Федеральное Агентство Науки и Образования
Сибирский Федеральный Университет
Институт Фундаментальной Биологии и Биотехнологии
Кафедра биотехнологии
РЕФЕРАТ
На тему: Основные субклеточные структуры растительной клетки
Выполнил: студент группы Б-31
Холодилова Валентина
Проверил: доцент кафедры
Биотехнологии, д. б. н
Голованова Т.И.
Красноярск, 2008 г.
Оглавление
Введение
Ядро
Аппарат Гольджи
Эндоплазматическая сеть (ЭПС)
Лизосомы
Сферосомы
Пероксисомы (микротельца)
Литература
Введение
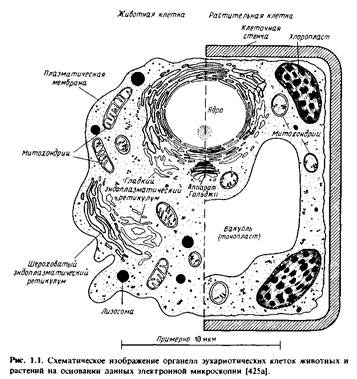
Клетки всех организмов имеют единый план строения, в котором четко проявляется общность всех процессов жизнедеятельности. Каждая клетка включает в свой состав две неразрывно связанные части: цитоплазму и ядро. Как цитоплазма, так и ядро характеризуются сложностью и строгой упорядоченностью строения и, в свою очередь, в состав их входит множество разнообразных структурных единиц, выполняющих совершенно определенные функции.
Ядро
Термин "ядро" (лат. nucleus) впервые применил Р. Броун в 1833 году, когда описывал шарообразные структуры, наблюдаемые им в клетках растений.
Ядерная оболочка. Внутреннее пространство клеточного ядра отделено от цитоплазмы ядерной оболочкой, состоящей из двух мембран. Мембраны оболочки ядра сходны по строению с другими мембранными компонентами клетки и построены по тому же принципу: это тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который встроены молекулы белков. Пространство между внутренней и внешней ядерными мембранами называется перинуклеарным. На поверхности внешней ядерной мембраны обычно располагается большое количество рибосом, и иногда удается наблюдать непосредственный переход этой мембраны в систему каналов гранулярной эндоплазматической сети клетки. Внутренняя ядерная мембрана связана с тонким волокнистым белковым слоем - ядерной ламиной, состоящей из белков ламинов. Густая сеть фибрилл ядерной ламины способна обеспечить целостность ядра, даже после растворения липидных мембран оболочки ядра в эксперименте. С внутренней стороны к ламине крепятся петли хроматина, заполняющего ядро.
Ядерная оболочка имеет отверстия диаметром около 90 нм, образующиеся за счет слияния внешней и внутренней ядерных мембран. Такие отверстия в оболочке ядра окружены сложными белковыми структурами, получившими название комплекса ядерной поры. Восемь белковых субъединиц, входящих в состав ядерной поры, располагаются вокруг перфорации ядерной оболочки в виде колец, диаметром около120 нм, наблюдаемых в электронный микроскоп с обеих сторон ядерной оболочки. Белковые субъединицы комплекса поры имеют выросты, направленные к центру поры, где иногда видна "центральная гранула" диаметром 10-40 нм. Размер ядерных пор и их структура стандартны для всех клеток эукариот. Число ядерных пор зависит от метаболической активности клеток: чем выше уровень синтетических процессов в клетке, тем больше пор на единицу площади поверхности клеточного ядра. В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некое молекулярное сито, пропуская ионы и мелкие молекулы (сахара, нуклеотиды, АТФ и др.) пассивно, по градиенту концентрации, и осуществляя активный избирательный транспорт крупных молекул белков и рибонуклеопротеидов, то есть комплексов рибонуклеиновых кислот (РНК) с белками. Так, например, белки, транспортируемые в ядро из цитоплазмы, где они синтезируются, должны иметь определенные последовательности примерно из 50 аминокислот, (т. наз. NLS последовательности), "узнаваемые" комплексом ядерной поры. В этом случае комплекс ядерной поры, затрачивая энергию в виде АТФ, активно транслоцирует белок из цитоплазмы в ядро.
Хроматин. Клеточное ядро является вместилищем практически всей генетической информации клетки, поэтому основное содержимое клеточного ядра - это хроматин: комплекс дезоксирибонуклеиновой кислоты (ДНК) и различных белков. В ядре и, особенно, в митотических хромосомах, ДНК хроматина многократно свернута, упакована особым образом для достижения высокой степени компактизации. Ведь все длинные нити ДНК, общая длина которых составляет, например, у человека около 164 см, необходимо уложить в клеточное ядро, диаметр которого всего несколько микрометров. Эта задача решается последовательной упаковкой ДНК в хроматине с помощью специальных белков. Основная масса белков хроматина - это белки гистоны, входящие в состав глобулярных субъединиц хроматина, называемых нуклеосомами. Всего существует 5 видов белков гистонов. Нуклеосома представляет собой цилиндрическую частицу, состоящую из 8 молекул гистонов, диаметром около 10 нм, на которую "намотано" чуть менее двух витков нити молекулы ДНК. В электронном микроскопе такой искусственно деконденсированный хроматин выглядит как "бусины на нитке". В живом ядре клетки нуклеосомы плотно объединены между собой с помощью еще одного линкерного гистонового белка, образуя так называемую элементарную хроматиновую фибриллу, диаметром 30 нм. Другие белки, негистоновой природы, входящие в состав хроматина обеспечивают дальнейшую компактизацию, т.е. укладку, фибрилл хроматина, которая достигает своих максимальных значений при делении клетки в митотических или мейотических хромосомах. В ядре клетки хроматин присутствует как в виде плотного конденсированного хроматина, в котором 30 нм элементарные фибриллы упакованы плотно, так и в виде гомогенного диффузного хроматина. Количественное соотношение этих двух видов хроматина зависит от характера метаболической активности клетки, степени ее дифференцированности. Так, например, ядра эритроцитов птиц, в которых не происходит активных процессов репликации и транскрипции, содержат практически только плотный конденсированный хроматин. Некоторая часть хроматина сохраняет свое компактное, конденсированное состояние в течение всего клеточного цикла - такой хроматин называется гетерохроматином и отличается от эухроматина рядом свойств.
Репликация и транскрипция. Клетки эукариот содержат обычно несколько хромосом (от двух до нескольких сотен), которые теряют в ядре (в интерфазе, т.е. между митотическими делениями) клетки свою компактную форму, разрыхляются и заполняют объем ядра в виде хроматина. Несмотря на деконденсированное состояние, каждая хромосома занимает в ядре строго определенное положение и связана с ядерной оболочкой посредством ламины. Строго закреплены на внутренней поверхности оболочки ядра такие структуры хромосом, как центромеры и теломеры. На определенной стадии жизненного цикла клетки, в синтетическом периоде, происходит репликация, т.е. удвоение всей ДНК ядра, и хроматина становится в два раза больше. Белки, необходимые для этого процесса, поступают, конечно, из цитоплазмы через ядерные поры. Таким образом, клетка готовится к предстоящему клеточному делению - митозу, когда общее количество ДНК в ядре вернется к первоначальному уровню.
Реализация генетической информации, заключенной в ДНК в виде генов, начинается с транскрипции, т.е. с синтеза информационных РНК (и-РНК) - точных копий генов, по которым затем будут строиться в цитоплазме на рибосомах белки. Этот процесс проходит в различных точках в объеме ядра, морфологически ничем не отличающихся от окружающего хроматина. Чаще всего удается наблюдать транскрипцию диффузного, т.е. деконденсированного хроматина.
Кроме хроматина, составляющего хромосомы, в ядрах эукариот обычно содержится одно или несколько ядрышек. Это плотные структуры, не имеющие собственной оболочки и представляющие собой скопления молекул другого типа РНК - рибосомной РНК (р-РНК) в комплексе с белками. Такие комплексы называют рибонуклеопротеидами (РНП). Ядрышки имеют стандартную морфологию и образуются в ядре после деления клетки вокруг постоянно действующих точек активного синтеза рибосомной РНК. Гены рибосомной РНК, в отличие от большинства других генов, кодирующих белки, содержатся в геноме в виде многочисленных копий. Эти копии, расположенные в молекуле ДНК тандемно, т.е. друг за другом, располагаются в определенных районах нескольких хромосом генома. Такие районы хромосом называют ядрышковыми организаторами. Морфологически в ядрышке с помощью электронного микроскопа можно выделить следующие 3 зоны: гомогенные компактные фибриллярные центры, содержащие ДНК ядрышковых организаторов; плотный фибриллярный компонент вокруг них, где идет транскрипция генов рибосомной РНК и массивный гранулярный компонент ядрышка, состоящий из частиц РНП - будущих рибосом. Эти гранулы РНП, образующиеся в ядрышке, транспортируются в цитоплазму и образуют рибосомы, осуществляющие синтез всех белков клетки. Третий основной тип клеточных РНК - мелкие транспортные РНК - транскрибируются в различных участках ядра и выходят в цитоплазму через ядерные поры. Там они, как известно, обеспечивают транспортировку аминокислот к рибосомам в процессе синтеза белков.
Ядерный белковый матрикс. Для осуществления процессов репликации, транскрипции, а также поддержания определенного положения хромосом в объеме ядра существуют каркасные белковые структуры, называемые ядерным белковым матриксом. Такой матрикс состоит, по крайней мере из трех морфологических компонентов: периферического фиброзного слоя - ламины; внутреннего, или интерхроматинового матрикса ядра и матрикса ядрышка. Наблюдения показывают, что компоненты ядерного матрикса - это не жесткие застывшие структуры, они динамичны и могут сильно видоизменяться в зависимости от функциональных особенностей ядер. Показано, что белковый матрикс имеет множество точек прочного связывания с ДНК ядра, которая, в свою очередь, имеет специальные последовательности нуклеотидов, необходимые для этого.
Аппарат ГольджиАппарат Гольджи - это органоид клетки, получивший свое название по имени ученого К. Гольджи, который впервые увидел его в цитоплазме нейронов и назвал сетчатым аппаратом (1898). Во многих клетках этот органоид действительно имеет форму сложной сети, расположенной вокруг ядра. Иногда же его сетевидная структура приобретает вид шапочки, расположенной над ядром, или тяжа, опоясывающего ядро. В клетках многих беспозвоночных животных и растений комплекс Гольджи представлен в виде отдельных элементов, обладающих формой округлых, серповидных или палочковидных телец, носящих название диктиосом. Такая рассеянная форма аппарата Гольджи свойственна и некоторым клеткам позвоночных животных. Исследование многочисленных клеток животных и растений с помощью электронного микроскопа показало, что, несмотря на многообразие формы и строения комплекса Гольджи, структура его элементов однотипна в разных клетках. По данным электронно-микроскопического исследования, ультраструктура комплекса Гольджи включает три основных компонента.1. Система плоских цистерн, ограниченных гладкими мембранами. Цистерны расположены пачками, по 5 - 8; причем они плотно прилегают друг к другу. Количество цистерн, их величина и расстояние между ними варьируют в разных клетках.2. Система трубочек, которые отходят от цистерн. Трубочки анастомозируют друг с другом и образуют довольно сложную сеть, окружающую цистерны.3. Крупные и мелкие пузырьки, замыкающие концевые отделы трубочек. Все три компонента аппарата Гольджи взаимосвязаны друг с другом и могут возникать друг из друга. Согласно электронно-микроскопическим данным, мембранам всех трех компонентов свойственно такое же трехслойное строение, как и наружной цитоплазматической мембране и мембранам эндоплазматической сети. В состав мембран аппарата Гольджи входят липиды, или, точнее, фосфолипиды и белки. Следовательно, в мембранах его содержится тот же белково-липидный комплекс, что и в мембранах других клеточных органоидов. В элементах комплекса Гольджи обнаружены ферменты и среди них ферменты, связанные с синтезом полисахаридов и липидов. Структуры аппарата Гольджи накапливают либо уже готовые, либо почти готовые продукты деятельности клеток. Формирование и накапливание секреторных гранул - это основная, очень важная, но не единственная функция аппарата Гольджи. При делении клеток часть аппарата Гольджи из материнской клетки передается в дочернюю. Этот клеточный органоид представляет, поэтому, преемственную структуру, и при делении обычно материал его распределяется поровну между материнской и дочерней клетками. Возможность образования аппарата Гольджи заново не доказана.
Эндоплазматическая сеть (ЭПС)
Эндоплазматическая сеть принадлежит к числу органоидов клетки, открытых совсем недавно (1945 - 1946). Расположение сетчатых структур во внутренней части цитоплазмы - эндоплазме (греч. "эндон" - внутри) - и послужило основанием для того, чтобы вновь открытому органоиду дать название эндоплазматической сети или эндоплазматического ретикулума. Дальнейшее электронно-микроскопическое изучение ультратонких срезов разнообразных клеток показало, что сетчатые структуры состоят из сложной системы канальцев, вакуолей и цистерн, ограниченных мембранами. Мембраны ЭПС имеют типичную трехслойную структуру, такую же, как и та, что свойственна и наружной мембране клетки. Каналы, вакуоли и цистерны образуют ветвящуюся сеть, которая пронизывает всю цитоплазму клетки. Форма каналов, вакуолей и цистерн эндоплазматической сети непостоянна и широко варьирует как в одной и той же клетке в разные периоды ее функциональной деятельности, так и в клетках различных органов и тканей. Для каждого типа клеток характерна определенная структура ЭПС.
Типы эндоплазматической сети Детальное изучение мембран, ограничивающих каналы, вакуоли и цистерны ЭПС, позволило установить, что во многих клетках на наружной поверхности этих мембран располагаются многочисленные округлые плотные гранулы. Эти гранулы носят название рибосом. Рибосомы часто образуют скопления на поверхности мембран, ограничивающих цистерны и каналы. Однако есть участки ЭПС, где рибосом нет. Поэтому в клетках различаются два типа эндоплазматической сети: гранулярная, или шероховатая, т.е. несущая рибосомы, и гладкая.
Зная, что рибосомы в цитоплазме служат местом синтеза белка, можно предположить, что гранулярная сеть в большей степени представлена в тех клетках, где идет активный синтез липидов. Оба вида ЭПС не только участвуют в синтезе органических веществ, но и накапливают и транспортируют их к местам назначения, регулируют обмен веществ между клеткой и окружающей ее средой. ЭПС была обнаружена во всех клетках многоклеточных животных и растений, подвергавшихся электронно-микроскопическому исследованию.
Функции ЭПС. Столь широкое распространение в клетках всех типов у огромного большинства организмов позволяет рассматривать ЭПС как один из универсальных клеточных органоидов, выполняющих важные и разносторонние функции Совершенно определенно установлено, что гранулярная ЭПС принимает активное участие в синтезе белка.
Обе формы ЭПС способны накапливать в каналах, вакуолях и цистернах продукты синтеза: белковые вещества (гранулярная ЭПС) и жиры или гликоген (гладкая ЭПС). Все эти вещества, накапливающиеся в просвете каналов и цистерн, в дальнейшем транспортируются к различным органоидам клетки. Транспортная роль ЭПС не ограничивается только передвижением и распределением синтезированных на ее мембранах веществ к разным органоидам клетки, т.е. к местам, где они либо потребляются, либо накапливаются. ЭПС представляет циркуляторную систему клетки, через которую происходит транспорт разнообразных веществ из окружающей среды в цитоплазму. Кроме того, ЭПС, соединяясь со многими органоидами клетки, обеспечивает связь между отдельными внутриклеточными структурами.
ЛизосомыЛизосомы (от лиз- и греч. soma - тело), структуры в клетках животных и растительных организмов, содержащие ферменты, способные расщеплять (т.е. лизировать - отсюда и название) белки, полисахариды, пептиды, нуклеиновые кислоты.
Это очень пестрый класс пузырьков размером 0,1-0,4 мкм, ограниченных одиночной мембраной (толщиной около 7 нм), с разнородным содержимым внутри. Они образуются за счет активности эндоплазматического ретикулума и аппарата Гольджи и в этом отношении напоминают секреторные вакуоли. Основная их роль - участие в процессах внутриклеточного расщепления как экзогенных, так и эндогенных биологических макромолекул. Характерной чертой лизосом является то, что они содержат около 40 гидролитических ферментов: протеиназы, нуклеазы, фосфатазы, гликозидазы и др., оптимум действия которых осуществляется при рН5. В лизосомах кислое значение среды создается из-за наличия в их мембранах протоновой "помпы", потребляющей энергию АТФ. Кроме того, в мембраны лизосом встроены белки-переносчики для транспорта из лизосомы в цитоплазму продуктов гидролиза: мономеров расщепленных молекул - аминокислот, сахаров, нуклеотидов, липидов. Чтобы не переварить самих себя, мембранные элементы лизосом защищены олигосахаридами, мешающими гидролазам взаимодействовать с ними. Среди различных по морфологии лизосомных частиц выделяют четыре типа: первичные и вторичные лизосомы, аутофагосомы и остаточные тельца.
Первичные лизосомы. Обычно это мелкие мембранные пузырьки диаметром около 100 нм с бесструктурным содержимым, содержащим активную кислую фосфатазу - маркерный фермент для лизосом. Показано, что ферменты лизосом синтезируются, как обычно, в гранулярном ретикулуме, а упаковываются в мембранные пузырьки в аппарате Гольджи. Весь путь образования первичных лизосом очень сходен с образованием секреторных гранул, например, в клетках поджелудочной железы.
Вторичные лизосомы. Фагоцитарные или пиноцитозные вакуоли, сливаясь с первичными лизосомами, образуют вторичные лизосомы. Начинается процесс расщепления поглощенных клеткой субстратов под действием гидролаз, содержавшихся в первичной лизосоме. Биогенные вещества расщепляются до мономеров, которые транспортируются через мембрану лизосомы в цитоплазму, где они реутилизируются, включаются в различные синтетические и обменные процессы. Разнообразие величины и структуры клеточных лизосом связано в первую очередь с разнообразием вторичных лизосом - продуктов слияния эндоцитозных вакуолей с первичными лизосомами.
Остаточные тельца. Расщепление биогенных макромолекул внутри лизосом может идти не до конца. В этом случае в полости лизосомы накапливаются непереваренные продукты, и вторичная лизосома становится остаточным тельцем (телолизосомой). Их содержимое уплотняется и перестраивается. Часто в остаточных тельцах наблюдается вторичная структуризация непереваренных липидов, которые образуют сложные слоистые структуры. Там же откладываются пигментные вещества.
Аутолизосомы. По морфологии аутолизосомы (аутофагосомы) относят ко вторичным лизосомам, но внутри этих частиц встречаются фрагменты или даже целые цитоплазматические структуры, такие как митохондрии, пластиды, рибосомы, элементы ретикулюма и т.д. Предполагается, что процесс аутофагоцитоза связан с отбором и уничтожением измененных, "сломанных" клеточных компонентов.
СферосомыЭто мембранные пузырьки, встречающиеся в клетках растений, они окрашиваются липофильными красителями, имеют высокий коэффициент преломления и поэтому хорошо видны в световой микроскоп. Сферосомы образуются из элементов эндоплазматического ретикулума. На конце цистерны ЭР начинает накапливаться осмиофильный материал, затем от этого участка отшнуровывается и начинает расти мелкий пузырёк, достигающий диаметра 100-150 нм. Это "просферосома", окруженная одинарной двуконтурной мембраной. Рост сферосом и перестройка их содержимого связаны с накоплением в них масла, так что сферосома постепенно превращается в масляную каплю. Кроме жиров в составе сферосом обнаруживают белки и среди них фермент липазу, встречающуюся во всех изученных сферосомах.
Пероксисомы (микротельца)Это небольшие вакуоли (0,3-1,5 мкм), одетые одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина или нуклеотид (ничего не имеющий общего с нуклеотидом бактерий и вообще к ядерным структурам не относящийся).
В зоне сердцевины часто видны кристаллоподобные структуры, состоящие из регулярно упакованных фибрилл или трубочек. Изолированные сердцевины пероксисом содержат фермент урат-оксидазу.
Пероксисомы обнаружены у простейших (амёбы, тетрахимена), у низших грибов (дрожжи), у высших растений в некоторых эмбриональных тканях (эндосперм) и в зеленых частях, способных к фотореспирации, у высших позвоночных животных они обнаруживаются главным образом в печени и почках.
Пероксисомы бывают тесно связаны с мембранами ЭР и, вероятно, образуются из расширенных концов цистерн ЭР, которые заполняются плотным материалом. У зелёных растений пероксисомы часто находятся в тесном контакте с митохондриями и пластидами.
Впервые пероксисомы были выделены из печени и почек. Во фракциях пероксисом обнаруживаются ферменты, связанные с метаболизмом перекиси водорода. Это ферменты (оксидазы, уратоксидаза, оксидаза d-аминокислот) окислительного дезаминирования аминокислот, при работе которых образуется перекись водорода и каталаза, разрушающая её. Так как перекись водорода является токсическим веществом для клеток, то каталаза пероксисом может играть важную защитную роль.
У животных и некоторых растений (проростки клещевины) пероксисомы играют важную роль при превращении жиров в углеводы. Так, в клетках эндосперма клещевины в пероксисомах (глиоксисомах) содержатся ферменты глиоксалатного цикла.
Литература
1. Ченцов Ю.С. Общая цитология. М., Изд-во Моск. ун-та, 1978. - 344 с., с ил.
2. http://www.krugosvet.ru
Похожие работы
... ассоциированные с ними дополнительные белки, так называемые МАР-белки. Эти белки, стабилизируя микротрубочки, ускоряют процесс полимеризации тубулина. Строение и функции микрофиламентов Микрофиламенты представляют собой очень тонкие и длинные нитевидные белковые структуры, встречающиеся во всей цитоплазме. Под плазматической мембраной микрофиламенты образуют сплошное сплетение, формируя ...
... хотя бы некоторое представление о соответствующих методах исследования клетки. Световая микроскопия Самым древним и, вместе с тем, наиболее распространенным методом изучения клетки является микроскопия. Можно сказать, что и начало изучения клетки было положено изобретением светового оптического микроскопа. Невооруженный человеческий глаз имеет разрешающую способность около 1/10 мм. Это ...
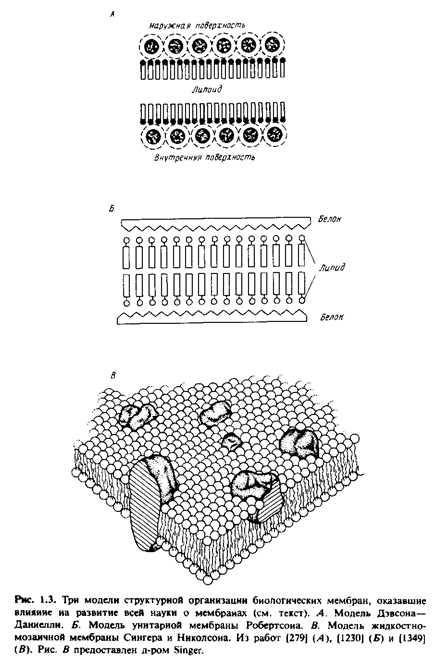
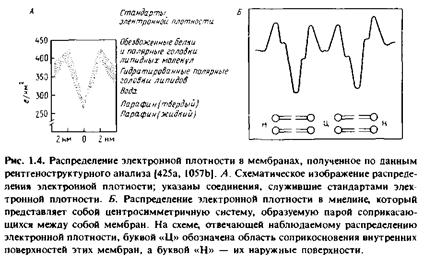
... правильным только благодаря взаимной компенсации ошибок, однако в историческом плане эта работа имела большое значение, поскольку с тех пор концепция липидного бислоя как структурной основы биологических мембран стала доминирующей и на самом деле оказалась верной. Концепция бимолекулярной липидной мембраны получила дальнейшее развитие в предложенной в 1935 г. модели Дэвсона-Даниелли, или ...
... , входящие в состав каждого из этих препаратов, можно разделить на две основные популяции. Однако элетронно-микроскопическое исследование не позволило обнаружить никаких различий в структуре этих частиц. Рабочая методика выделения митохондрий из печени крыс. • Среда выделения: 0,25 М раствор сахарозы, содержащий 0,001 М ЭДТА, рН 7,4 • Сахароза - 0,25 М раствор • НС1 - 1 н и 0,1 н растворы ...
0 комментариев