Процесс опыления у растений
Опыление - жизненно важный процесс для всех цветковых растений, и природа немало позаботилась о том, чтобы он проходил успешно.
В отличие от животных, растения не могут передвигаться в поисках партнера, и им приходится полагаться на помощь внешних сил (ветра, воды, насекомых), чтобы передать пыльцу другому растению (или другой своей части) для создания новых семян.
Опыление так важно для растения потому, что от него зависит сохранение вида. А перекрестное опыление - когда пыльца переносится на рыльце другого цветка, на том же растении или другом, но того же вида - может привести к мутациям, которые помогут данному виду лучше приспособиться к окружающей среде.
Опыление ветром
Опыление ветром, когда невесомая пыльца переносится с потоками воздуха, очень распространено в природе. Так опыляются многие деревья, как, например, дуб, ясень и сосна, а также кукуруза и злаковые. Опыляемые ветром растения вынуждены производить огромное количество пыльцы, чтобы увеличить шансы ее попадания на рыльце соответствующего растения. Пыльца должна быть очень легкой, чтобы "плыть" по воздуху; она почти невесома. Если встряхнуть созревшую сережку орешника, цветущую ветку хвойного дерева или тимофеевку луговую, вы увидите в воздухе целое облако пыльцы. У некоторых растений есть крохотные воздушные пузырьки, которые помогают пыльце подольше продержаться на ветру.
Опыление насекомымиКак правило, цветы растений, опыляемых насекомыми, очень яркие и обладают сильным ароматом. Если отдельные цветочки слишком малы, они группируются в соцветия или располагаются в окружении разноцветных листьев под названием прицветник, чтобы привлечь внимание насекомых.
"Цветы" мексиканского кустарника пуансетии в действительности являются прицветником, привлекающим внимание насекомых к достаточно неприметным цветам. Пыльцевые зерна опыляемых насекомыми цветов обычно крупнее и шероховатей, чем у цветов, опыляемых ветром. Они могут быть клейкими, чтобы прилипать к насекомым.
Адаптация к насекомым
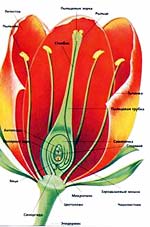
У растений, опыляемых насекомыми, можно наблюдать чрезвычайно остроумные и изобретательные приемы адаптации к ним. Растения с открытыми (например, маргаритки) или чашевидными (лютики) цветами устроены так, что могут воспользоваться услугами всех насекомых, будь то пчела, жук или муравей, а иногда они не брезгуют и помощью мелких животных, прикоснувшихся к цветку. Насекомое может стряхнуть пыльцу из пыльника на рыльце или перенести ее на другой цветок. Некоторые цветы более разборчивы и могут опыляться с помощью только одного вида насекомых. Цветы люпина, душистого горошка и их ближайших родственников устроены так, что раскрываются под весом насекомого, севшего к ним на "крылышки", и выпускают тычинки и рыльца, трущиеся о тельце гостя. У некоторых растений цветы могут опылить только насекомые с длинным хоботком, такие как мотыльки и пчелы. Примером такого растения может послужить яснотка белая. Чтобы добраться до нектара, спрятанного на дне цветка, насекомому приходится просовывать голову глубоко вовнутрь и тесно прижиматься к нижнему лепестку. В это время пыльник или рыльце, расположенные в верхних лепестках, прижимаются к спинке насекомого. У некоторых растений с перекрестным опылением сначала из цветка появляется пыльник, и только когда вся пыльца высыплется, выглядывает рыльце, чтобы именно оно, а не пыльники, пришло в непосредственный контакт с насекомым. Опыление насекомыми - это основной вид опыления цветов, однако в тропиках Австралии и Южной Америки довольно распространено опыление птицами. Это главным образом колибри, и зачастую они не больше шмеля. Они сосут нектар закрытым клювиком, используя язык как поршень.
Водное опылениеЭто самый редкий вид опыления, но он является основным для сугубо водных цветущих растений - например, зостеры (морской травы). Их нитеобразная пыльца с удельным весом, соответствующим плотности морской воды, может плавать на любой глубине, пока ее не захватит похожее на перышко рыльце.
Свободное опыление как метод селекции часто используется начинающими селекционерами, однако уже после первых лет работы с такими сеянцами становится ясной малая перспективность этого метода для получения форм с хорошими декоративными признаками. Как правило, в потомстве от свободного опыления появляются признаки, характерные для диких предков культурного гладиолуса, и почти отсутствуют те, которые появились позже в процессе селекции. Более перспективной для селекции является, несомненно, искусственная гибридизация с продуманным подбором родительских пар. Вместе с тем можно полагать, что потомство от свободного опыления будет биологически более устойчивым и жизнеспособным. Наблюдение за потомством от свободного опыления сортов гладиолуса показало, что лучше всего передается потомству окраска цветка материнского растения. Даже в потомстве белоцветковых сортов, являющихся рецессивными по окраске, 80-90 % сеянцев имели белые цветки. Так как в коллекции сорта территориально близко расположены друг к другу и растений каждого сорта относительно немного, то предположить во всех случаях опыление в пределах своего сорта трудно, хотя это и не исключено. Преобладала окраска цветка материнского сорта у сеянцев лососево-розовых и оранжевых сортов. Очень стойко передается штриховатость окраски; все сеянцы от свободного опыления розового с малиновыми штрихами сорта Биби имели примерно такую же окраску цветка; 54,5 % гибридов от свободного опыления сорта Северное сияние имели кремово-белые цветки с розово-красными штрихами, сходные с окраской материнского сорта. Форма цветка материнского сорта при свободном опылении тоже наследуется значительным числом сеянцев.
Так, от гофрированных сортов Алекс, Сноуспрайт все сеянцы имели такую же форму цветка, тогда как от сортов с простым цветком все сеянцы имели ровные края околоцветника. Однако были и исключения. Так, от сорта Джилси Карнавал, имеющего оранжевые с красными пятнами гофрированные цветки, лишь у 16,6 % гибридов были такие же цветки. По окраске 50 % сеянцев оказались желтыми, 33,4 % имели кремовые, розово-красные и розово-лиловые цветки с ровными долями. От сорта Пинк Сайз (беловато-сиреневый очень мелкоцветковый) были получены светло-фиолетовые сеянцы с крупным цветком, и только треть из них по форме и окраске походила на материнский сорт. Большинство сеянцев от свободного опыления имеют невыровненное соцветие, редкое расположение цветков, длинную трубку, что делает цветок сильно наклоненным. Эти признаки свойственны дикорастущим видам гладиолуса. Так, капюшоновидный сильно наклоненный верхний сегмент околоцветника, часто встречающийся у сеянцев от свободного опыления (и у некоторых сортов), характерен для гладиолуса попугайного (G. psittacinus) и гладиолуса первоцветного (G. primulinus), использовавшихся в селекции культурных форм. Воронковидная форма цветка, очевидно, исходит от гладиолуса кардинальского (G. cardinalis). Сильно изогнутая и длинная трубка - от гладиолуса пурпурно-золотистого (G. purpureo-auratus) или гладиолуса первоцветного. Редкое соцветие, небольшое число цветков, рыхлое их расположение - все эти признаки нередко доминируют у сеянцев от свободного опыления, так как являются признаками диких предков культурного гладиолуса. Вследствие этого свободное опыление как метод селекции новых сортов оказывается мало перспективным, особенно когда селекция ведется по декоративным признакам. Возможно, что при селекции на устойчивость сеянцы от свободного опыления окажутся перспективными. Близкородственное скрещивание является одним из методов селекции растений и животных. Он приводит к процессу разложения популяции, каковой является каждый сорт культурного растения, на линии с различными генотипами, при этом гены, находящиеся в гегерозиготном состоянии, переходят в гомозиготное. При последующих скрещиваниях этих линий могут возникнуть особи, сочетающие в себе ряд признаков, находившихся ранее в рецессивном состоянии.
Получающиеся в результате инбридинга растения оказываются более выровненными, однородными и надежнее передают свои свойства потомству. Известно между тем, что близкородственное скрещивание приводит к уменьшению потомства за счет гомозиготизации летальных генов. У гладиолуса вследствие его полиплоидности снижение жизненности при однократном инцухте сказывается слабо, однако при опылении цветков гладиолуса своей пыльцой (пыльцой с другого цветка этого же растения) уменьшается число коробочек и созревших семян по сравнению с этими показателями при искусственном межсортовом опылении. Гибель растений, полученных из семян от близкородственного опыления, отмечается на стадии проростков и дальнейшего развития молодых растений, так что цветения достигает едва половина всех всходов, развитие их происходит медленно, так что цветение наступает не ранее чем на 4-й год после посева, тогда как сеянцы от скрещивания или свободного опыления зацветают на 2-3-й год. Так как почти все сорта гладиолуса произошли от скрещивания сортов друг с другом, то потомство от самоопыления по существу является гибридом второго поколения (F2) и в нем проявляются признаки исходных сортов.
Так, в потомстве от самоопыления крупноцветкового сорта Раннего розового, происшедшего от скрещивания сортов Хеппи Энд и Спринг Мейд, 86,6 % гибридов имели мелкий цветок, характерный для отцовского сорта (Спринг Мейд), и 13,4 % - средний, как у материнского сорта- Окраска цветка варьировалась от белой до светло-красной (исходные сорта имели оранжево-красные и желтые цветки). От самоопыления мелкоцветкового, желтого сорта Спринг Мейд получились сеянцы с преобладанием желтых мелких цветков. В потомстве от самоопыления у гладиолуса проявляются рецессивные признаки (крупный цветок, белая окраска); оно характеризуется большей однородностью окраски цветка, но вместе с тем и большим количеством "малокультурных" признаков: короткое редкое соцветие, неправильное расположение цветков, слабые доли, длинная трубка. Сеянцы от самоопыления сами не дают материала для отбора перспективных форм, но могут быть использованы для повторных скрещиваний с целью создания гетерозисных мощных форм культурного гладиолуса. Пока что этот прием в селекции гладиолуса используется слабо.
Опыление плодовых культур заметно отличается от опыления полевых и огородных культур. Плодовые растения цветут в ранние весенние месяцы, когда дикие насекомые и медоносные пчелы еще малочисленны. Лёту насекомых-опылителей препятствует холодная погода. Большинство плодоводов хорошо знает, что при холодной погоде в саду особенно необходимы медоносные пчелы. При нормальном цветении взрослых деревьев на гектаре яблоневого сада бывает примерно 2,5 млн. цветков, из которых только 3—5% оплодотворяются и превращаются в пригодные для реализации плоды.
Среди плодовых культур сорта яблони и черешни являются лучшими примерами самостерильности. Чтобы происходило эффективное перекрестное опыление, в саду должны быть совместимые сорта. Если в саду растут яблони двух или большего числа сортов, но при перекрестном опылении плоды не завязываются, значит, сорта интерстерильны. Практически все сорта груш самостерильны, хотя для них характерны большие колебания степени стерильности, чем у яблонь и черешен. Сливы японского происхождения наиболее самостерильны, в то время как сливы европейского происхождения обладают склонностью к самофертильности. Сорта вишен, персиков, черной смородины, малины и ежевики практически самофертильны, но для гарантии самоопыления им нужны насекомые.
Самостерильным сортам необходимы насекомые, обеспечивающие эффективное перекрестное опыление. Часто придерживаются неправильной точки зрения, что у самофертильных растений опыление происходит без участия насекомых. Перекрестное опыление между двумя самофертильными сортами всегда обеспечивает лучший урожай. Для завязывания плодов у самофертильных типов растений требуется меньшая работа насекомых, чем для образования урожая у самостерильных растений. Однако крайне важно, чтобы насекомые равномерно распределялись по саду. У самостерильных сортов даже при удачном размещении сортов-опылителей перекрестное опыление зависит от многих взаимосвязанных сложных факторов. Не все посещения цветков насекомыми оказываются эффективными. Так, насекомое, хорошо покрытое пыльцой, обеспечивает перекрестное опыление лишь нескольких цветков, потому что запас полноценной пыльцы на теле насекомого быстро расходуется.
Для опыления в саду при неблагоприятной погоде нужны семьи, состоящие примерно из 2,3 кг взрослых пчел и не менее 6 рамок расплода. Если для опыления пчелы закупаются на юге, то вес пакета для заселения одного улья должен быть не менее 2,3 кг. Пакетные пчелы должны прибыть на место за 3—5 дней до начала цветения, чтобы они успели образовать семью и освоились с новой местностью до начала опылительной работы. Пчелам следует предоставить достаточно места для складывания запасов корма, а матке — для откладки яиц. Для перезимовавших семей вполне достаточен двухкорпусный улей. Пакетным пчелам второй ульевой корпус с сотами дают примерно через 10 дней после пересадки их в улей, что - бы они имели достаточно места для расплода и складывания запасов корма. Следует шире использовать безматочные пакеты пчел для пополнения слабых семей.

Рис. 4. Использование двух пакетов пчёл (по 1,3 кг) часто даёт лучшие результаты чем использование одного пакета пчёл весом 2,3 кг.
Пчеловоды юга высылают также пакеты пчел (2,3 кг) специально для садов. Эти пакеты используются в тех случаях, когда невозможно достать ульи с пчелами. Пчелы специальных пакетов для сада посещают цветки хуже, чем пчелы перезимовавших семей или пчелы, пересаженные из пакетов в ульи (рис. 4). Для мелких плодоводов, владеющих плодовыми садами площадью от 1,2 до 4 га, лучше иметь собственные пчелиные семьи. Крупным плодоводам в большинстве случаев выгоднее арендовать семьи у заслуживающих доверие пчеловодов.
Размещение пчелиных семей зависит от размера сада. Если сад занимает 1—2,5 га, ульи можно расположить в защищенном месте внутри сада или по его краю, и пчелы будут равномерно посещать все цветущие деревья. В больших садах ульи распределяют из расчета 2,5 семьи на гектар. При такой расстановке ульев возможно хорошее опыление даже при низких температурах, слабом солнечном сиянии и преобладании сильного ветра.
Наличие пчел в садах с перекрестно-опыляемыми деревьями не обеспечивает получение урожая, если отсутствуют сорта-опылители (совместимые сорта). Деревья основных сортов следует высаживать блоками, состоящими из 3—4 рядов. Вокруг блоков должны находиться сорта-опылители, причем сроки цветения у всех сортов должны совпадать. При неправильной посадке в ведра с водой ставят цветущие ветки сортов-опылителей. Ведра подвешивают на треногах из кольев возле ветвей дерева, нуждающегося в перекрестном опылении. Поскольку подвешивание ведер с ветками является временной мерой, в крону основного сорта нужно привить глазок или черенок сорта-опылителя. Через 3—5 лет привой начинает цвести. Для перекрестного опыления самофертильных сортов крайне важно иметь правильно размещенные промежуточные посадки самофертильных сортов - опылителей.
Насекомые не одинаково активно посещают цветки разных растений, что объясняется в основном неодинаковым содержанием сахара в нектаре, а также разным ароматом цветков. Медоносные пчелы собирают самый сладкий нектар, а так как концентрация сахара в нектаре меняется в течение дня, то и активность лёта пчел бывает разной. Указанная особенность растений приводит к тому, что на деревьях одних сортов в саду завязывается слишком много плодов, а с деревьев других сортов не снимают даже среднего урожая.
Другая серьезная проблема во многих районах плодоводства заключается в изобилии одуванчика и горчицы. Одуванчик обычно зацветает вместе с яблонями, а горчица отцветает вместе с яблонями. Некоторые наблюдения показывают, что медоносная пчела охотнее берет нектар с одуванчика и горчицы, что важно учитывать при размещении ульев в садах. В благоприятные годы при 5 ульях и 2 га яблоневого сада происходит настолько интенсивное опыление, что часть цветков приходится удалять, чтобы получить плоды нормального размера. В этом случае нужно ограничивать деятельность пчел определенным количеством часов или применять химические вещества для прореживания цветков.
Похожие работы
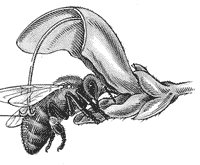
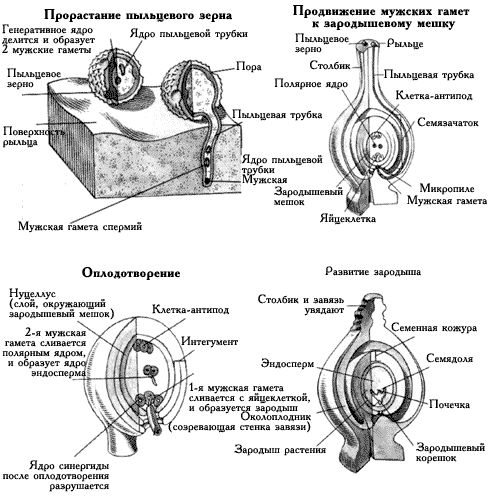
... лишь с развитием более тонкой, чем во времена Кельрейтера, микроскопической техники. Поэтому нет ничего удивительного, что в начале XIX в. первые успехи в раскрытии процесса оплодотворения у цветковых растений сделал не ботаник, а крупный специалист в области оптики и микроскопической техники, итальянский ученый, профессор Д.Амичи (1786–1863). Он заведовал в университете своего родного города ...
... ; - ствол, если он древесный и разветвленный, как у большинства деревьев; или же древесный, но не разветвленный, с листьями на верхушке, как у пальм. В зависимости от плотности стебля растения подразделяются на: - травянистые, у которых нежный, не одревесневший стебель; - полукустарниковые, у которых стебель одревесневает ствол только у основания; - кустарниковые, у которых все ветви ...
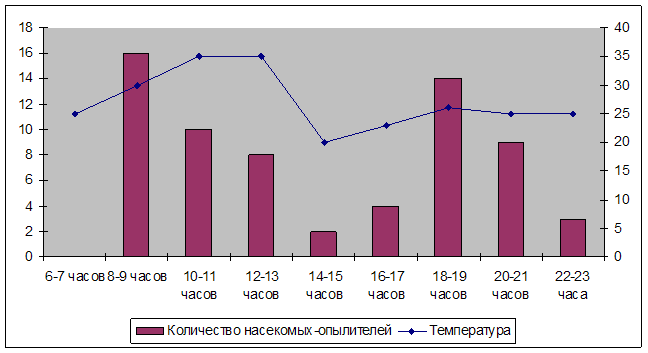
... четкой временной приуроченности видов ко времени опыления отмечено не было. В вечернее время основными опылителями растений были бражники. График №1 Суточная активность насекомых-опылителей на кальцефильной степи в Донском природном парке 4. Экологическая структура энтомонаселения Герпетобионты – насекомые, живущие среди растительных или иных органических остатков ...
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
0 комментариев