Клетка – сложное образование, в ней обнаружены различные микроскопические и субмикроскопические структуры, обладающие высокой динамичностью и способностью закономерно изменяться соответственно изменениям условий существования организма и приспосабливаться к ним.
Наиболее важными структурами являются ядро и цитоплазма, которые тесно связаны между собой и не могут существовать друг без друга, однако строение и функции их неодинаковы.
Структурные элементы клетки можно разделить на три большие функциональные группы:
1) органеллы, которые катализируют превращение энергии, – митсхондрии и хлоропласта;
2) органеллы, которые катализируют репликацию белков, – рибосомы, полирибосомы;
3) клеточные гранулы и другие образования, которые принимают участие в синтетических реакциях, обмене веществ (сферосомы, цитосомы, элайопласты, транслосомы, осмиофильные гранулы, аппарат Гольджи и т.д.). Электронно-микроскопическими исследованиями установлено, что растительная клетка состоит из большого количества мембранных систем. Многие из субклеточных компонентов имеют мембранное строение. Клеточные мембраны состоят из динамических структур двух типов: мембраны, которые быстро изменяются, однако медленнее, чем мембраны эндоплазматической сети, и мембраны, циркулирующие между определенными внутриклеточными компонентами. Мембраны образуются из закрытых пузырьков (везикул), которые циркулируют в клетке.
Ядро – главный управляющий органоид клетки. Оно покрыто тонкой двухслойной мембраной с порами для соединения ядра с цитоплазмой. Ядро обычно имеет шарообразную форму, но в некоторых случаях – палочкообразную, лопастную и даже сетчатую. В ядре хорошо видна еще меньшая по размеру структура – ядрышко, в котором накапливается и, по-видимому, синтезируется РНК, которая затем переносится в цитоплазму, где она становится основной структурной единицей рибосом. В ядрах содержится большое количество ферментов, обеспечивающих освобождение энергии, ее трансформацию и осуществление многочисленных синтезов. Таким образом, энергетически ядра являются полностью независимыми органоидами клетки, состав их ферментов обеспечивает образование аденозинтрифосфорной кислоты (АТФ) в процессе гликолиза и свидетельствует о высокой метаболической активности.
Химическими компонентами ядра являются в основном белки и нуклеиновые кислоты.
Основные (щелочные) белки – это протамины и гистоны, которые содержат много основных аминокислот: лизина, гистидина и особенно аргинина. Протамины и гистоны с нуклеиновыми кислотами образуют нуклеопротеиды. Другие белки могут содержаться в ядре в виде самостоятельной фазы. Ядрышко состоит из больших гранул, которые по размеру близки к рибосомам (диаметр 15 нм) и содержат большое количество РНК. Основное вещество ядра называется нуклеоплазмой. Ядро содержит хромосомы, которые являются носителями наследственности. Хромосомы имеют хроматиновые структуры, основным компонентом которых является ДНК, а также РНК.
Цитоплазма – пронизана системой мембран, которые отходят от ядерной оболочки и соединяются с внешней мембраной клетки. Эти внутренние клеточные мембраны, образующие густо переплетенную мембрану с многочисленными канальцами и полостями, называют эндоплазм этической сетью. Функциональное значение последней разнообразно. Она принимает участие в синтезе и перемещении ассимилятов. Мембраны эндоплазматической сети пронизывают и объединяют в единое целое клетку, а также служат своеобразными путями перемещения ассимиляторов и передачи раздражения от клетки к клетке, которая осуществляется последовательной перезарядкой поверхности мембран. Химически клеточные мембраны гетерогенны и состоят из протеидов (80%, из них 25% приходится на белки-ферменты) и липидов (20%), среди которых много фосфолипидов.
Митохондрии (от греч. митос – нить, хондрион – гранулы) – органеллы шарообразной формы, диаметром 0,5 мкм и длиной 2 мкм. Это нестойкие структуры, в липофильных жидкостях они разрушаются, а в воде набухают; имеют двойную оболочку, состоящую из внутренней и внешней мембран. Между мембранами есть просвет (10 нм), заполненный сывороткой. Внутренняя мембрана митохондрий образует кристы, или трубочки. К внутренней мембране со стороны стромы присоединены с помощью «стебельков», или ножек, частицы, которые получили название оксисом, потому что они содержат окислительные ферменты. Внутреннее пространство митохондрий заполнено матриксом, или хондриоплазмой, – вязким раствором, содержащим ферменты.
Митохондрии состоят из белка и липидов, среди которых половина приходится на фосфолипиды. Неотъемлемыми компонентами митохондрий являются дезоксирибонуклеиновая кислота (ДНК) и все типы РНК. Находящаяся в матриксе митохондрий ДНК в виде нитей способна к независимой от ДНК ядра репликации. В митохондриях обнаружены специфические рибосомы, которые обеспечивают автономный синтез некоторых белков. Так, митохондрии проростков гороха содержат (в%): белка – 30–40, РНК – 0,5–1,0 и фосфолипидов – 30. В митохондриях сосредоточены ферменты цикла трикарбоновых кислот, флавопротеиды и цитохромы. Митохондрии, дыхательные центры клетки обладают следующими функциями:
1) осуществляют окислительные реакции, являющиеся источником электронов;
2) переносят электроны по цепи компонентов, синтезирующих АТФ;
3) катализируют синтетические реакции, идущие с использованием энергии АТФ;
4) регулируют биохимические процессы в цитоплазме.
Пластиды – имеют двойную мембранную оболочку, внутри которой находится гранулярное вещество, называемое стромой. В начале развития строма пластид имеет гранулярное строение. Структура хлоропласта формируется в несколько этапов. Первичная дифференциация пластиды начинается с инвагинаций внутренней мембраны до образования проламеллярного тела (без света). Второй этап связан с образованием ламеллярногранулярного строения, биосинтезом и накоплением хлорофилла. В хлоропластах высших растений образуются граны, которые состоят из серии ламелл или двойных мембран. Каждая двойная мембрана образует закрытый мешочек, или сумку, которая называется тилакоидом. Ламеллы состоят из белков и липидов. Химический анализ ламелл. выделенных из хлопопластов шпината, показал, что они на 52% состоят из белка и на 48% из липидной фракции, которая включает хлорофилл а и в, каротиноиды (ксантофиллы и каротины), пластохинон, витамин К1, фосфолипиды (галактозилглицериды, фосфоглицериды), сульфолипиды.
Рибосомы – это рибонуклеопротеидные частицы сферической формы, диаметром 15–35 мм. Они состоят приблизительно из одинакового количества структурного белка и высокополимерной РНК. Комплексы из пяти и более рибосом называются полирибосомами или полисомами. Каждая рибосома состоит из двух субъединиц с различными коэффициентами седиментации, которые агрегируют в рибосому с помощью ионов магния. Слипаясь по две, они образуют димеры.
Рибосомы очень пористые и отличаются высокой степенью гидратации, выполняя чрезвычайно важные функции в обмене веществ – это центры биосинтеза белка в клетке. Функции рибосом в белковом синтезе заключаются в том, что они осуществляют процесс, в котором активированные аминокислоты конденсируются, образуя полипептидную цепь.
Сферосомы – субмикроскопические компактные частицы цитоплазмы диаметром 0,4–0,8 мкм, содержат белковую строму и цитохромоксидазу, ферментативно активные, богаты жирами. Сферосомы, по-видимому, осуществляют биосинтез жиров, а именно последний его этап – переэтерифнкацию глицерофосфата путем обмена между фосфорной кислотой и жирными кислотами. Таким образом, Сферосомы считают специализированными органеллами, функция которых – биосинтез жиров.
Лизосомы по структуре и химическому составу близки к сферосомам, но богаче ферментами – в них найдены ферменты нуклеазы, фосфатазы, протеазы и т.д. Вполне вероятно, что лизосомы переваривают макромолекулярные продукты, поглощенные путем пиноцитоза. Они принимают участие и в автолизе клетки.
Цитосомы – мелкие гранулы, которые находятся в тесном контакте с мембранами эндоплазматической сети, чем и отличаются от свободнолежащих сферосом и лизосом.
Транслосомы – толстостенные гранулы, функция которых, по-видимому, заключается в накоплении продуктов метаболизма фенольных производных и их транспортировке в вакуоль.
Наконец, так называемые диктиосомы, или тельца Гольджи, которые могут состоять из отдельных пластинок, палочек и чешуек, разбросанных по всей цитоплазме клетки. Возможно, что аппарат Гольджи принимает участие в управлении общим ходом физиологических процессов, в образовании вакуолей и клеточных оболочек.
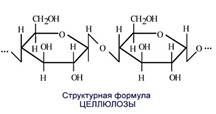
Клеточная оболочка состоит из клетчатки, или целлюлозы (С6Н10О6)n, – полисахарида, который гидролизуется до глюкозы; клетчатка является главным веществом хлорофиллоносных растений и по абсолютному количеству занимает первое место среди всех органических веществ на земной поверхности.
Клеточная оболочка начинает развиваться с образования клеточной пластинки в анафазе митоза. Это происходит сразу после деления ядра. Целлюлоза в виде микрофибрилл образует каркас. Микрофибриллы – эластичный строительный элемент клеточной оболочки (стенки). Диаметр микрофибриллы составляет 10–30 нм, длина несколько микрометров.
Основная функция вакуоли – поддержание гомеостаза клетки. В клеточном соке вакуоли в растворенном состоянии содержатся соли, сахаристые вещества, белки, аминокислоты, органические кислоты, липоиды, а также пигменты, которые относятся главным образом к группе флавоноидов. Так, пигменты антоцианы придают лепесткам цветков и другим частям растения красную, фиолетовую, синюю окраску. В корнях столовой свеклы красный цвет обусловливается присутствием в клеточном соке бетанина – гликозида β-цианина (азотсодержащего аналога антоцианина).
2. Макроэргические соединения (сахарофосфаты, АТФ, УДФ и др.). Их роль в метаболизме клеткиМакроэргические соединения – органические соединения живых клеток, содержащие богатые энергией, или макроэргические связи. Эти соединения образуются в результате фото- и хемосинтеза и биологического окисления. К Макроэргическим соединениям относятся аденозинтрифосфорная кислота (АТФ) и другие вещества, распад которых сопровождается освобождением свободной энергии, используемой клетками для осуществления биосинтеза необходимых веществ или различных видов работы.
АТФ (аденозинтрифосфорная кислота) – нуклеотид, образованный аденозином и тремя остатками фосфорной кислоты. Во всех живых организмах выполняет роль универсального аккумулятора и переносчика энергии. Под действием специальных ферментов концевые фосфатные группы отщепляются с освобождением энергии, которая идет на синтетические и другие процессы жизнедеятельности.
Аденозиндифосфат (АДФ) – нуклеотид, образованный аденозоном и двумя остатками фосфорной кислоты. Участвует в энергетическом обмене живых организмов.
АДФ получает энергию путем дефосфорилирование фосфоэнолпировиноградной кислоты под действием фермента трансфосфорилазы, которая переносит макроэргическую связь с кислоты на АДФ.
Уридиндифосфорная кислота (УДФ) и ее производные принимают участие во взаимопревращении углеводов.
При биосинтезе гликозидной связи используется уридиндифосфатглюкоза (УДФГ), образующаяся из глюкозы‑1‑фосфата и уридинтрифосфата (УИФ). Если УДФГ передает глюкозу фруктозе, то образуется сахароза, а если цепочке декстрина – полисахарид. Аналогично образуются гликозиды, гликопротеиды и др.
Взаимопревращение моносахаридов проходит через фосфорные эфиры сахаров или их уридиндифосфатпроизводные (УДФ-производные). УДФ-производные сахаров представляют собой тот или иной сахар, соединенный через два остатка фосфорной кислоты с уридином.
Сахарофосфаты являются источником фосфорного питания растений. Могут быть соли орто-, мета- и пирофосфорной кислоты и органические фосфаты. Лучшие из них – водорастворимые калиевые, натриевые, аммониевые, кальциевые и магниевые соли фосфорной кислоты.
3. Условия, необходимые растению для нормального водообмена. Физиологические особенности засухоустойчивых растенийДля растений необходимо, чтобы осадки относительно равномерно распределялись в период активного роста.
Под засухой понимают режим погоды, характеризующийся отсутствием осадков, повышением температуры воздуха и падением относительной влажности воздуха. Различают два типа засухи – атмосферную и почвенную. Следует подчеркнуть, что засуха, за очень редким исключением, всегда начинается как атмосферная засуха, переходя затем в почвенную. В то же время атмосферная засуха может быть и при отсутствии почвенной, когда она проявляется в виде горячего ветра-суховея. Вред от суховея наблюдается и тогда, когда в почве еще имеются значительные запасы воды.
Если в почве имеется недостаточное количество доступной для растения воды, то отрицательное влияние перегрева становится особенно сильным. Перегрев вызывает повреждение растения, называемое запалом. Запал обнаруживается через некоторое время в виде различно окрашенных некротических пятен на листьях. На пшенице появляются желтые пятна, на овсе – красные, у большинства растений – коричневые.
Встречается и другой вид повреждений от атмосферной засухи – захват. Он наблюдается реже, чем запал, и проявляется в том случае, когда при сравнительно не очень высоких температурах наблюдаются сильный ветер и большая сухость воздуха. При этом листья просто высыхают, сохраняя зеленую окраску.
Недостаток воды в тканях растений (водный дефицит) может возникнуть в жаркую солнечную погоду к середине дня, при этом увеличивается сосущая сила листьев, что активирует поступление воды из почвы. Растение регулирует уровень водного дефицита открытием или закрытием устьиц. В этот период происходит временное завядание листьев. Обычно в вечерние и утренние часы это явление устраняется.
Отсутствие в почве доступной для растения воды приводит к глубокому завяданию. Это завядание чаще всего приводит к гибели растения. Характерным признаком устойчивого водного дефицита является сохранение его в тканях утром, прекращение выделения пасоки из срезанного стебля. Действие засухи приводит в первую очередь к уменьшению в клетках свободной воды, что нарушает гидратные оболочки белков цитоплазмы и сказывается на функции белков-ферментов.
Приспособление растений к засухе. Известно, что растения неодинаково реагируют на перегрев и обезвоживание в разные периоды онтогенеза. У каждого вида в онтогенезе имеется такой период, когда недостаток воды резко сказывается на течении всех физиологических процессов, этот период называется критическим периодом. Из этого, однако, не следует, что остальные периоды своего развития растение не нуждается в воде и не страдает от ее недостатка.
Установлено, что вязкость цитоплазмы, ее эластичность в критический период резко падают, что и является одним из условий высокой чувствительности растений к перегреву и обезвоживанию в этот период развития. В критический период происходят интенсивные ростовые процессы и образование новых органов – цветков.
Ф.Д. Сказкин считал, что высшие растения в своем онтогенезе повторяют черты далекого прошлого и во время формирования половых органов и оплодотворения нуждаются в повышенной обводненности тканей. Сказкин считает, что началом критического периода следует считать момент появления материнских клеток пыльцы в археспориальной ткани пыльников, а концом данного периода – оплодотворение.
Засухоустойчивые сорта при значительном водном дефиците характеризуются синтетической направленностью в работе своих ферментов, тогда как у менее засухоустойчивых преобладает гидролитическая направленность. Засухоустойчивые сорта озимой пшеницы характеризуются большим содержанием связанной воды, которая трудно обменивается во время засухи, повышенной концентрацией клеточного сока в период цветения и налива зерна, высшим порогом коагуляции белков, более интенсивным накоплением сухого вещества зерна, более стойкой к неблагоприятным условиям пигментной системой. Этим и характеризуется физиологическая природа засухоустойчивости растений.
4. Нарушение водообмена. Его причины и последствияНарушение водообмена у растений происходит при недостаточном водоснабжении, то есть, при наступлении засухи.
Засуха – неблагоприятное сочетание метеорологических условий, которые не обеспечивают потребностей растения в воде. Засуха бывает почвенная и атмосферная. Причины почвенной засухи – отсутствие дождя на протяжении длительного времени, испарение с поверхности почвы и транспирация, что приводит к высушиванию корнеобитаемого слоя почвы. Последнюю высушивают также сильные ветры. При почвенной засухе влажность воздуха бывает пониженной. Атмосферная засуха вызывается массами сухого и нагретого воздуха (суховея). Засуха, как почвенная, так и атмосферная, задерживает рост растений, уменьшает их листовую поверхность, в результате чего урожай снижается.
Влияние почвенной (обезвоживание) и атмосферной (перегрев) засух проявляется многими признаками. Под влиянием обезвоживания нарушается синтетическая способность растений, наблюдается распад белков и изменяется коллоидно-химическое состояние цитоплазмы, снижается количество накопленного растением органического вещества из-за задержки роста листьев и уменьшается их рабочая поверхность. Особенно чувствительны к засухе листья, находящиеся в фазе эмбрионального роста. При подсыхании из листьев исчезает крахмал. Усиленный распад этого полисахарида не сопровождается накоплением гексоз, поскольку они используются в процессе дыхания. При потере воды усиливается распад белковых веществ.
Опыты с растениями тыквы и огурца (В.Н. Жолкевич, Т.Ф. Корецкая) показали, что почвенная засуха вызывает глубокое нарушение фосфорного обмена в корневой системе, что приводит к уменьшению в корнях количества АТФ, нуклеиновых кислот – РНК и ДНК, уменьшению содержания белков и увеличению в 7–10 раз сахаров, особенно фруктозы и глюкозы. Задержка фосфорилирования Сахаров вызывает уменьшение содержания органических кислот – пировиноградной кислоты, а также кислот цикла Кребса – α-кетоглютаровой, янтарной, фумаровой, щавелевоуксусной, яблочной и лимонной, являющихся акцепторами аммиака при синтезе аминокислот.
В корнях и в пасоке повышается содержание амидов, преимущественно глютамина. Следовательно, симптомами угнетения растений почвенной засухой прежде всего является нарушение энергетического обмена через уменьшение фосфорилирования, а также задержку синтеза белков. Длительные засухи приводят к резкому снижению урожая.
Как показали исследования Н.М. Сисакяна, при обезвоживании растительных тканей изменяется направленность действия ферментов в сторону гидролиза и задерживаются синтетические процессы и рост. Ростовые процессы задерживаются некоторое время даже после возобновления водоснабжения, поскольку восстановление синтетической направленности ферментов происходит не сразу. Различные части растения неодинаково устойчивы к засухе и неодинаково на нее реагируют.
Установлено, что растения, перенесшие небольшую засуху, повторную переносят с меньшими потерями. Однако устраивать искусственную закалку взрослых растений в производственных условиях трудно. П.А. Генкель предложил подвергать закаливанию наклюнувшиеся семена. Опыт показал, что растения, выросшие из семян, подвергнутых предпосевной закалке, приобретают повышенную устойчивость к засухе.
Растворы сахаров (глюкоза, галактоза, сахароза, маннит, лактоза, мальтоза, раффиноза) при инфильтрации в ткани листьев значительно повышают их устойчивость к высоким температурам. Считают, что сахара «консервируют» структуру митохондрии, которая становится нечувствительной к тепловому влиянию, и этим сохраняют функцию митохондрий – процесс дыхания и окислительное фосфорилирование (Ю.Г. Молотковский, И.М. Жесткова).
Засухоустойчивые сорта при значительном водном дефиците характеризуются синтетической направленностью в работе своих ферментов, тогда как у менее засухоустойчивых преобладает гидролитическая направленность. Засухоустойчивые сорта озимой пшеницы характеризуются большим содержанием связанной воды, которая трудно обменивается во время засухи, повышенной концентрацией клеточного сока в период цветения и налива зерна, высшим порогом коагуляции белков, более интенсивным накоплением сухого вещества зерна, более стойкой к неблагоприятным условиям пигментной системой. Этим и характеризуется физиологическая природа засухоустойчивости растений.
5. Источники углерода для растений. Усвоение углекислоты и лучистой энергии солнца при фотосинтезе. Лист как орган фотосинтезаУглекислый газ поступает в растения из воздуха, превращаясь с помощью лучистой энергии солнца в сложные, высокоэнергетические органические соединения, которыми питается животный мир. Животные, используя потенциальную энергию органических веществ, снова освобождают углекислый газ. Согласно современным представлениям, приведенное выше уравнение фотосинтеза можно изобразить в виде схемы:
Следовательно, фотосинтез состоит из двух сопряженных систем реакций: окисления воды до кислорода и восстановления углекислого газа водородом воды до полисахаридов.
Лист сверху и снизу покрыт бесцветной кожицей, малопроницаемой для газов кутикулой. Углекислый газ, который усваивается в процессе фотосинтеза, поступает в лист через устьица. На 1 см2 поверхности листа на долю устьиц приходится лишь 1 мм2, остальная площадь – на непроницаемую кутикулу. Диффузия СО2 в лист происходит очень интенсивно. Например, 1. см2 листовой поверхности катальпы поглощает 0,07 см3 СО2 за 1 ч, а такая же поверхность раствора щелочи – 0,12–0,15 см3, или в 2 раза больше.
В процентах указаны траты поглощенной листом световой энергии на различные виды работ
Для процесса фотосинтеза имеют значение особенности строения листа. К верхней стороне листа прилегает палисадная ткань, клетки которой расположены перпендикулярно, плотно соприкасаются друг с другом и богаты хлоропластами. Палисадная паренхима является преимущественно ассимиляционной тканью. К нижнему эпидермису прилегает губчатая паренхима с рыхлорасположенными клетками и межклетниками. Это приспособление у растений имеет значение для лучшего проникновения газов в клетки (рис. 1).
Чтобы процесс фотосинтеза проходил непрерывно, клетки должны быть достаточно насыщены водой. В этих условиях устьица до определенной степени бывают открыты. При этом будут осуществляться транспирация, газообмен, листья будут снабжаться в достаточной мере углекислым газом, т.е. процесс фотосинтеза будет проходить нормально.
Лист пронизан проводящими пучками, которые обеспечивают отток из него продуктов ассимиляции, что очень важно для нормального течения процесса фотосинтеза, поскольку в клетках, переполненных продуктами ассимиляции, в частности крахмалом, фотосинтез угнетается и может совсем прекратиться.
6. Выращивание растений при искусственном освещении. Условия наилучшего использования электрического света.Исследования показали, что на развитие растений в значительной мере влияет интенсивность и спектральный состав света. В связи с этим большой интерес представляют опыты В.И. Разумова, который доказал, что красный свет действует как естественное дневное освещение, а синий воспринимается растением как темнота. Если освещать растения короткого дня ночью красным светом, то они не зацветают; растения длинного дня в этих условиях зацветают раньше, чем в обычных. Освещение растений в ночное время синим светом не нарушает влияния темноты. Следовательно, длинноволновый свет воспринимается как дневной свет, а коротковолновый – как темнота. Таким образом, качественный состав света оказывает влияние на развитие растения.
Однако существует иной взгляд, а именно, что все световые лучи, если они достаточно интенсивны, воспринимаются растением как дневное освещение. Считают, что спектральный состав света в течение дня почти одинаковый. В значительной мере изменяется лишь его интенсивность – наименьшая утром и к вечеру и наибольшая в полдень.
Установлено, что свет люминесцентных ламп по спектральному составу сходен с солнечным светом, поэтому для выращивания растений при искусственном освещении используют именно эти лампы.
Светильники с люминесцентными лампами, преимущественно размещаются рядами, желательно параллельными стене с окнами или длинной стороне узкого помещения. Но в помещениях, предназначенных для растений, оптимальным является такое расположение светильников, при котором направление света приближается к направлению естественного света.
Необходимо помнить, что излишек света пагубно сказывается на растениях, процесс фотосинтеза приостанавливается, растения ослабевают и хуже переносят неблагоприятные условия. Наибольшую продолжительность светового дня переносит фасоль – до 12 часов.
7. Оксидазы, их участие в аэробном дыхании
Оксидазы – аэробные дегидрогеназы. для которых акцептором водорода может быть только кислород воздуха:
АН2+1/2О2 ↔ А+Н2О.
Ферменты оксидазы, которые активируют молекулярный кислород и делают его способным восстанавливаться до перекиси водорода, действуют на конечном этапе дыхания, когда водород окисляемого вещества надо выделять из системы:
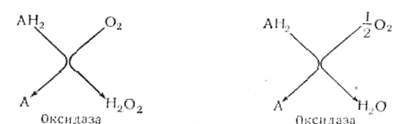
Эта группа ферментов многочисленна, но основная роль в ней принадлежит оксидазам, содержащим медь (полифенолоксидаза) и железо (цитохромоксидаза с цитохромами, цитохромная система).
Полифенолоксидазы или фенолоксидазы в присутствии молекулярного кислорода окисляют полифенолы в соответствующие хиноны. Фермент полифенолоксидаза встречается в тканях различных растений. Высокая активность полифенолоксидазы характерна для тканей листьев чая, картофеля, корней сахарной свеклы, клубней картофеля, семян люпина, гороха, тыквы и многих других растений.
Согласованность процесса окисления полифенолов с помощью полифенолоксидазы и восстановления хинонов можно наблюдать только в живой неповрежденной ткани растений. При повреждении ткани координация между окислительной и восстановительной фазами дыхания обычно нарушается, вследствие чего имеет место накопление различных темноокрашенных пигментов; например, при разрезании яблок, клубней картофеля.
Похожие работы
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... цитохромоксидаза, подтверждением чего служит также максимальная величина подавления дыхания азидом . Соотношения указанного типа не являются, однако, общей закономерностью, поскольку в ряде случаев максимальная интенсивность дыхания обнаруживается в клетках зоны растяжения. Такая закономерность выявляется при пересчете на белок, а также на единицу кислотонерастворимого фосфора. Вполне вероятно, ...
... и утрачивается к началу полового созревания. 36. Учение Павлова о типах ВНД. Общин типы ВНД. Пластичность ВНД. Учет типов ВНД в индивидуальном обучении и воспитании Среди вопросов физиологии ВНД человека особое значение для совершенствования учебно-воспитательного процесса и разработки естественнонаучных теорий воспитания и обучения имеет учение о типах ВНД. Это связано с тем, что именно ...
... относят противоязвенный фактор(витамин U), пангамовую кислоту (витамин В15), а также липоевую, оротовую, парааминобензойную кислоты и карнитин. Обмен веществ (метаболизм) совокупность всех химических изменений и всех видов превращений веществ и энергии в организмах, обеспечивающих развитие, жизнедеятельность и самовоспроизведение организмов, их связь с окружающей средой и адаптацию к изменениям ...
0 комментариев