Онтогенез – это индивидуальное развитие организма, в ходе которого происходит преобразование его морфофизиологических, физиолого-биохимических и цитогенетических признаков. Онтогенез включает две группы процессов: морфогенез и воспроизведение (репродукцию): в результате морфогенеза формируется репродуктивно зрелая особь. Онтогенез характеризуется устойчивостью – гомеорезом. Гомеорез – это стабилизированный поток событий, который представляет собой процесс реализации генетической программы строения, развития и функционирования организма.
С точки зрения эволюции рассматриваются следующие моменты онтогенеза: эмбриональные адаптации; филэмбриогенезы; автономизация онтогенеза; эмбрионизация онтогенеза.
Основные атрибуты онтогенеза
Исходная запрограммированность процессов. Наличие уникальной неизменной генетической программы развития, сформированной вследствие мейоза и оплодотворения
Необратимость онтогенеза. При реализации генетической программы невозможен возврат к предыдущим стадиям
Углубление специализации: по мере развития уменьшается вероятность смены траектории онтогенеза
Адаптивный характер: поливариантность онтогенеза обеспечивает возможность приспособления к различным условиям
Неравномерность темпов: скорость процессов роста и развития изменяется.
Целостность и преемственность отдельных этапов. Признаки, появляющиеся на более поздних стадиях, базируются на признаках, проявляющихся на ранних стадиях
Наличие цикличности: существует цикличность старения и омоложения
Наличие критических периодов, связанных с выбором пути в узловых точках (точках бифуркации) или с преодолением энергетических порогов.
Основные типы онтогенеза
1. Онтогенез организмов с бесполым размножением и/или при зиготном мейозе (прокариоты и некоторые эукариоты).
2. Онтогенез организмов с чередованием ядерных фаз при споровом мейозе (большинство растений и грибов).
3. Онтогенез организмов с чередованием полового и бесполого размножения без смены ядерных фаз. Метагенез – чередование поколений у Кишечнополостных. Гетерогония – чередование партеногенетического и амфимиктического поколений у червей, некоторых членистоногих и низших хордовых.
4. Онтогенез с наличием личиночных и промежуточных стадий: от первично-личиночного анаморфоза до полного метаморфоза. При недостатке питательных веществ в яйце личиночные стадии позволяют завершить морфогенез, а также в ряде случаев обеспечивают расселение особей.
5. Онтогенез с выпадением отдельных стадий. Утрата личиночных стадий и/или стадий бесполого размножения: пресноводные гидры, олигохеты, большинство брюхоногих моллюсков. Утрата конечных стадий и размножение на ранних этапах онтогенеза: неотения.
Таким образом, существует множество основных типов онтогенеза и еще большее число производных типов. В теории эволюции обычно рассматривается онтогенез на примере цветковых растений и позвоночных животных.
Рассматривая морфологическую сторону эволюции онтогенеза, наиболее существенную для палеонтолога, необходимо прежде всего иметь в виду взаимосвязь между изменениями в индивидуальном развитии и эволюцией взрослого организма. Именно эта проблема составляет, например, суть всего учения о филэмбриогенезах А. Н. Северцова [926, 927, 928]. Хотя такое внимание к итогу развития иногда считают чем-то вроде пережитка геккелевского мышления [163, 540] или, в лучшем случае, допустимым методическим приемом [1661], на деле оно неизбежно, если нас интересует реальный эволюционный процесс. Только во взятом изолированно онтогенетическом цикле взрослая стадия может казаться наименее значимой, поскольку она полностью обусловлена предыдущим развитием, но сама уже не определяет ничего. В эволюционном же аспекте дело представляется совсем иначе. Взрослая стадия — единственная, на которой в норме (по крайней мере, у Metazoa) лежит функция размножения, создающего генетическое разнообразие, т. е. материал эволюционного процесса. От успеха взрослых организмов в борьбе за существование зависит, какого сорта гаметы послужат для создания следующего поколения популяции и, следовательно, с каким феноти-пическим материалом будет в дальнейшем иметь дело отбор. В процессе отбора вместе с неудачными фенотипами элиминируются и реализующие их онтогенезы.
Это означает, что какие бы удачные приспособления ни создавались на промежуточных стадиях развития, они не дают преимуществ, если в итоге онтогенез не обеспечивает осуществления нужного фенотипа. Взрослая стадия как бы диктует свои условия всему онтогенезу. Поэтому столь часто критикуемые слова Э. Геккеля о том, что филогения (понимаемая им как совокупность. взрослых стадий) есть причина онтогении, в действительности имеют глубокий смысл, хотя и не совсем тот, который подразумевался автором биогенетического закона.
То, что изменяются все стадии онтогенеза, ни у кого не вызывает сомнений. Вопрос в другом — как соотносятся их изменения? Эволюирует ли взрослая стадия самостоятельно или под действием одновременных морфологических изменений в раннем развитии, или же обоими путями сразу? Наконец, возможно ли в филогенезе обратное воздействие поздних изменений на ход предшествующих стадий — т. е. такое, которое немыслимо в пределах одного онтогенеза?
В ответе на первый вопрос обнаруживается поразительное противоречие в существующих взглядах на роль онтогенеза в эволюции, объединяющих, по сути, две несовместимые концепции. С одной стороны, признается, что нормальное развитие есть зарегулированный процесс, устойчиво направленный к достижению конечного результата и стремящийся свести на нет все возникающие на этом пути уклонения [1192, 1196, 2439, 2441]. Отсюда следует, что эти вариации не должны непосредственно влиять на облик взрослого организма. Эволюция последнего должна быть связана с формообразовательными уклонениями, относящимися к концу развития, где возможности регуляции снижаются. И в то же самое время господствует убеждение, что изменения ранних или промежуточных стадий могут служить причиной немедленного отклонения (девиации) всего хода онтогенеза, радикально меняющего облик взрослой формы у потомков. Обычно этого противоречия даже не замечают. Обе концепции существуют давно и одинаково берут начало в классической морфологии, но лишь первая из них,. как будет показано, может быть согласована с требованиями эволюционной теории.
Первооснову для нее создали два эмпирических обобщения К. М. Бэра [1291]. Одно из них касается уменьшения эмбриональной изменчивости в последовательных стадиях (говорящее, по-мнению К. М. Бэра, о присутствии высшего целеполагающего контроля, регулирующего развитие). Второе составляет известный закон Бэра, или закон зародышевого сходства. В рамках эволюционной теории, как указывал уже Ч. Дарвин [309], это явление означает, что отбор стремится изменять организмы главным образом в позднем возрасте. В дальнейшем эти представления получили поддержку со стороны механики развития, экспериментально показавшей способность морфогенетических процессов к саморегуляции, в результате чего взрослый организм оказывается более устойчивым, чем способ его осуществления (в онтогенезе или при регенерации). Это обобщение, известное как «правило Ру» [921, 2184], или принцип эквифинальности [1524], подтвердило высокую консервативность ранних стадий развития, показывающую, что эволюция взрослой организации должна происходить преимущественно путем изменений в позднем онтогенезе [2185].
Однако главным источником таких представлений послужил основной биогенетический закон Э. Геккеля [1687], представляющий собой переложение на эволюционный язык старой идеи о параллелизме индивидуального развития и «лестницы существ». Выполнение этого закона (т. е. краткое повторение филогенеза в онтогенезе) невозможно представить себе иначе, как'на основе конечных изменений развития.
Поскольку Э. Геккеля интересовала в онтогенезе не столько его эволюция, сколько получение метода для филогенетических реконструкций, то о причинах такого хода изменений мы не находим у него специальных разъяснений, кроме многочисленных и мало что говорящих «законов» наследственности и приспособления. Тем не менее в биогенетическом законе содержится вполне целостная теория эволюции онтогенеза, подразумевающая два положения:
а) взрослая стадия эволюирует посредством прибавления к онтогенезу новых конечных стадий, что обеспечивает рекапитуляцию в онтогенезе облика взрослых предков (палингенез);
б) промежуточные стадии имеют собственную адаптивную эволюцию, искажающую онтогенетическую запись преобразований взрослой стадии (ценогенез).
Биогенетический закон обычно связывают с механоламарк-истскими взглядами Э. Геккеля [359, 565, 1196], иногда утверждая даже, что он был опровергнут самим открытием менделевских факторов и мутационного процесса [1661, 1892]. Однако в действительности вопрос о природе наследственности не играет решающей роли в оценке закона [1822]. В самом деле, независимо от того, связывается ли появление эволюционных новшеств с поздними соматическими изменениями, передаваемыми каким-то образом через половые клетки (ламаркистская концепция) или же с непосредственным изменением самих этих клеток (концепция наследственных факторов), в любом случае новый цикл развития неизбежно должен начинаться с преобразований зиготы, т. е. отличаться от родительского с самого начала.
Э. Геккель ясно понимал это обстоятельство и отнюдь не видел в нем препятствия для биогенетического закона, указывая, что изменения, возникшие в определенный момент развития и передаваемые через родительские гаметы, обнаруживают себя впервые у потомков на той же самой стадии («закон одновременного наследования» [1687]). В этом он прямо следовал Ч. Дарвину [310], сформулировавшему принцип «наследования в соответствующем возрасте» и подчеркивавшему, что появление наследственного уклонения в гаметах и его видимое проявление в индивидуальном развитии у потомства суть разные вещи. Поэтому неудивительно, что признание детерминантов или генов в качестве носителей наследственности не мешало многим исследователям признавать биогенетический закон [1380, 1692, 2472] или, по крайней мере, связь эволюционных изменений с поздними стадиями [503, 709].
Противоположный взгляд на эволюцию онтогенеза, допускающий изменение взрослых организмов путем уклонения на ранних стадиях развития, ведет свое начало от Э. Жоффруа Сент-Илера [2198] и Ф. Мюллера [719]. У них и у современных авторов он чаще всего имеет одно и то же обоснование — сравнение нормальных онтогенезов (или морфогенезов гомологических органов) и отождествление наблюдаемой конечной онтогенетической разницы (зародышевой дивергенции Бэра) с реальным эволюционным процессом. Если, например, морфогенезы гомологичных органов А и Б у двух разных форм совпадают до стадии х, а затем расходятся, то утверждают, что орган Б возник из А путем девиации на стадии х [2141]. Методологическая несостоятельность этой аргументации будет нами показана. Представления о возможности или неизбежности такого пути эволюции никогда не составляли в отличие от геккелевских какой-либо целостной теории. Тем не менее уже к началу XX в. они стали преобладающими.
Одной из причин этого было накопление фактов сравнительной эмбриологии, не согласующихся, по мнению исследователей, с биогенетическим законом. Признание эволюции путем ранних уклонений развития устраняло тезис о первичной неизбежности рекапитуляции, правда, ценой того, что само их существование превратилось в загадку [565, 1869]. Другой момент в критике геккелевских взглядов был связан с растущим убеждением эмбриологов, что подлинное объяснение хода индивидуального развития следует искать в изучении его непосредственных причинных факторов, а не в конструировании умозрительных исторических принципов, подменяющих опытное знание. Этот подход и привел к появлению экспериментальной (каузальной) эмбриологии, которая выявила чрезвычайное разнообразие и специфичность структуры зародышевых клеток у различных организмов. Последнее с очевидностью показывало, что зигота является таким же продуктом эволюции, как и взрослый организм, и что начальная стадия развития высшего организма не есть повторение его одноклеточного предка. «Яйцо курицы не более соответствует начальному звену филогенетической цепи, чем сама курица» [1733]. Все это привело к убеждению, что эволюция происходит путем изменения всего онтогенетического цикла снизу доверху в каждом очередном поколении, а не за счет прибавления конечных стадий [1448, 1763, 2259]. Онтогения, таким образом, творит филогению, а не повторяет ее [1623]. При этом одни авторы имели в виду только постепенные преобразования, другие — возможность резких уклонений, скачкообразно меняющих взрослую форму.
В рамках этих взглядов исчезала, однако, всякая теоретическая возможность появления параллелей между онто- и филогенезом вопреки, казалось бы, очевидным фактам их существования. Это затруднение пытались обойти с помощью утверждения, что такие параллели имеют чисто морфогенетические, а не исторические причины, т. е. что анцестральная взрослая стадия может удерживаться в онтогенезе потомка лишь постольку, поскольку она составляет необходимую базу для реализации последующих стадий [1623]. Другое объяснение состояло в том, что большинство подобных примеров, используемых биогенетическим законом, в действительности относится не к повторению облика взрослых низших форм в онтогенезе высших, а просто к сохранению общих стадий развития у тех и у других, т. е. к проявлениям зародышевого сходства, на что указывал еще К. М. Бэр [1623, 1767, 1970 и др.]. Этот довод, постоянно повторяемый и позднее, например [1317, 1319], обычно считают решающим опровержением геккелевской модели эволюции. Но, как ни странно, при этом не замечают, что при перестройке онтогенеза каждый раз с начальной стадии сохранение зародышевого сходства становится таким же невероятным, как и появление рекапитуляции!
В целом, однако, позиция позднейших исследователей по отношению к биогенетическому закону выглядит более умеренной и допускает возможности разных путей онтогенеза. Соответственно выдвигаются различные классификации этих путей, или модусов [565, 926, 928, 1317, 1319, 1604, 2141, 2124], в которых геккелевский способ эволюции путем поздних надставок занимает то или иное место — от сравнительно большого, например у А. Н. Северцова, до ничтожно малого у Г. де Вира [1317]. Почему онтогенез меняется в разных случаях по-разному, этими взглядами не объясняется (если не считать обычного утверждения, что ранние изменения—это путь к созданию крупных систематических групп). А. Н. Северцов [927] прямо указывает, что ответ на этот вопрос вообще не входит в его задачу, которая состоит лишь в том, чтобы выяснить, как эволюция может идти. Он признает, что выдвигаемые им представления о филэмбриогенезах не зависят от принятия той или иной эволюционной теории.
Само разнообразие выделяемых модусов нередко приводит их авторов к выводу, что все логически мыслимые пути изменений индивидуального развития действительно реализуются в эволюции [2141]. Это равносильно признанию, что в преобразованиях онтогенеза нет сколько-нибудь общих закономерностей. Такой малоутешительный итог столетнего изучения проблемы после Э. Геккеля, в общем, неудивителен в условиях, когда оно основывалось на сравнении одних лишь нормальных онтогенезов, вне связи с анализом механизмов, реализующих материал эволюции — онтогенетическую изменчивость.
Очередным подтверждением этого служит работа С. Гулда [1661] о соотношении онто- и филогенеза, представляющая собой наиболее обширное исследование на эту тему за последние десятилетия. Методология здесь все та же — выделяются эволюционные модусы и оценивается, насколько они отражают ход филогенеза. Попытка связать их с двумя типами адаптивных стратегий (г и 7'С-отбором) не меняет традиционного характера подхода к проблеме. Эволюция онтогенеза остается суммой изолированных процессов, каждый из которых идет в своих особых условиях. Вопрос об их общих первопричинах по-прежнему остается в стороне.
ОНТОГЕНЕЗ И СТАБИЛИЗИРУЮЩИЙ ОТБОР
Морфологическая эволюция онтогенеза не может быть понята, исходя из одних лишь ее адаптивных результатов, т. е. из простого сопоставления «готовых» типов нормального развития. В ее основе лежит процесс эволюции формообразовательных механизмов, преобразующий типичную взрослую организацию (адаптивную норму [1184]), а вместе с ней и стандартный путь ее осуществления в онтогенезе. Каждое элементарное изменение взрослой нормы связано с отбором одного из ее уклонений, т. е. с сохранением любых индивидуальных вариантов развития, реализующих данное выгодное уклонение. Этот отбор варьирующих индивидуальных циклов по принципу однозначности их итога неминуемо должен перестраивать типичный ход развития в целом (см. 7.3), и главная задача эволюционной теории состоит в уяснении закономерностей этой морфогенетической перестройки. Морфологическая эволюция онтогенеза является лишь ее внешним выражением.
Основа для решения этой проблемы содержится в теории стабилизирующего отбора, или эпигенетической концепции эволюции (см. 7.3), согласно которой создание элементарного адаптивного изменения выражается в росте устойчивости онтогенетического осуществления исходной для него вариации.
В свете этих взглядов помехоустойчивость типичного развития, вскрываемая экспериментальной эмбриологией, рассматривается как результат отбора на максимальную защиту нормального формообразования. Механизмы этой защиты (саморегуляции) наиболее хорошо изучены для «регуляционных» онтогенезов (позвоночные, иглокожие и т. д.), где развитие на ранних стадиях идет путем индукционного взаимодействия зачатков, определяющего направление их дальнейшей дифференцировки. Устойчивый ход развития поддерживается за счет широкого раздвигания порогов нормального реагирования зачатков по самым различным параметрам: количеству и активности метаболитов, протяженности периодов компетенции (способности к нормальному взаимодействию) и т. д. Поэтому небольшие колебания в этих показателях, вызываемые генетическими или внешними нарушениями, в достаточно широких пределах забуфериваются и не влияют на дальнейший ход развития. Сильные же воздействия, необратимо нарушающие развитие, ведут либо к гибели, либо к дефинитивным аномалиям, снижающим жизнеспособность.
Такая же устойчивость характерна и для онтогенезов «мозаичного» типа (моллюски, членистоногие и др.), где органогенезы протекают в небольших группах клеток, и регуляция, очевидно, имеет место внутри них. Зачатки здесь в принципе так же муль-типотентны, как и в «регуляционных» онтогенезах (например, зачаток глаза насекомого способен развиваться в антенну или конечность), но нормальная детерминация осуществляется столь надежно, что на первый взгляд кажется единственно возможной. Между двумя типами развития нет четкой грани и в других отношениях: «мозаичные» фазы имеют место в «регуляционных» онтогенезах и наоборот. В целом, благодаря саморегуляции, нормальный онтогенез представляет собой канализированную последовательность событий (креод, или, буквально, «необходимый путь» [2441]), стремящуюся выправить все помехи на пути к осуществлению стандартной организации.
Рассматриваемая нами проблема соотношения эволюции взрослого организма и его онтогенеза вначале должна быть поставлена несколько иначе — каким образом становление новых взрослых особенностей связано с перестройкой их формообразовательных механизмов?
Согласно эпигенетической концепции [1178, 1180, 1181, 1192, 1196], в главных чертах эта связь сводится к следующему. Элементарное эволюционное изменение создается отбором как модификация прежней фенотипической нормы (см. рис. 26,. 27, см. 7.3), осуществляемая лишь в тех условиях, на фоне которых она имеет адаптивное преимущество. Это — примитивное (зависимое) формообразование, в котором среда выступает прямо или косвенно (через посредство функции) как детерминирующий фактор развития. По мере того как новый признак приобретает значение оптимальной адаптации для любого варианта нормальных условий, его развитие становится все более независимым от специфических внешних стимулов, благодаря установлению регуляторных взаимосвязей с другими формообразовательными процессами, составляющими нормальный онтогенез. Реализация адаптивного изменения становится, таким образом, неотъемлемой составной частью осуществления фенотипической нормы как целого. В итоге становление новой элементарной адаптации есть процесс замены внешних факторов ее развития внутренними.
Идея о таком ходе эволюционных изменений принадлежит, как известно, неоламаркизму, но лишь теория стабилизирующего отбора впервые дала ей рациональное обоснование. Адаптивная модификация не возникает одноактно, а создается путем отбора среди элементарных индивидуальных реакций (морфозов), представляющих собой видоспецифичные уклонения нормального развития (см. 7.3). Ее последующее фиксирование—результат селективного преобразования частного варианта нормы в безусловную норму, а не выражение «прямого унаследования». Признаки вообще делятся не на «наследственные» и «приобретенные», а лишь на устойчивые к внешним изменениям и зависимые от них [1194, 1196, 2501]. Унаследование новой особенности не означает ничего иного, кроме стабилизации ее онтогенетического осуществления [1180, 1181].
Прямым экспериментальным подтверждением описанного хода эволюционных преобразований являются опыты К. X. Уоддингтона [2441] по генетической ассимиляции морфозов у дрозофилы, в которых индуцированные внешними воздействиями изменения в ходе отбора превращались в стойкие наследственные признаки (подробнее см. 7.3). На такой путь создания адаптации указывает также и множество наблюдений сравнительного характера. Прежде всего это хорошо известные факты параллелизма между адаптивными модификациями у отдельных видов или рас и устойчивыми признаками у их ближайших родственников, что многократно отмечалось для растений, позвоночных, насекомых и т. д. [405, 1192, 1196]. Естественно предполагать, что мы наблюдаем здесь как бы последовательные стадии стабилизации этих признаков, тем более, что указанные модификации у первых («пластичных») видов обычно возникают при воздействии условий, свойственных в норме для вторых («стабильных»). Другие подобные доказательства дают эмбриология и палеонтология. Например, у позвоночных в позднем развитии очень велика формообразую-щая роль функции, в связи с чем еще В. Ру выделял в их онтогенезе дофункциональный (автономный) и функциональный периоды. Сравнение показывает, что граница между ними сдвигается в эволюции за счет нарастания автономности развития у более высокоорганизованных форм. Так, у водных хвостатых амфибий образование легочных альвеол обусловлено дыханием; у более наземных жаб оно индуцируется гормонально и лишь в конечных стадиях дифференцировки зависит от функции; наконец, у амниот оно протекает вполне автономно в раннем развитии [632].
Во многих случаях устойчивого формообразования первичная зависимость развития от внешних факторов самоочевидна. Например, образование мозолей у млекопитающих и бегающих птиц есть обычная адаптивная реакция кожи на трение. Однако в участках, подвергающихся трению постоянно (например, подошвы конечностей), утолщение эпидермиса наступает еще в эмбриональный период [1196]. Стертая поверхность зубов у дюгоня явно возникла первично на основе жевательных движений, однако в онтогенезе она развивается вне связи с жеванием, путем эмбриональной резорбции поверхности зубов [1317]. Еще один пример—развитие вдавленности на внутренней стороне оборотов раковины наутилоидей. Первоначально она возникла как пассивный результат плотного смыкания оборотов, так что у девонских форм ее еще не было на ранних стадиях развития раковины, предшествующих смыканию. Но уже у некоторых карбоновых видов, как и у современных, вогнутость появляется независимо от механического воздействия — на стадии несвернутой раковины [1776].
Похожие работы


... системы адаптации экоморфы достаточно очевидны, т. е. особенности экоморф практически целиком определяются развитием адаптации организменного уровня. Среди рассмотренных аспектов классификации надорганизменных живых систем наиболее существенным представляется деление их на гомогенные и гетерогенные, поскольку главным образом именно оно определяет как общие возможности интегративной эволюции ...

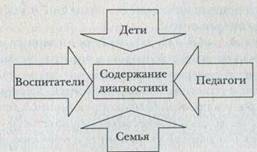
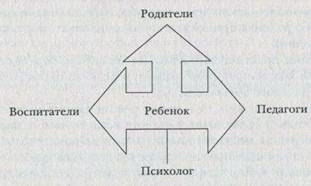
... учреждения. 9. Сформулируйте особенности организации и проведения психокоррекции в условиях дошкольного учреждения. 10. Выделите направления психопрофилактической деятельности детского практического психолога. 11. Сформулируйте особенности организации и проведения психопрофилактики в условиях дошкольного учреждения. 12. Определите содержание и направление психологического просвещения в ...
... Е.И. Жизненная перспектива и профессиональное самоопределение. – Киев, 1988. Ковалева Олеся Ивановна Ставропольский государственный университет, г.Ставрополь КООНТОГЕНЕЗ ЛИЧНОСТИ ПОДРОСТКА В СРЕДЕ ОБЩЕОБРАЗОВАТЕЛЬНОЙ ШКОЛЫ Коонтогенез личности - взаимосогласованное развитие системы человеческого организма с окружающим его миром в опосредованно общей структуре связей при сохранении ...
... в некое квазистабильное состояние. И более чем вероятно, что в этом новом состоянии параметры биосферы окажутся неподходящими для жизни человека, а может быть, и жизни вообще. 3. ОСНОВНЫЕ УРОВНИ ИЕРАРХИИ БИОЛОГИЧЕСКИХ СИСТЕМ Органический мир представляет собой единое целое, так как составляет систему взаимосвязанных частей (существование одних организмов зависит от других), и в то же время ...
0 комментариев