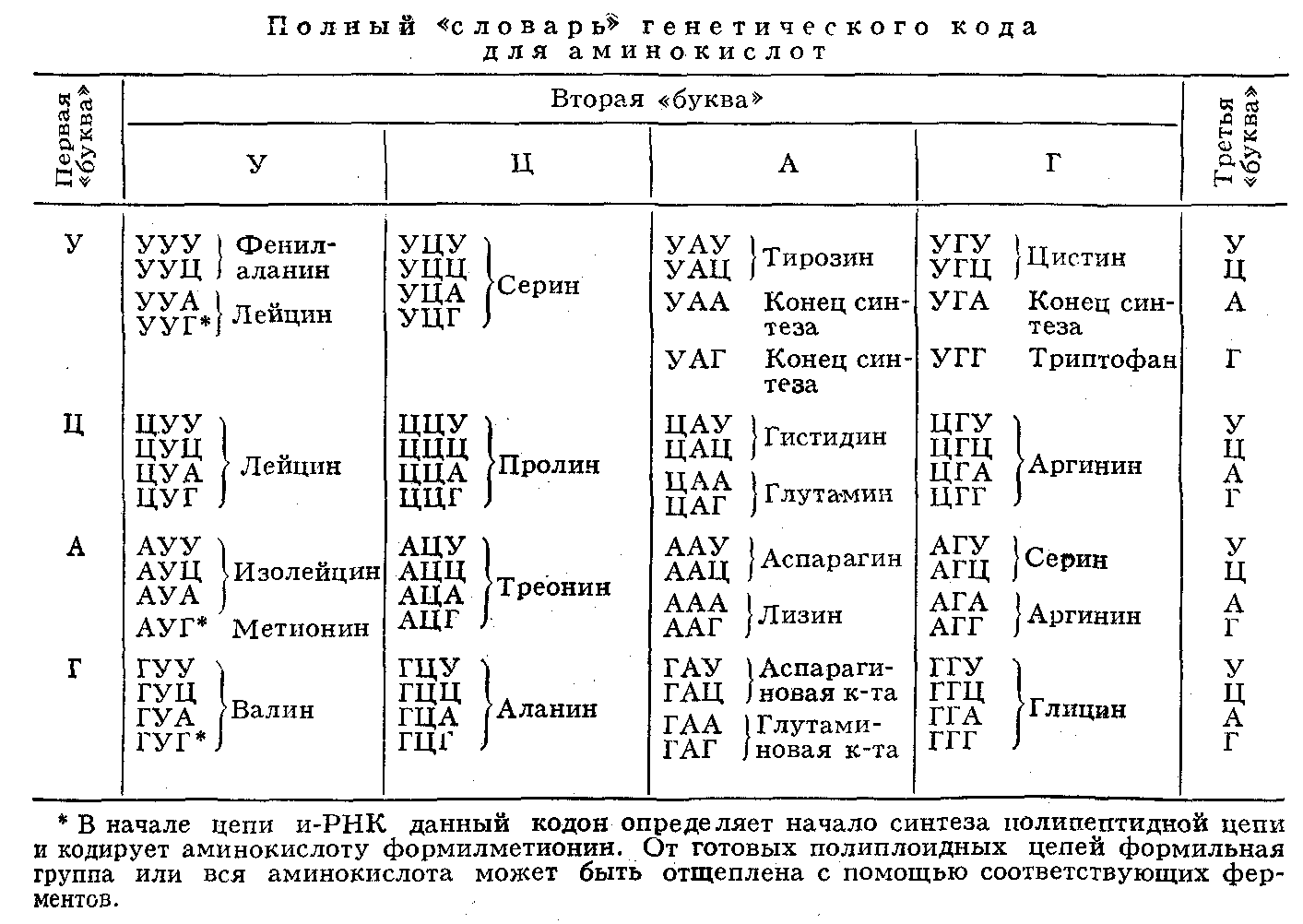
... живые организмы-и удивительное многообразие генов, кодирующих эти белки. В геноме каждого человека есть какие-то области, определяющие его индивидуальность. Некоторые гены человека отличаются от генов крысы всего несколько нуклеотидами-знаками генетического кода. Другие гены у них разные, но одинаковые у двух людей. Изменчивость, связанная с существованием генов , подобных генам группы крови у ...
0 комментариев