Актуальність. В епоху широкомасштабного впливу людської цивілізації на навколишнє середовище проблема вивчення закономірностей генезису, динаміки боліт і заболочених земель як важливого біоенергетичного компонента та носія значного обсягу біорізноманіття біосфери здобуває велике значення. Надзвичайно важливою є роль боліт у необоротному вилученні техногенної вуглекислоти з атмосфери. Поглиблені дослідження сучасної рослинності боліт, фізико-хімічних властивостей торфу й торф'яних покладів сприяють розкриттю цих закономірностей. Крім того, став можливою розробка наукових основ охорони та раціонального використання боліт. На теренах України переважають низинні болота з евтрофною рослинністю. Вони надзвичайно різноманітні за своєю природою, що пояснюється умовами водно-мінерального живлення.
Предмет дослідження – процеси, які лежать в основі утворення евтрофних боліт.
Об`єкт дослідження – рослинність евтрофних боліт.
Мета роботи – проаналізувати процес утворення евтрофних боліт та особливості їх рослинності.
Завданнями нашої роботи є:
1) розглянути формування евтрофних боліт;
2) дати поняття про евтрофну рослинність;
3) охарактеризувати основні типи евтрофних боліт;
4) розглянути особливості угруповань рослин, поширених серед основних типів евтрофних боліт.
Практичне значення. Дослідження евтрофних процесів сприяє більш повному розумінню впливу діяльності людини на виникнення евтрофних боліт.
1. Формування евтрофних боліт
Озерний тип болотоутворення досить розповсюджений в Україні. Озера Полісся льодовикового чи карстового походження. За час геобіогенної стадії, що настала після відступу льодовика і наступного потепління клімату, відбувалося обміління озер і заселення їх рослинами і тваринами, відмирання й опадання залишків яких призвело до утворення сапропелю потужністю 2–4 м, а на деяких озерах понад 8–10 м. Сапропель в елювенільній фазі змінювався торфом, частіше очеретяним, відкладеним очеретяними материнськими ценозами в умовах значного зволоження. Ця фаза в розвитку боліт тривала недовго і змінилася осоково-очеретяною, що відображала розвиток лісових боліт за низинним типом. На очеретяно-осоковому торфі при вступі в перехідну фазу матуративної стадії звичайно залягає осоково-очеретяно-сфагновий, а на ньому – осоково-сфагновий торф., що відкладався відповідними фітоценозами при меншому зволоженні і біднішому мінеральному живленні. Ця частина стратиграфії відображає розвиток лісових боліт за перехідним типом. У сучасному рослинному покриві переважають сосново-чагарничково-сфагнові і сосново-березово-осоково-сфагнові ценози.
В інших випадках на сапропелях залягає очеретяно-гіпновий чи деревно-очеретяний торф, що характеризують низинну фазу матуративної стадії розвитку боліт. Залягання останнього на сапропелі здається дивним, оскільки сапропель відклався у водоймах, де після їхнього обміління можна сподіватися на розвиток спочатку трав'яного чи трав'яно-мохового покриву. Зміна сапропелю деревно-очеретяним торфом відзначена й іншими авторами (Зеров, 1938). Очевидно, тут мало місце заростання водойми шляхом наростання осокової сплавини, на якій з її потовщенням з'являвся очерет і рідкі деревця вільхи. Досягши значної товщини, сплавина опустилася на дно водойми, покрите сапропелем.
Потому, в період перехідної фази матуративної стадії розвитку боліт деревно-очеретяний торф став субстратом для трав'яно-мохових чи де-ревно-трав'яно-мохових ценозів, які відклали драговинні чи лісодраговинні торфи, що стали субстратом для сучасних рослинних угруповань.
Одним зі шляхів розвитку лісових боліт є заторфовування відмерлих і сучасних водойм за допомогою утворення сплавини. Розвиток боліт починався з проювенільної і елювенільної фаз, частіше з гіпнової сплавини внаслідок відмирання й опадання її залишків, що утворили шари гіпнового торфу, котрий заповнював ложа водойм. Наростання торф'яного шару й наступне обміління водойм обумовили перехід до низинної фази матуративної стадії розвитку і сприяли зростанню спочатку очеретяно-гіпнових і очеретяних заростей, а при помірному зволоженні – деревно-очеретяно-гіпнових і деревно-гіпнових материнських ценозів, що відклали аналогічні торфи.
Відома роль гіпнового, очеретяно-гіпнового й осоково-гіпнового торфів у будові покладів лісових боліт. Місцями вони утворюють придонний шар покладу товщиною 0,25–1,25 м, рідше осоково-гіпновий торф залягає зверху, підтримуючи думку З.Н. Денисова (1951) й ГФ. Бачуріної(1964), що осоково-гіпнова фаза завершує розвиток низинного лісового болота.
У придонних шарах в проювенільній фазі розвитку лісових боліт частіше переважає вимогливий до умов існування Зсогрідіит зсогріоісіез, а в поверхневих шарах евтрофної фази матуративної стадії – рослини менш вимогливі до зольного живлення. Знаходження скорпідієвогс гіпнового торфу в придонних шарах покладів пов'язують із дрібними післяльодовиковими водоймами. Занурені мохи, що заповнювали водойми без утворення сплавини, стали початковою проювенільною фазою розвитку боліт, які сформувалися шляхом заростання водойм.
2. Рослинність низинних боліт. евтрофна рослинність
Такі болота розвиваються на низинних торфах і торфових покладах, наведемо дані про вміст поживних елементів у їх складі. Низинні торфи містять 1,82–3,46% азоту, 1,42–3,90% кальцію, 0,26–0,42% фосфору, 0,19–0,51% калію. їх зольність становить 5,63–12,35%, але часто вона буває вищою, особливо в торфів лісостепових районів; реакція середовища на болотах кисла (рН 3,78–5,90, а в лісостепових районах рН 7 і більше), місцями реакція фунтового розчину лужна. Подібні коливання вмісту найголовніших зольних елементів обумовлені значним поширенням боліт, що розвиваються на різних підстилаючих породах, відмінностями їх джерельного і алювіально-делювіального живлення. Високий вміст поживних речовин у торфовищі і доступність їх для рослин можнапояснити геоморфологічною приуроченістю боліт, їх тепловим і гідрологічним режимами протягом вегетації.
Болота, які розміщуються в заплавах річок, живляться багатими алювіальними водами, а болота, які розміщуються на других терасах, приозерних западинах, стічних улоговинах – делювіальними і джерельними водами. Крім того, всі геоморфологічні типи боліт додатково живляться ще й атмосферними опадами.
За умов багатого водно-мінерального живлення та періодичного підсихання низинні болота мають різноманітний видовий склад, добре виявлену горизонтальну і вертикальну почленованість та високу продуктивність. На них росте більше 200 видів квіткових і вищих спорових рослин. Багато з них знаходять тут оптимальні умови і нормально розвиваються. Однак у складі евтрофної рослинності та її флори відбувається диференціація видів за здатністю в цих умовах інтенсивно акумулювати і трансформувати енергію і речовину, створюючи специфічні групи болотних видів та їх фітоценотичні взаємозв'язки. Ці види є едифікаторами рослинних угруповань. Вони визначають будову та видовий склад кожного фітоценозу, ранг синтаксонів, за їх участі формуються певні асоціації, котрі становлять флористичне ядро синтаксона.
Залежно від ступеня зволоження, проточності і застійності болотних вод, на болотах розвиваються різні типи рослинного покриву. Мало обводнені болота вкривають лісові болотні угруповання. Дуже обводнені болота населяють рідколісні угруповання з низькорослим і пригніченим деревостаном або крупнотравні безлісні болотні угруповання – очерету, рогозу, схенусу озерного тощо. Менш зволожені болота зайняті трав'янистими безлісними угрупованнями (при проточному водно-мінеральному живленні), або трав'янисто-гіпновими чи трав'янисто-сфагновими угрупованнями з суцільним моховим покривом (при застійному водному живленні). Особливістю останніх типів рослинних угруповань є не тільки домінування, але й зміна фітоценотичної ролі мохового покриву, який у даному випадку виступає як головний едифікатор рослинних угруповань, що визначає суть і природу еволюції болотних синтаксонів.
Залежно від фітоценотичної ролі певної життєвої форми рослин у складі евтрофної рослинності виділяють лісові, чагарникові, трав'янисті і трав'янисто-мохові підтипи та їх угруповання. Перші із них нерідко називають лісоболотними або закритими болотними угрупованнями, тимчасом як трав'янисті і трав'янисто-мохові та мохові підтипи кваліфікують як відкриті болота. В Україні переважають відкриті евтрофні болотні угруповання.
Лісові болота – це надмірно зволожені ділянки поверхні Землі зі специфічною болотною (гелофітною) рослинністю, добре розвинутим деревостаном із зімкнутістю крон не менше 0,3, шаром торфу не менше 0,5 м, насиченим кореневими системами лісоутворюючих порід.
Постійно або тимчасово надмірно зволожені ділянки поверхні зі специфічною болотною рослинністю та шаром торфу менше 0,3 м в неосушеному стані або без нього називають заболоченими землями. Вони з розвинутим дерево станом і зімкнутістю крон 0,3 – 0,1 утворюють заболочені ліси і являють собою одну із стадій формування лісових боліт. Залежно від участі у формуванні синтаксонів лісоутворюючих порід у складі лісових боліт виділяють три групи – хвойнолісову, листянолісову та хвойно-листянолісову.
3. Загальна характеристика основних видів евтрофних боліт та їх рослинності 3.1 Лісові болота
Лісові болота листянолісової групи включають угруповання, в яких переважають листянолісові лісоутворюючі породи, насамперед Alnus glutinosa, A. incana, Betula pubescens, B pendula, Populus tremula, Fraxinus excelsior, Salix alba, S. fragilis
Вільхові лісоболотні угруповання досить поширені на Поліссі, де їх фітоценози займають близько 435 тис. га. Багато їх у Придніпров'ї із Східному Поліссі, де вони є характерною ознакою ландшафту, менше – у Лісостепу, там ці фітоценози приурочені головним чином до долин річок, межиріч, навколо озер, ставків, біля джерел, у притерассі.
Основною лісоутворюючою породою є Ainus giutinosa, менше поширені угруповання A. incana, які частіше трапляються на болотах західних регіонів.
Характерною рисою вільхових боліт є розчленування їх поверхні на мало зволожені пристовбурні горби («п'єдестали») і дуже зволожені або обводнені зниження між ними. Подібна диференціація поверхні визначає еколого-ценотичні відмінності цих боліт, насамперед гідрологічний режим, характер живлення і диференціальний структурний розподіл флористичного складу ценозів.
По надмірно обводнених болотах з поділом поверхні на пристовбурні горби та зниження під наметом деревостану чагарниково-трав'янистий і моховий покрив розподілені нерівномірно. На підвищеннях (до 1,5 м) поселяється Alnus glutinosa з домішкою Betula pubescens, Pinus sylvestris і Picea abies. Тут же ростуть кущі Padus avium, Ribes nigrum, Rubus idaeus, Viburnum opulus. У трав'яно-моховому покриві ростуть болотні, лучні і лісові види: Oxalis acetosella, Majanthemum bifolium, Lycopus europaeus, Dryopteris carthusiana. Thelypteris palustris, Climacium dendroides, Sphagnum palustre, S. squarrosum, S. teres. Обводнені зниження, де тривалий час стоїть вода навіть улітку, займають Salix cinerea і осоки: Сагех гірагіа, С. elatar C. acuti-formis, C. appropinquata, C. elongata. Місцями часто трапляються Filipendula denudata, Menyanthes trifoliata, Hottonia palustris, Comarum palustre.
За складом лісоутворюючих порід і структурою деревостану, домінантною участю чагарниково-трав'янисто-мохових видів, режимом зволоження і проточністю води розрізняють екологічні ряди, які включають окремі групи асоціацій. На болотах, які живляться швидкопроточними водами, алювіальними і джерельними, розвиваються вільхово-гадючникові, вільхово-розривтравові, вільхово-кропивові та вільхово-малинові асоціації. Вони утворюють екологічний ряд, котрий з'єднує торфові лісові болота і ліси на мінеральних ґрунтах.
Із погіршенням проточності води, шаруватості ґрунту та зниженням мікробіологічної активності торфовища розвиваються вільхово-папоротеві, вільхово-осокові і вільхово-очеретяні асоціації. Цей екологічний ряд з групами асоціацій характеризується середньою проточністю води і помірним мінеральним живленням, що призводить до деякого зрідження деревостану і зниження його продуктивності. На таких болотах центральне положення займають вільхово-осокові угруповання. Вони неоднорідні за ступенем росту лісоутворюючих порід, структурою та складом деревостану, а також за домінантною участю видів трав'яно-мохового покриву і представлені численними асоціаціями. їх різноманітність обумовлена не стільки рівнем зольного живлення, скільки режимом зволоження.
У структурі ценозів вільхових боліт найбільш зволожені вільхово-очеретяні угруповання. Вони менш поширені, ніж вільхово-осокові, і не займають великих площ. Частіше трапляються в Лісостепу, рідше на Поліссі. За умов високого стояння води деревостан вільхи розвинутий мало. У віці 40–60 років дерева досягають 16–18 м заввишки та діаметра стовбурів 14–40 см. Стовбури збіжисті, з погано виявленою кроною. Зімкнутість крон деревостану 0,5–0,8. Бонітет III–IV, рідко II. До вільхи домішуються у невеликій кількості Betula pubescens і сосна.
Характерною ознакою цих угруповань є домінування в трав'янистому покриві Phragmites australis, участь якого в проективному покритті коливається від 20–30% у високозімкнутих деревостанах до 60–80% у малозімкнутих. Густота стеблостояння, затіненість, висока насиченість вологою ґрунту і застійність або мала проточність води пригнічують розвиток травостою, він розріджений і слабо диференційований. Найчастішими супутниками очерету є: Lysimachia vulgaris, Lycopus europaeus, Solarium nigrum, Impatiens noli-tangere, Galium palustris, Filipendula denudata, Thelypteris palustris, Calamagrostis canescens, Peucedanum palustris. Разом з мезофільними лісовими мохами (Thuidium philibertii, Abietinella abietina, Fissidens adianthoides, Polytrichum commune, Climacium dendroides, Mnium undulatum) вони заселяють підвищені пристовбурні горби, а на обводнених або надмірно зволожених ділянках поміж ними ростуть Iris pseudacorus, Hottonia palustris, Oenanthe aquatica, Ranunculus lingua, Calla palustris, Naumburgia thyrsiflora, Carex rostrata, C. riparia, C. acutiformis, Equisetum fluviattle.
При застійності вод і меншій обводненості в наземному покриві виділяються синузії гідрофільних мохів – Drepanocladus fluitans, D. verni-cosus, D. aduncus, Calliergon giganteum, Calliergonella cuspidata, Tomenthypnum nitens,
Малозволожені або тимчасово надмірно зволожені болота вкриваються вільхово-осоковими угрупованнями з домінуванням у травостої Carex acutiformis, C. riparia, C. vesicaria. Деревостан має добрі таксаційні показники, звичайно росте за ІІ – ІІІ бонітетом. Підлісок розвинений слабо. Подекуди трапляються кущі Rubus idaeus, Viburnum opulus, Frangula alnus, Ribes nigrum. У наземному покриві у невеликій кількості ростуть типові болотні види: Ins pseudacorus, Phragmites australis, Galium palustre, Ranunculus lingua, Thelypteris palustris та ін.
На дуже зволожених болотах розвиваються вільхово-осокові ліси з домінуванням Carex elata та С. appropinquata. Деревостан НІ бонітету. Характер підліску та травостою подібний до описаного вище, але в цих асоціаціях на пристовбурних підвищеннях рясно ростуть Ribes nigrum, Urtica dioica, Oxalis acetoseila, Impatiens noli-tangere, а по зниженнях Menyanthes trifoliata, Comarum palustre, Filipendula denudata.
При слабопроточних і застійних водах, погіршенні аерації та мінерального живлення розвиваються вільхово-папоротево-гіпнові, вільхово-очеретяно-гіпнові, вільхово-осоково-гіпнові та вільхово-осоково-сфагнові асоціації. їх особливістю є рясний розвиток гіпнів (Calliergon giganteum, Calliergonella cuspidata, Drepanociadus aduncus) і сфагнів (Sphagnum palustre, S. squarrosum, S. teres). Внаслідок сповільненого стоку води у весняно-осінній період та через наявність у наземному покриві гіпнових і сфагнових мохів розвиток деревостану погіршується, а кількість видів скорочується. Бонітет деревостану вільхи знижується до IV класу. Цей екологічний ряд знаменує подальший ступінь мезотрофності вільшняків і зближення їх з перехідними лісовими болотами.
Березові лісові болота в Україні поширені головним чином на Поліссі, насамперед на Центральному та Західному Поліссі. Менше покриті лісоболотними березовими ценозами болота Північного Лісостепу. Вони розташовані по зниженнях других терас і межиріч, зрідка приурочені до проточних улоговин і долин річок, які періодично затоплюються повеневими водами.
Березові болота неоднорідні за продуктивністю і флористичним складом. Тут росте близько 100 видів квіткових і вищих спорових рослин. їх взаємопоєднання з лісоутворюючою породою та екологічними факторами місцезростання обумовили формування різних груп асоціацій. Найхарактернішими з них є березово-осоково-гіпнова з березово-дернистоосоково-гігантськокалійергоновими угрупованнями. При малому зволоженні зі слабкою проточністю ґрунтових вод або при деякому природному підсиханні (болото Татінов) утворюються високопродуктивні деревостзни. Висота берези становить 16–18 м, діаметр стовбурів 14–22 см, зімкнутість крон 0,6–0,8, бонітет II, запас деревини 140 м3/га. При більшому зволоженні (болота Довге Поле і Шабелька) утворюються малопродуктивні деревостани. Тут висота берези не перевищує 10 м, діаметр стовбурів 10–14 см, зімкнутість крон 0,6, бонітет IV, запас деревини 60 м3/га. У травостої домінують Carex caespitosa та С. appropinquata, які утворюють купини заввишки до 40 см. Невелика кількість у травостої Carex rostrata, C. acuta таC. elata, Agrostis stolonifera, Calamagrostis canescens, Thelypteris paiustris, Menyanthes trifoliata, Comarum palustre. У моховому покриві переважає Calliergon giganteum, Calliergonella cuspidata, Drepanocladus aduncus.
Рідше трапляються березово-куничниково-гіпнові, березово-папоро-тево-гіпнові і березово-різнотравно-гіпнові болота. На давно осушеному болоті Смердюче виявлено дуже рідкісне березово-осоково-гіпнове угруповання з домінуванням Carex brizoides, властивої лісовим угрупованням. Це угруповання заслуговує на охорону і вивчення фітоценотичних зв'язків, які виникають під впливом осушення лісових боліт.
Осикові лісові болота належать до дуже рідкісних болотних угруповань не тільки в Україні, але й на теренах інших країн СНД. На болотах Старе Гало і Смердюче (Білокоровицький лісгоспзаг Житомирської обл.) висота Populus tremula за оптимальних умов водно-мінерального живлення досягає 22 м, діаметр стовбурів 20–28 см, зімкнутість крон 0,6, бонітет 1–11. На торфовищах потужністю до 4,0 м, що живляться слабопроточними водами, розвиваються осиково-осоково-гіпнові, а на болотах із застійним зволоженням – осиково-осоково-сфагнові угруповання, які за характером трав'янисто-мохового покриву наближуються до перехідних лісових боліт.
Д.К. Зеров (1938) вперше в Україні описав осиково-березово-осоково-гіпнові і осиково-березово-осоково-сфагнові угруповання на болоті Корма (Олевський лісгоспзаг Житомирської обл,).
На цьому болоті росте така рідкісна для України рослина, як Chamaedaphne calyculata. Це невеличкий кущик із довгасто-еліптичними шкірястими листочками і звисаючими однобічними китицями красивих жовто-кремових квіточок. Квітки і сама рослина мають дуже привабливий вигляд, а тому варто оцінити можливості введеня її у культуру зеленого будівництва. Природний ареал хамедафни проходить на півночі Білорусі, Прибалтики, Російської Федерації та прилеглих територіях. Там вона домінує під наметом деревостану, а на Поліссі має острівне поширення. Chamaedaphne calyculata, як і осикові угруповання, потребує повної охорони.
Хвойно-листянолісові лісоболотні угруповання досить поширені у поліських і лісостепових районах України. Ценози цих угідь являють собою подальший ступінь в напрямку мезотрофності і характеризують завершальні стадії евтрофності та перехід до мезотрофності умов місцезростання групи, що знаходить своє відображення в характері розвитку рослинного покриву, У складі цієї формації виділяється ряд формацій, найголовніші з яких будуть охарактеризовані нижче.
Сосново-вільхові лісові болота також досить поширені на Поліссі і зустрічаються в комплексі з іншими болотами. Частково вони поширені на болотах Північного Лісостепу. Нами такі формації описані на 14 болотах Західного та Східного Полісся. їх особливістю є поєднання в деревостані порід різної екологічної вимогливості – Alms glutinosa, що потребує проточної води, доброї аерації та багатого мінерального живлення, і Pinus sylvestris, яка витримує тривале застійне зволоження і менш вибаглива щодо аерації та зольного живлення. На болотах Локницьке і Городецьке (відповідно Зарічанський і Дубровицький лісгоспзаги Рівненської обл.) за оптимальних умов висота вільхи та сосни досягає 14–18 м, діаметр стовбурів 12–22 см, зімкнутість крон 0,8, бонітет III–IV В складі деревостану панує вільха, для якої тут, ймовірно, склалися кращі умови, ніж для сосни.
Іншою характерною рисою сосново-вільхових боліт є майже суцільний моховий покрив із Sphagnum palustre, S. fallax, S. centrale і S. girgensohnii. Наявність останніх свідчить, що ці болота вступили в мезотрофну стадію розвитку. Індикаторами мезотрофності служать такі види чагарниково-трав'яного ярусу, як Oxycoccus paiustris, Carex lasiocarpa, C. rostrata, Menyanthes trifoliata, Molinia coerulea, Andromeda polifolia та ін.
Сосново-вільхові болота потребують охорони. Особливої уваги серед них заслуговують болота Зарічина та Високий Мох (Володими-рецький лісгоспзаг Рівненської обл.). Тут на торфовищах потужністю 1,5–2,5 м лісоутворюючі породи досягають висоти 20–26 м. Це найкращі лісоболотні насадження не тільки на Поліссі, а й взагалі в Україні. Зони можуть бути еталонами створення високопродуктивних насаджень на осушуваних болотах.
Вільхово-ялинові болота мають обмежене поширення. Ці болотні ценози займають невеликі ділянки. Нами вони були описані на болотах Черемоське і Дитині Грудки (відповідно Городотоцький і Ратнівський лісгоспзаги Волинської обл.), а також на болоті Визбиця (Зарічнянський лісгоспзаг Рівненської обл.). Ці болота покриті вільхово-ялиново-трав'янисто-сфагновими угрупованнями з домінуванням у травостої Carex riparia, Calamagrostis canescens, Thelypteris paiustris, а на одній з ділянок Vaccinium mytrillus.
Деревостаи одноярусний, часом з домішкою Betuia pubescens. Склад деревостану 2Я8В-4Я6ВБ, висота 18–22 м, діаметр стовбурів вільхи 20–28 см, ялини – 20–36 см, зімкнутість крон 0,7–0,9, бонітет МІ.
Одне з південніших місцезростань вільхово-ялинових болотних лісів в Україні відмічене Є.М. Брадіс (1969) у Дубнівському лісгоспзагу Рівненської обл.
Вперше в 1974 р. описано вільхово-ялиново-осоково-сфагнові угруповання на болоті Тички (Олександрівське лісництво Добрянського лісгоспзагу Чернігівської обл.). Глибина залягання торфу 0,5–1,0 м. Склад деревостану 8В2Я-6В4Я. Висота дерев 18–24 м, діаметр стовбурів вільхи 14–20 см, ялини 18–30 см, зімкнутість крон 0,8, бонітет ІІ-ІІІ. В підліску трапляються окремі кущі Frangula alnus і Salix cinerea. Домінантами чагарничково-трав'янистого покриву є: Carex elongata і С. caespitosa. З покриттям 1–10% тут представлені: Carex vesicaria, Caltha paiustris, Calla paiustris, Lysimachia vulgaris, Menyanthes trifoliata, Dryopteris carthusiana, Galium palustre, Viola paiustris та ін. Ці види властиві вільховим болотам іприурочені до пристовбурових горбів вільхи або ростуть по надмірно зволожених зниженнях поміж ними. На пристовбурних підвищеннях ялини ростуть Lysimachia vulgaris, Majanthemum bifolium, Trientalis europaea, Rubus saxatilis, Vaccinium rnyrtilius, Oxalis acetoseiia.
На ділянках, залитих водою шаром до 20 см, у наземному покриві виділяються окремі синузії гіпнових і сфагнових мохів. В умовах незначного зволоження домінують сфагнові мохи Sphagnum palustre, S squarrosum, S. girgensohnii.
Ці рідкісні рослинні угруповання на Українському Поліссі потребують охорони і збереження їх як пам'яток лісових боліт.
Вільхово-ялиново-березові болота також досить рідкісні для Полісся України. На болоті Черемоське (Городоцький лісгоспзаг) вперше описано вільхово-ялиново-березово-трав'янисто-мохову асоціацію з домінуванням у травостої Thelypteris palustris і Hottonia palustris, а в моховому покриві Caliergonella cuspidata і Sphagnum squarrosum. Від попередніх ці болота відрізняються наявністю в складі деревостану Betula pubescens. Залежно від ступеня зволоженості її участь змінюється від 4Я4В2БС до 2Я6Б2В. За флористичним складом і продуктивністю деревостани схожі на вільхово-ялинові, з яких розвиваються внаслідок зменшення ступеня зволоження і збіднення мінерального живлення.
Соснові лісоболотні евтрофні болота поширені досить мало. Низинні соснові лісові болота трапляються спорадично по зниженнях других терас і межиріч. На болоті Кривуха (Городоцький лісгоспзаг Рівненської обл.) описано рідкісне низинне сосново-сфагнове угруповання Pinetum sphagnosum. Деревостан одноярусний. За оптимальних умов середньопро-точного зволоження, доброї аерації та відносно багатого мінерального живлення висота Pinus sylvestris досягає 10–16 м, діаметр стовбурів 14–20 см, зімкнутість крон 0,6, бонітет IV, запас деревини 90 м3/га. На болотах із застійним зволоженням (Грузке, Густі Острови) і низьким рівнем мінерального живлення висота деревостану не перевищує 6 м, діаметр стовбурів 6–10 см, зімкнутість крон 0,3–0,4, бонітет Va, запас деревини 40 м3/га. У складі цих боліт виділяються сосново-папоротево-гіпнова, сосново-осоково-гіпнова і сосново-сфагнова групи асоціацій з домінуванням у травостої Carex appropinquata, Thelypteris palustris, а в моховому покриві – Caliergon giganteum, Caliergonella cuspidata, Sphagnum palustre.
Ялинові болота поширені лише на Поліссі і мають острівний характер поширення. Ще рідше трапляються на болотах. Найбільш характерними є ялиново-чагарничково-мохові угруповання, які були описані на болоті Кривуха (Маневицький р-н Волинської обл). Потужність торфовища 1,5 м. Висота ялини 16–18 м, діаметр стовбурів 12–20 см, зімкнутість крон 0,9. До ялини домішуються Pinus sylvestris, Betula pubescens, Alnus glutinosa, Poputus tremula, Frangula alnus, Salix cinerea. Чагарничково-трав'яний покрив слаборозвинутий. У ньому переважають Vaccinium mytrillus, Oxalis acetoseiia. З покриттям 1–5% тут ростуть: Carex lasiocarpa, Phragmites australis.
Моховий покрив (75%) утворюють болотні сфагнові та лісові гіпнові мохи. Типовими тут є: Pleurozium schreberi, Polytrichum commune, Dicranum undulatum, Climacium dendroides, Sphagnum palustre, S. acuti-formis, S. squarrosum.
Всі лісоболотні угруповання з участю ялини унікальні та дуже рідкісні на болотах Полісся. Ялина належить до бореальних видів і суцільні масиви ялинових лісів утворює північніше на території Білорусі, Російської Федерації та в гірських районах Карпат. На Поліссі вона має острівний ареал, тому зустрічається тут звичайно на найбільш зволожених і холодних ділянках, що відповідає еколого-географічній природі цієї лісоутворюючої породи.
Безумовно, ялинові лісові угруповання потребують охорони і вивчення можливостей для того, щоб створювати штучні насадження з участю ялини в північних районах Полісся.
3.2 Чагарникові болотаЗа останні десятиріччя площ з чагарникових боліт помітно зростає, оскільки значно зменшено сінокосіння на болотах, а також розчищення сінокосів і пасовищ від чагарників. Ці рослинні угруповання часто трапляються на долинах малих річок, де малі притоки і верхів'я річок майже повністю покриті чагарниками. їх окремі ділянки трапляються на заплавних, долинних і староруслових болотах, часто вони повністю покривають торфорозробки.
До чагарникових боліт відносять болота, в яких чітко виділяється ярус чагарників, а їх зімкнутість становить не менше 0,3 або 30% загального покриття. Болота зі ступенем зімкнутості 0,1–0,2 належать до рідкочагарникових. їх особливістю є домінування гелофітів у трав'янисто-моховому покриві і насиченість поверхневого шару торфу коренями чагарників.
За характером домінант – це угруповання Salix cinerea, Salix caprea, зрідка трапляються S. triandra i S. pentandra, зовсім невеликими ділянками – крушинові угруповання з Frangula alnus. У більшості випадків чагарникові болота дуже обводнені.
Листопадні чагарникові болота об'єднують вільховочагарникові, та угруповання Saiix cinerea.
Угруповання Salix cinerea. За оптимальних умов гідрологічного режиму та мінерального живлення Salix cinerea досягає висоти 3–4 м, а за гірших умов 1,5–2,5 м. У разі відсутності обмежуючої дії кущі швидко розростаються і вкривають болота, утворюючи суцільні густі зарості. Щільність пагоностояння спричинює те, що під їх пологом росте обмежена кількість видів. Тому такі чагарникові угруповання флористично бідні і фізіономічно одноманітні. Багатші та різноманітніші в цьому плані угруповання з розрідженою зімкнутістю чагарників. За домінуванням у флористичному складі трав'янистих і видів мохового покриву виділяються вербово-очеретяні, вербово-осокові, вербово-різнотравні, вербово-злаково-осокові та вербово-осоково-гіпнові угруповання.
На поліських і особливо лісостепових болотах з високим ступенем обводнення трапляються лозово-очеретяні угруповання, в яких участь Phragmites australis досягає 50–70%. Щільність стеблостояння чагарника й едифікатора травостою обмежує поширення інших болотних видів. Найчастіше з високим ступенем сталості тут трапляються Oenanthe aquatica, Solanum dulcamara, Galium palustre, Ranunculus lingua, Lythrum salicaria, Rumex hydrolapathum, Valeriana stolonifera, Peucedanum palustris, Rorippa amphibia.
В умовах меншого обводнення, але періодичного затоплення поширені (особливо вздовж річок) лозово-купинястоосокові угруповання, в яких основними домінантами в чагарниковому ярусі виступають Salix cinerea із зімкнутістю крон 0,4–0,7, а в травостої Carex eiata, яка утворює купини до 50 см заввишки, котрі займають до 40% поверхні болота. Проміжки між ними заповнені водою і заселені гідрофільними видами рослин (Comarum palustre, Lythrum virgatum, Naumburgia thyrsiflora, Lysimachia vulgaris, Equisetum fluviatile. Calla palustris, Mentha arvensis, Geum rivale).
Із зниженням обводеності і за періодичного підсихання в травостої збільшується участь дрібних осок, болотного різнотрав'я та злаків. Тому тут виникають лозово-кореневищноосокові угруповання, в яких істотну роль відіграють Carex vesicaria, C. acuta, C. nigra, C. flava. При цьому чагарниковий ярус більш розріджений і низькорослий (1,5–2,5 м), його зімкнутість становить 0,3–0,5. За таких умов проективне покриття травостою збільшується до 60–80%. Осоки домінують з загальним покриттям 50–60%.
На підсихаючих або антропогенно порушених болотах, особливо на тих, де проводилось систематичне сінокосіння та випасання, у травостої зростає роль злаків, а тому виникають лозово-осоково-злакові угруповання, в яких злаки співдомінують з осоками. До вже згаданих осок домішуються Agrostis stolonifera, Calamagrostis canescens, Glyceria fluitans (20–45% загального покриття). З різнотрав'я часто трапляються Ranunculus flammula, R. repens, Myosotis palustris, Galium palustre, Gnaphalium ultgi-nosum, Sagina nodosa, Prunella vulgaris, Thelypteris palustris.
Вільхові чагарникові болота мають спорадичне поширення. Частіше трапляються на болотах Західного Лісостепу, рідше – на болотах в інших регіонах. Вони виникли на місці зведених вільхових або вільхово-березових лісів чи внаслідок самосіву вільхи, проте через антропогенний вплив (насамперед використання цих боліт як пасовищ, сінокосів тощо) не досягли розвитку жерняків або лісових насаджень. Нерідко з'являються на місці торфорозробок, у заплавах річок, особливо у дуже заболочених верхів'ях малих річок та струмочків.
Розвиваються вільхові чагарникові болота на торфовищах різної потужності (від 0,5 до 5 – 7 м завглибшки) часто трапляються на заболочених землях з потужністю торфового шару 0,3–0,5 м, тобто на торф'янисто-глейових або торф'яно-глейових чи мулуватих ґрунтах. В останньому випадку їх поверхня вирівняна, майже без купин, тимчасом як на торфовищах в міру зростання потужності торфового шару зростає висота пристовбурних підвищень у чагарників пневого походження, однак у чагарників насіннєвого походження ця закономірність не спостерігається (такі підвищення в них менших розмірів).
Через диференціацію поверхні, на якій ростуть вільхові чагарники, вони набувають різного ступеня зволоження. Що стосується гідрологічних вимог едифікатора, то він віддає перевагу проточним водам; за цих умов частіше формуються жердняки, які трансформуються в лісові угруповання різного ступеня розвитку з різними еколого-ценотичними взаємозв'язками.
Вільхові чагарникові болота досить різноманітні у флористичному і ценотичному відношенні. Вільхові кущі за оптимальних умов проточності фунтових вод і багатства водно-мінерального живлення досягають висоти 2–3 м, а подекуди 4–5 м. Гірше вільха розвивається в умовах застійності води на улоговинних болотах, тут вона так і залишається на чагарниковій стадії розвитку. За приуроченістю найбільш поширені притерасні болотні угруповання. Тут вони і найрізноманітніші, що обумовлює флористичний склад та його асоційованість з едифікатором.
На лісостепових болотах досить часто трапляються вільхово-очеретяні угруповання, де співдомінантом виступає Phragmites australis, котрий при обводненні міжкупинних проміжків досягає висоти вільхи, а його проективне покриття становить 40–50%. Менша участь тут інших видів рослин.
Часто трапляються вільхово-осокові угруповання з домінуванням у травостої Carex elata (30–40%), С. acuta (20–40%), С. vesicaria (20–35%) та ін. При цьому спостерігається, з одного боку, розрідженість чагарникового ярусу, а з іншого – зростання видової насиченості рослинних угруповань видами лучно-болотної екології.
Подальше зрідження чагарникового ярусу до зімкнутості 0,3–0,4 та поступове підсихання або осушення боліт спричинюють розвиток вільхово-злакових і вільхово-злаково-різнотравних ценозів неоднорідної екології та асоційованості видів. Із співдомінантів тут поширені Agrostis stolonifera, Glyceria fluitans, Poa palustris.
Зрідка трапляються вільхово-папоротеві угруповання з домінуванням у трав'янистому ярусі Thelypteris palustris, рідше – Dryopteris filixmas та Equisetum palustre.
3.3 Трав'янисті болотаНа низинних болотах Полісся та Лісосепу переважають відкриті трав'яні рослинні угруповання. В їх сучасному складі виділяються дві групи: природні болотні угруповання, що розвиваються на неосушених або вторинно заболочених болотах, та штучні болотні угруповання, які сформувалися на осушених болотах, новоутворені або природновідновлювані в процесі постмеліоративних змін.
Трав'яні болотні угруповання характеризуються рядом еколого-ценотичних, біологічних і господарськи цінних ознак, а саме:
– трав'яні болота не мають будь-якого виявленого деревного або чагарникового ярусу. Лише на осушених болотах уздовж меліораційної мережі трапляються поодинокі дерева Alnus glutinosa або Betula pubescens;
– всі трав'яні угруповання утворені виключно трав'янистими видами квіткових або вищих спорових рослин, котрими визначаються ценотичні взаємозв'язки між ценоелементами, їх фізіономічність, структурна почленованість і продуктивність;
– оптимальні умови для свого розвитку трав'янисті види знаходять на долинних, староруслових, заплавних, улоговинних болотах з проточними, хоча б тимчасово болотними водами;
– домінуючими едифікаторами травостою є осоки і злаки, меншою мірою види болотного різнотрав'я, хвощі та папороті. їх щільність та фітомаса є лімітуючими факторами, якими визначається взаємозв'язок як між компонентами фітоценозів, так і між останніми та торфовим субстратом з його тепловим і гідрологічним режимом;
– проточність ґрунтових вод, алювіальність і делювіальність обумовлюють те, що трав'яні ценози бідні за видовим складом, малоструктуровані, мають слаборозвинутий моховий покрив.
Високотравні болота є складі лісостепових боліт досить поширені. Менше їх на болотах Полісся. Такі угруповання приурочені до найбільш знижених і обводнених ділянок болотних масивів. Це здебільшого повністю заболочені заплави малих річок з незадовільним стоком води, тальвеги балок, стариці, старі русла річок, заростаючі ставки, приозерні ділянки, на яких надовго застоюється повенева, дощова або тала вода. Лише в роки з дуже жарким літом вони місцями пересихають.
Високотравні угруповання специфічні, вони мають свої особливості екології та асоціювання. На нашу думку, це достатня кількість тепла, вологи, елементів зольного та азотного живлення. Площі високотравних угруповань збільшуються в напрямку з півночі на південь, де в період вегетації температура підвищується до 30 °С і більше, добре прогріваються болотні води, а їх випаровування сприяє зростанню вологоємності малорухливого повітря всередині фітоценозів. У лісостеповій зоні, де підстилаючими грунтоутворюючими породами є леси, болота краще забезпечені зольним І азотним живленням, ніж на Поліссі. У зв'язку з цим тут склалися оптимальні умови для формування високотрав'я, до складу якого входять Phragmites australis, Typha latifolia, T. artgustifolia, Scirpus lacustris, Glyceria maxima, Acorus calamus, Sparganium recta, а місцями крупні осоки та види різнотрав'я.
Характерною особливістю високотравних угруповань є моно домінантність у більшості випадків їх основним едифікатором виступає високо-травний вид із загальним покриттям ценозів 50–90%. Саме вони й визначають структуру болотних угруповань. Як правило, такі ценози одноярусні, рідше двох- і багатоярусні, вони мають бідний флористичний склад і низьку видову насиченість, спрощену одноярусну будову. Кількість видів у ценозах збільшується при зниженні обводненості рослинних угруповань. Мала вертикальна і горизонтальна почленованість високотравних угруповань, мабуть, обумовлена одноманітністю травостою та вирівняністю поверхні місцезростання, його екологічною однорідністю.
Таким чином, високотравні види утворюють своєрідну ценотично і фізіономічно відокремлену групу болотних видів рослин, котрі, асоціюючися між собою, формують досить стійкі трав'янисті комплекси, які визначають їх продуктивність та господарську цінність.
3.3.1 Очеретяні угруповання
Очеретяні угруповання в Лісостепу (менше на Поліссі) найхарактерніші для високотравних фітоценозів Звичайно ці угруповання мають ряд особливостей. По-перше, серед усіх високотравних угруповань вони відзначаються найбільшою висотою (3–4 м, а подекуди й більше), по-друге, їм притаманна найменша як горизонтальна, так і вертикальна почленованість; по-третє, очеретяні угруповання очерету мають просту структуру, але вони різноманітні фітоценотично.
Вони дуже поширені на заплавних болотах-багатьох річок і їх приток, по малодіяльних руслах малих річок і заболочуваних заплавах, по старицях і заростаючих ставках, вироблених торфовищах. Тут завжди високий рівень стояння води і підтоплення очеретяних заростей. Phragmites australis за умов високого обводнення (1,0 – 2,0 м), достатнього мінерального живлення, яке забезпечується за рахунок джерельних вод, нормального температурного режиму протягом тривалого часу досягає максимальної висоти.
Висота очерету, його облисненість, щільність стеблостояння зумовлюють своєрідний тепловий, світловий і температурний режими фітоценозу, від якого значною мірою залежать флористичний склад, структурна організація та продуктивність травостою. З висотою, ступенем затіненості і водозабезпеченістю пов'язані поширення та розподіл видів різних екологічних груп рослин очеретяних угруповань різного синтаксономічного рангу. Phragmites australis на постійно та тривало обводнених ділянках з пухким субстратом торфу і особливо у водоймах, утворює довгі, міцні, густо переплетені між собою кореневища, котрі за вегетаційний період дуже розростаються, і на відкритих водоймах разом з іншим болотним різнотрав'ям він стає основою сплавини. Очерет інтенсивно розростається в ширину, а також активно заглиблюється в мулистий чи торфовий грунт на глибину 1,5–2,5 м. Унаслідок відмирання і наступної неповної мінералізації його решток утворюється основна маса торфу, який складає потужні пласти однойменних очеретяних покладів, що виповнюють ложе боліт.Із морфолого-анатомічними та біологічними особливостями очерету взаємопов'язані асоціативність, життєвість, частота трапляння, кількісні та якісні ценотичні показники. Особини з добре розвинутою гідроморфною структурою приурочені до водойм з проточною водою, а особини з гігроморфною структурою переважають на заболочених або періодично підсихаючих болотах. У першому випадку очерет домінує в травостої, а в другому – поділяє участь зі співедифікаторами, виявляючи таким чином взаємозбалансовані ценотичні властивості компонентів ценозів.
Очеретяні угруповання є прикладом монодомінантних одноярусних угруповань, що сформувалися в результаті одночасного і дружного проростання діаспор, інтенсивного розвитку бруньок відновлення кореневищ. За цих умов утворюються густі однорідні зарості. Завдяки щільності стеблостояння рослини інтенсивніше ростуть у висоту. Ця тенденція притаманна всім високотравним видам угруповань.
У високотравних очеретяних угрупованнях створюється специфічний мікроклімат: тут повітряні маси малорухливі, температура коливається менше і трохи нижча, ніж над травостоєм, вище насиченість вологою, оскільки високий рівень стояння болотних повільно рухливих вод забезпечує високу випаровуваність, а отже насиченість фітоценозу водяною парою. Все це, безумовно, позначається на розвитку фітоценозу та його ценотичних властивостях.
Високотравність та специфічність очеретяних угруповань ценотичних і екологічних умов їх місцезростання є причиною того, що вони мають бідний і одноманітний видовий склад. Основним едифікатором таких рослинних угруповань виступає Phragmites australis. Крім того, як свідчать дані обстеження, у 21 фітоценозі звичайноочеретників ростуть 72 види квіткових і вищих спорових рослин, у тому числі злаків – 10 видів, бобових – 1, осокових -10, ситникових – 2, хвощів – 2, папоротей – 2 і різнотравних – 45 видій. Моховий покрив розвинутий слабо, в ньому відмічено 8 видів гіпнових мохів. Слід відзначити, що флористичний склад інших високотравних фітоценозів даної формації ще бідніший, хоча кількість видів у цих фітоценозах зростає під впливом проведення меліоративних та інших робіт.
У ценотичному аспекті угруповання Phragmites australis також досить сталі і одноманітні: у фітоценозах налічується 11–35 видів, а частіше 13–21 вид, що значно менше, ніж в осокових ценозах.
Серед співдомінантів Phragmites australis ми відмітили 7 видів: з них із сталістю 80–90% – Glyceria maxima, Calamagrostis neglecta, Carex riparia, C. acutiformis; зі сталістю 50–70% – Carex elata. Carex appropinquata, Thelypteris palustris, Typha angustifolia; зі сталістю 25–40% – Calystegia sepi-um і Deschampsia caespitosa. Участь співдомінантів у рослинному покриві становить 20–55%, частіше 25–40%. Неподільно домінує Phragmites australis, проективне покриття якого становить 50–90%, а інколи й більше.
В Степу та Лісостепу чисті очеретяні угруповання займають досить великі площі на мілководдях з малорозвинутим шаром торфу або без нього. За умов достатнього зволоження, тепла повітря, грунту та води, що складаються на таких болотах, очерет дуже активно росте і протягом вегетації може утворювати кореневища до 5–15 м завдовжки, продукуючи велику кількість бруньок відновлення і пагонів. У результаті формуються монодомінантні угруповання, в яких травостій на 80–90% утворений Phragmites austraiis. Як уже відзначалося, висока щільність стеблостояння, затіненість, обводненість та інші властивості очеретяних угруповань зумовлюють бідність і одноманітність рослинного покриву. Високу ценотичну сталість виявляють лише деякі види, котрі тут разом з очеретом утворюють флористичне ядро очеретяних угруповань. Це, зокрема, Typha latifolia, T. angustifolia, Schenoplectus lacustris, S. tabernae-montanii, Carex vesicaria, C. acuta, C. elata, C, caespitosa, C. acutiformis, C. riparia, Oenanthe aquatica, Myosoton aquaticum, Scuteliaria galericulata, Rumex hydrolapathum, Ranunculus lingua, Sium latifoliurn, Leersia oryzoides. Всі ці види не досягають висоти едифікатора і знаходяться в різних умовах матеріально-енергетичного забезпечення, а звідси мають різну енергію росту, неоднорідну ярусну почленованість, видову насиченість, а отже, й різну продуктивність.
3.3.2 Угруповання Typha angustifoliaУгруповання Typha angustifolia поширені на залитих водою болотах стариць, розмитих знижень других терас, вигорілих та вироблених кар'єрів. Поселяється рогіз як на мінеральних ґрунтах і вологих або затоплених пісках прируслової зони заплав, так і на торфових покладах різної потужності.
Місцезростання Typha angustifolia заливаються талими і повеневими водами, рівень стояння яких протягом вегетації коливається від 0 до 1,75 м. У заплаві р. Дніпра, на околицях Кончі-Озерної, біля дамби, де вода постійно підпружується. рогіз росте при товщі води до 2,0 м. Частіше рогіз займає ділянки з товщею води до 20 см, а в літній період зі стоянням води навіть нижче поверхні ґрунту.
За даними літератури, Typha angustifolia досить стійкий до кислотності ґрунтового розчину. При цьому підвищення його стійкості до засолення ґрунту спостерігається при просуванні з півночі на південь. Ми неодноразово відзначали, що на Поліссі ценози Typha angustifolia ростуть на ґрунтах з кислою реакцією ґрунтового розчину, а в лісостепових районах – на засолених, насамперед з карбонатним засоленням та лужною реакцією. За оптимальних умов Typha angustifolia досягає 2,0–2,5 м заввишки, має значну продуктивність. За даними Д.В. Дубини зі співавт. (1993), на водосховищах у Поліссі його продуктивність становить 2,6–3,4 кг/м2, у лісостепових районах – 2,1–5,2, а в степовій зоні (в пониззі Дніпра) – 2,8–5,8 кг/м2.
Typha angustifolia – це типовий водний геофіт, добре закріплений у ґрунті, з повзучим кореневищем. Він утворює корені двох типів: товсті, занурені в мул, які вбирають з нього поживні речовини, і тонкі, спрямовані вгору, розташовані здебільшого у водному середовищі, котрі вбирають з води поживні речовини, воду і кисень, а також допомагають швидкому розселенню рослини. Екологія рослини дуже неоднорідна: на обводнених ділянках з його участю виникають високопродуктивні, ценотично малоструктуровані та флористично бідні ценози, що пов'язано з їх одноманітним екологічним та матеріально-енергетичним середовищем. Навпаки, в умовах помірного зволоження структура чітко виявляється у почленуванні травостою на 2–3 яруси, флористичний склад їх збагачується за рахунок інвазії лучно-болотних видів зі злаків та різнотрав'я. Якість травостою поліпшується, а продуктивність ценозів знижується. Слід відзначити, що рогіз не витримує тривалого осушення, тому на меліорованих болотах та при зниженні рівня ґрунтових вод він поступово втрачає ценотичні позиції в травостої і практично на 3–4-й рік повністю зникає зі складу фітоценозу.
Вузьколисторогозові угруповання монодомінантні. Вони складаються майже повністю з Typha angustifolia із загальним покриттям 70–90%, мають спрощену структуру з добре виявленим одним ярусом, утвореним основним едифікатором. Разом з тим рогозові болотні угруповання бідні за флористичним складом і одноманітні за структурою і фізіономічністю трав'яно-мохового покриву.
У чистих заростях рогозу внаслідок щільності стеблостояння, затоплення та засоленості ґрунту росте обмежена кількість видів, чим пояснюється низька флористична насиченість. У їх складі чимало водно-болотних видів, зокрема, Sagittaria sagittifolia, Sium latifolium, Typha latifo-lia, Salvinia natans, Ranunculus lingua, Oenanthe aquatica, Nymphaea alba, Nuphar luteum, Phragmites australis.
Рогозові угруповання досить різноманітні ценотично. Вони представлені кількома типовими групами асоціацій – рогозово-очеретяною, рогозово-куговою, рогозово-лепешняковою, рогозово-осоковою. Однією з найпоширеніших є рогозова та її монодомінантна асоціація Typhetum (angustifoliae). Едифікатор – Typha angustifoiia з проективним покриттям до 95%. Завдяки високому та щільному стеблестоянню в угрупованні росте дуже мало видів; із сталістю 80–100% зустрічаються Phragmites australis, Typha latifolia, Scirpus lacustris, Carex acutiformis; 60–80% – Carex rostrata, Glyceria fluitans, Calamagrostis neglecta, Thelypteris palustris, Galium palus-tre, Menyanthes trifoliata. Зі сталістю менше 60% ростуть звичайно Carex elata та С. appropinquata, Iris pseudacorus, Lysimachia vulgaris, Nymphaea alba, Sparganium, Ranunculus flammula та інші менш поширені види.
3.3.3 Рогозово-очеретяні угрупованняРогозово-очеретяні угруповання найчастіше трапляються по водоймах і найбільш знижених долинних і староруслових болотах, уздовж заторфованих русел річок, у верхів'ях заторфованих річок з рівнем стояння води до 1,0 м і більше та щільному стеблостоянні високотравних видів. Як результат обводненості місцезростання, в травостої слабо виявлена його вертикальна почленованість та флористична насиченість судинних видів. Характерними представниками цих угруповань є: Potamogeton crispus, P. lucens, P. natans, Polygonum amphibium, Nuphar luteum, Nymphaea alba, Hydrocharis morsus-ranae, Stratiotes aloides, Schoenoplectus lacustris,
Lemna trisulca, Oenanthe aquatica тощо, які займають відкриті водойми, поступово заторфовуючи їх ложе внаслідок утворення торфових покладів. Асоціативність цих співдомінантів забезпечує високу щільність стеблостояння, однак тут відзначаються висока продуктивність травостою, низька флористична насиченість ценозів, бідний видовий склад асоціацій, слабка структурна почленованість угруповань.
Звичайно очеретово-вузьколисторогозові угруповання на досліджених нами болотах є найпоширенішими. На рогіз у травостої припадає 45–75% загального проективного покриття, на Phragmites australis – 25–55%. Ці два види утворюють бідомінантні угруповання з одно – або двох'ярусною почленованістю. Крім того, тут часто трапляються з високою сталістю й інші види, які були відмічені для попередньої асоціації.
Вузьколисторогозово-озернокугові угруповання трапляються на прибережних дуже залитих водою ділянках боліт або на заростаючих обмілинах водосховищ. їх особливістю є домінування двох видів – едифікатора Typha angustifolia з проективним покриттям 60–80% та співедифікатора Schoenoplectus lacustris з проективним покриттям 20–40%. Видова насиченість фітоценозів становить 10–20 видів водно-болотної екології. Решта видів така ж, як у першій асоціації, але з деякою процентною відмінністю.
Рогозово-лепешнякові угруповання властиві менш обводненим болотним екосистемам, де висота стояння води становить 20–40 см, рідше 60 см. За таких умов у травостої зростає флористична насиченість і структурна почленованість, надземна і підземна ярусність. Ценотичні властивості пов'язані зі ступенем зволоження: чим вищий рівень стояння води, тим більша участь рогозу (35–70%) і, навпаки, в умовах спаду води зростає щільність заселення екотопу Glyceria maxima (до 65%). Постійними супутниками цих угруповань є: Glyceria fluitans, Naumburgia thytsinora, Sagittaria sagittifolia, Lythrum salicaria, Hottonia palustris, Ranunculus lingua, Euphorbia palustris, Stachys palustris тощо.
Рогозово-осокові угруповання досить поширена в умовах перемінного зволоження або періодичного затоплення у весняний період. За умов застійного зволоження по копанках, виїмках торфу, по пересихаючих канавах зростають угруповання рогозу (30–50%) зі співдомінуванням осок (20–40%), зокрема Carex elata, C. riparia, C. appropinquata, C. rostrata, C. acutiformis. Ці осоки створюють другий ярус висотою до 80 см та проективним покриттям 40–60%. За домінуванням серед осок окремих видів виникають рогозово-високоосокова, рогозово-зближеноосокова, рогозово-гостровидноосокова, рогозово-побережноосокова асоціації з виявленням флористичних та ценотичних особливостей.
Вузьколисторогозово-побережноосокові угруповання властиві для прируслових і притерасних знижень з періодичною проточністю води, що відіграє важливе значення для їх оптимізації. Відмінністю цього таксона є чітке почленування травостою на яруси; співдомінантна участь Typha аngustifolis тa Carex riparia; антропогенний прес, що зумовлює появу стадій і фаз сукцесійних змін рослинності болотних екостистем.
На болотах з перемінним зволоженням часто зустрічаються рогозово-мітлицеві угруповання. Особливістю цих боліт є пересихання екотопу в літній період, розрідженіть травостою рогозу до 25–40%, суцільний розвиток Agrostis stolonifera та поява видів лучно-болотної екології.
Місцями на болотах-блюдцях до Typha angustifolia та Agrostis stolonifera додаються гіпнові мохи, які утворюють рогозово-осоково-гіпнові та рогозово-мітлицево-гіпнові угруповання. Постійні компоненти цих угруповань – Alisma plantago-aquatica, Polygonum amphibia, Ranunculus repens, Oenanthae aquatica. Galium palustre, Glyceria fluitans, Acorus calamus, Carex vesicaria, C. acuta, Equisetum palustre, Juncus gerardii.
Як і решта високотравних видів, Giyceria maxima на торфових болотах і заболочених землях формує угруповання по окраїнах водойм, а також по зниженнях у центральній та притерасній заплавах малих і середніх річок. Його особливістю є пристосування до тривалого затоплення талими та проточними водами, де товща водного шару досягає 1,0–1,5 м. В умовах високого обводнення і достатньої кількості тепла, а також зольного живлення, Glyceria maxima знаходить оптимальні умови для свого розвитку і досягає максимума висоти і продуктивності травостою.
Серед лепешняків виділяємо два типи угруповань: перші розвиваються на мінеральних, торф'янистих ґрунтах і заболочених землях, а другі сформувалися на торфовищах різної потужності торфу. Перші з них являють лучно-болотні або лучні, а другі – болотні фітоценози, що мають відмінний ботанічний склад та продуктивність угруповань.
Характерною ознакою великолепешнякових угруповань є домінування в травостої основного едифікатора – Glyceria maxima. Його участь залежить від екології місцезростання: на обводнених ділянках він створює суцільні зарості висотою 1,0–2,0 м з проективним покриттям 70–90%. Навпаки, із зменшенням обводнення і зниженням рівня ґрунтових вод до 5–10 см або до 0 в рослинному покриві зростає участь середньорослих, менше низькорослих видів осок, злаків і болотного різнотрав'я.
У результаті змінюється структура, флористична насиченість і продуктивність рослинних угруповань. Так, у структурі рослинних угруповань виникають 2–3-ярусні угруповання. У флористичному складі збільшується кількість злаків і болотного різнотрав'я, у зв'язку з чим поліпшується кормова цінність великолепешнякових угруповань.
Зарості Acorus calamus досить часто зустрічаються навколо ставків, по долинах малих річок, по зниженнях центральної та притерасної заплав. Вони створюють монодомінантні угруповання висотою до 1,0–1,2 м. Зростаючи густими заростями, вони обмежують поширення інших видів рослин. Тому їх угруповання флористично бідні і одноманітні, досить прості за структурою.
Зарості Acorus calamus одно-двоярусні: перший ярус висотою до 1,2 м утворює Acorus calamus з покриттям 50–80%, Glyceria maxima (5–15%), Phragmites australis (1–10%), Calamagrostis negiecta (1–5%), Scnoenopiectus lacustris (1%), Typha angustifolia (1%). Місцями за сприятливих умов, особливо надмірного зволоження або затоплення, Glyceria maxima сильно розростається і досягає ролі співдомінанта, утворюючи кондомінантні лепешняково-лепехові угруповання.
Другий ярус більш-менш чітко виявлений в угрупованнях, які сформувалися на помітно зволожених або періодично підсихаючих ділянках. За цих умов рясно розвиваються Carex vesicaria, C. acuta, рідше Poa palustris та Agrostis stolonifera, які досягають 20–40% проективного покриття і відіграють роль співедифікаторів рослинних угруповань, утворюючи осоково-лепехові, мітлицево-пепехові і тонконогово-лепехові ценози. їх постійними супутниками тут є: Galium palustre, Lycopus europaeus, Menyanthes trifoliata, Alisma plantago-aquatica, Veronica beccabunga, Eleocharis palustris, Myosotis palustris, Coronaria flos-cuculi, Ranunculus flammula, Inula britannica, Gratiola officinalis, Thelypteris palustris та інші менш поширені види, але їх загальне проективне покриття не перевищує 20%.
Угруповання Equisetum fluviatile мало поширені, часто трапляються тільки на долинних і староруслових, рідше улоговинних болотах. Вони займають зниження, де застоюється вода, інколи вона стоїть шаром 20–40 см майже ціле літо. Звичайно вона опускається до рівня поверхні болота. Зарості Equisetum fluviatile ростуть на торф'янисто- і торф'яно-глейових, а частіше на торфових грунтах потужністю до 3,0 м.
Ценотично це майже виключно однодомінантні угруповання з низькою флористичною насиченістю. Equisetum fluviatile за оптимальних умов водного та зольного режиму утворює одноярусні зарості висотою до 1,2 м заввишки та зімкнуїіиію іравистою 60-» 0%. У невеликій кількості до нього домішуються Phragmites australis, Equisetum palustre, Sparganium erectum, Schoenoplectus lacustris.
Третій ярус формують гіпнові мохи. Основними домінантами виступають: Drepanocladus aduncus, D. vernicosus, Calliergon giganteum, Calliergonella cuspidata, Mnium medium, Cratoneurum filicinum, Tomenthypu nitens, але найчастіше домінантами виступають перші чотири види з проективним покриттям 40–70%.
Утворювані Sparganium erectum угруповання мають локальне поширення на лісостепових, а ще рідше на поліських болотах. Вони приурочені до найбільших понижень у заплавах річок, по долинних і староруслових болотах, рідко трапляються на улоговинних болотах других борових терас. їх особливістю є обводненість поверхні боліт. Вода стоїть шаром 20–40, інколи до 60 см. У зв'язку з цим зарості їжачої голівки часто заселяють дуже обводнені окраїни боліт, окраїни водойм, канави меліоративної мережі, копанки тощо, де вода стоїть шаром до 20 см навіть у найжаркіший період. За оптимальних умов зволоження і багатого мінерального живлення Sparganium erectum створює суцільні зарості висотою до 1,5 м з проективним покриттям до 90%. Тільки невелика участь інших видів доповнює цей ярус, частіше це Phragmites austraiis, Schoenoplectus lacustris та Typha angustifolia. Мало виявлений другий ярус. Часто він утворюється в результаті підсихання боліт та на болотах, які досягли високої евтрофності. Ці зарості ускладнюються з утворенням другого, а місцями навіть третього ярусу, особливістю якого є мала флористична насиченість. Другий ярус звичайно створюють: Oenanthe aquatica, Butomus umbellatus, Eleocharis paiustris, Alisma piantago-aquati-ca Glyceria fluitans, Veronica beccabunga, Poiygonum amphibium тощо.
На болотах, які періодично підсихають або тривалий час мають низький рівень стояння вод, трапляються угруповання Sparganium erectum зі співдомінуванням Carex rostrata (20–35%) та Oenanthe aquatica (20–30%). В цих угрупованнях, крім уже згаданих видів, досить рясно ростуть такі, як Mentha aquatica, Carex riparia, Naumburgia thyrsiflora, Menyanthes trifoliata, Comarum palustre, Hottonia palustris, Utricularia vulgaris, Lemna minor, Ranunculus flammula.
Рослинний покрив лісостепових боліт різноманітний і представлений певними еколого-морфологічними комплексами рослинних угруповань, їх специфічність виявляється у поєднанні рослинного покриву з навколишнім абіотичним середовищем. Помітне місце в структурі рослинного покриву боліт займають купинястоосокові болотні комплекси. Основним лімітуючим фактором їх розподілу є ступінь зволоження місцевості.
За умов високого стояння талих або повеневих вод, товща яких у весняно-літній і ранньовесняний періоди становить 20–30 см, а влітку в жаркий період знижується до рівня поверхні ґрунту або на 5–10 см нижче, формуються купини осок. У такий спосіб кореневі системи купинястих видів осок краще забезпечуються киснем, що є істотним фактором розвитку болотних і водно-болотних видів рослин.
На купинах внаслідок шільного корененасичення, щорічного відмирання і відчуження решток рослин – листків, квіток, суцвіть, плодів, насіння, а також осідання дрібноземистих пилових часток створюється рихло- і грубогумусовий субстрат для поселення інших видів квіткових і вищих спорових рослин.
У травостої неподільно домінує Carex elata, котра за оптимальних умов водно-мінерального живлення та проточності води досягає 1,0–1,5 м заввишки і утворює суцільні зарості з мінімальною видовою насиченістю Видовий склад цих угруповань збагачується тільки там, де водойми періодично підсихають, а рослини зазнають антропогенного впливу.
У складі зближеноосокових болотних угруповань виділяють такі угруповання: осокові, осоково-гіпнові і злаково-осокові, рідше злаково-осоково-гіпнові.
Угруповання Carex caespitosa не належать до числа поширених. Дернистоосокові рослинні угруповання не відіграють помітної ролі в структурі болотної рослинності. Як і попередні угруповання, луківники відносять до лучних угідь. На наш погляд, це типові болотні угруповання
Дернистоосокові угруповання часто займають обводнені ділянки боліт, їх окраїни, вони характеризують перехідну ланку між типовими болотами та луками і лісами, проходячи при цьому ряд лучно-болотних і болотно-лісових стадій. Основним едифікатором є Carex cespitosa. проективне покриття якої в таких умовах місцезростання збільшується від 35–50% до 60–75%. Carex cespitosa завжди росте в першому ярусі, рідше в другому Вона утворює купини до ЗО см заввишки і 20–60 см в діаметрі. Поміж купинами навесні та восени стоїть вода (рідше протягом вегетації). Звичайними супутниками осоки дернистої є: Carex acuta, C гірагіа, С. rostrata, C. nigra, Galium palustre, Naumburgia thyrsiflora Scuteilaria galericulata, Agrostis stolonifera, Poa palustris, Lysimachia vulgaris, Epilobium palustre, Lycopus europaeus, Ranunculus lingua, Lychnis flos-cuculi, Myosotis palustris, Hottonia palustris, Cardamine pratense та інші менш поширені види болотного різнотрав'я.
На помірно або дуже зволожених болотах дернистоосокові угруповання маюіь добре розвинутий покрив. Тут утворюються осоково-гіпнові болотні ценози, а на підсушених або періодично пересихаючих болотах – осокові або злаково-осокові фітоценози.
Під впливом антропогенного тиску відбуваються істотні зміни в структурі та флористичному складі побережноосокових угруповань, що позначається на їхній продуктивності. Зокрема, зі зниженням рівня болотних живлячих вод із травостою випадають перш за все гіпергідрофільні види болотного різнотрав'я та мохового покриву, внаслідок чого замість гіпергідрофільних фітоценозів формуються мезофільні та гігромезофільні. Відповідно до цього змінюються умови місцезростання. На зміну осоково-гіпновим ценозам з'являються злаково-осокові-гіпнові та злаково-осокові, котрі являють собою окремі ланки послідовних змін від болотних до лучно-болотних та лучних угруповань.
Угруповання Сагех acuta досить поширені і займають помітне місце в рослинному покриві боліт і заболочених територій. Вони ростуть на торф'яних і торф'яно-глейових ґрунтах з шаром торфу різної потужності, утворюючи торф'яні і торф'янисті відміни. Неоднорідні вони й за ступенем зволоженості, по зниженнях вони залиті водою, а по вирівняних ділянках боліт надмірно зволожені або періодично підсихають. Слід відзначити, що ряд геоботаніків відносять гостроосокові угруповання до лучних угідь. Ми їх відносимо до болотних і розглядаємо як самостійну формацію.
Гостроосокові болотні угруповання, зростаючи в умовах багатих евтрофних грунтів, у тому числі на торф'янисто- і торф'яно-глейових відмінах з достатнім мінеральним живленням і належним водозабезпеченням, досить різноманітні за структурою, видовим насиченням і продуктивністю.
У структурі гостроосокових угруповань виділяється кілька ярусів Перший з них утворює Сагех acuta, участь якої становить 80% загального проективного покриття. Висота травостою 70–100 см. Звичайними її супутниками є: Сагех acutiformis, Calamagrostis neglecta, Glyceria fiuitans. Sium latifolium, Poa palustris, Iris pseudacorus, але їх сумарне покриття не перевищує 20%.
У другому ярусі заввишки 40–70 см найчастіше і в більшій кількості трапляються: Agrostis stolonifera, Equisetum palustre, Poa trivialis, Butomus umbeliatus, Lysimachia vulgaris, Stellaria palustris, Comarum palustre тощо У третьому ярусі (20–40 см) постійними компонентами є: Mentha arvensis, Caltha palustris, Galium palustre, Myosotis palustris, Coronaria floscuculi, Juncus garardii.
Гостроосокові угруповання часто зустрічаються на заболочених землях, займаючи проміжне положення між типовими болотними і лучними угіддями. На осушених болотах вони являють собою один з етапів повторного заболочування раніше осушених низинних боліт, спричиненого підпруженням болотних вод у меліоративній мережі та антропогенним впливом на торфовище та його фізико-хімічні властивості, в результаті чого ущільнюється торфовище і посилюється гуміфікація торфу.
Угруповання Carex acutiformis займають обводнені або дуже зволожені ділянки (місцями з проточною водою) на лісостепових болотах За достатнього зволоження, багатого органічно-зольного живлення та хоча б періодичної проточності Carex acutiformis досягає 1 м заввишки. Нерідко вона утворює чисті зарості.
Угруповання Carex acutiformis є типовим прикладом одноярусних монодомінантних угруповань. На болотах, не порушених господарською діяльністю людини, часто формуються монодомінантні фітоценози, в яких Carex acutiformis займає провідне положення (85% загального проективного покриття). В утворюваних угрупованнях немає чіткого вертикального почленування травостою, через незначну участь у ньому інших видів та низьке проективне покриття останніх не виявляється повна відокремленість тих чи інших груп видів, що, мабуть, можна пояснити однорідністю вимог супутників Carex acutiformis до умов місцезростання. Таким чином, з одного боку, вирівняність екологічного фактора, а з іншого, – фітоценотична однорідність вимог рослин до абіотичного середовища й спричинили одноярусність гостровидноосокових угруповань.
Щільність стеблостояння, добра облисненість рослин, високе проективне покриття, а також обводненість місцевості – чинники, під впливом яких у травостої цих угруповань росте обмежена кількість видів (10–15 видів квіткових і вищих спорових рослин).
Carex acutiformis вимоглива до зволоження, за своєю екологією вона належить до гідрофітів і часто зустрічається тільки на непорушених болотних масивах. На меліорованих болотах о. гостровидна часто і в значній кількості випадає зі складу рослинного травостою. При підсиханні і помірному зволоженні утворює злаково-осокові угруповання мезогігрофільної екології.
Угруповання Carex rostrata досить поширені на дуже зволожених або обводнених долинних і заплавних притерасних болотах, але великих площ тут не займають. Carex rostrata часто зустрічається в комплексі з Carex acutiformis, C. riparia, C. acuta та іншими болотними видами, з якими і утворює специфічні рослинні угруповання. Такі угруповання займають проміжне положення між дуже обводненими та помірно або мало зволоженими осоковими відмінами.
Травостій здутоосокових угруповань досить розріджений. При зниженні обводнення і застійності води у наземному покриві зростає участь гіпнових мохів, а на періодично підсихаючих – злаків та болотного різнотрав'я Відповідно до цього в складі даної формації виділяють осоково-гіпнові, осокові, осоково-різнотравні угруповання.
Осокові та осоково-гіпнові угруповання характеризую! ьсн неподільним домінуванням у травостої С. rostrata, на яку припадає 45–90% загального проективного покриття. В осоково-гіпнових угрупованнях участь С. rostrata знижується, тут її проективне покриття становить 50–60% від загального. Висока щільність стеблостояння С. rostrata визначає продуктивність травостою болотних угідь, а її видова насиченість – їх кормову цінність та ценотичні властивості.
Здутоосокові угруповання за флористичним складом досить різноманітні. Звичайними супутниками домінанта є: Carex acutiformis, С. appropinquata, C. pseudocyperus, C. chordorhiza, Phragmites australis, Glyceria fluitans, Agrostis stolonifera, Poa palustris. Із різнотрав'я найчастіше трапляються такі види, як Lysimachia vulgaris, Galium palustre, Lythrum salicaria, Gratiolla officinalis, Ranunculus lingua, Oenanthe aquatics та інші види з проективним покриттям менше 1%.
Злаково-осокові і злаково-осоково-гіпнові угруповання з домінуванням Carex rostrata ростуть в умовах меншого обводнення та зволоження, що сприяло зростанню в травостої частки лучно-болотних злаків, на які тут припадає 25–40% загального покриття. Окремі види болотних злаків досягають значної участі та ценотичної ролі співдомінанта. Типовими для здутоосокових угруповань є: Glyceria fluitans, Poa palustris, Agrostis stolonifera, Calamagrostis neglecta. За їх поєднанням з домінантом виникають здутоосоково-повзучомітлицеві, здутоосоково-болотнотонконогові, здутоосоково-плаваючелепешнякові, здутоосоково-непомітнокуничникові та інші болотні угруповання, які генетично споріднені з меліоративними угрупованнями.
У складі формації Carex diandra виділяються монодомінантні та полідомінантні угруповання. У їх структурі виявляються два-три яруси.
Перший ярус утворюють: Carex diandra (домінант, покриття 50–70%), С. elata, Calamagrostis neglecta, Glyceria fluitans, Phragmites australis, Phalaroides arundinacea та деякі інші менш поширені види.
У другому ярусі заввишки 40–60 см зростає більше видів, з яких найтиповішми для даних угруповань є: Carex rostrata, C. vesicaria, C. limosa, Iris pseudacorus, Lysimachia vuigaris, Galium palustre, Agrostis stolonifera, Poa palustris, Equisetum fluviatile, Ranunculus lingua, Comarum palustre, Callapalustris, Oenanthe aquatica, Peucedanum palustre, Menyanthes trifoliate, Naurnburgia thyrsificra та інші види з проективним покриттям менше 1%.
Угруповання Carex diandra, на думку Д.К. Зерова (1938), характерніші для боліт, підстилаючими породами яких є леси або відслонення крейдяних порід. За даними Л.Г Раменського зі співавт. (1956), осока двотичинкова масово зростає на досить багатих і багатих болотних грунтах з тенденцією до розвитку в умовах засоленості.
Особливістю цих угруповань є слабо розвинутий наземний травостій і чітко виявлений моховий покрив. Типовими його компонентами тут є: Drepanocladus sendtneri, D. vernicosus, D. aduncus, Calliergon giganteum, Calliergonella cuspidata, Mnium intermedium, Climacium dendroides, Bryum ventricosum, Campylium polygamum та інші менш поширені види.
Угруповання Carex vesicana поширені на лісостепових болотах, де займають невеликі ділянки, і приурочені до місцезростань, які періодично затоплюються водою. Звичайними вони є по заболочених тальвегах балок, навколо ставків, уздовж річок. Вони не займають великих площ, але утворюють специфічну проміжну ланку між типовими торфовими болотами і заболоченими землями або болотистими луками, що ростуть на мінеральних грунтах чи на торф'яно- і торф'янисто-глейових відмінах.
Carex vesicaria утворює монодомінантні одно-двох'ярусні болотні угруповання Добре зволожені ділянки або періодично затоплювані водою покриваються одноярусними угрупованнями. Основним едифікатором у них є Carex vesicaria, котра за оптимальних умов водно-мінерального живлення досягає висоти 80 см і утворює густий одноманітний покрив з проективним покриттям 75–90%. Цей ярус збагачують лише Agrostis stolonifera, Carex vuipina, Glyceria fiuitans, котрі ростуть тут у невеликій кількості.
На менш зволожених ділянках, а також на осушених і вторинно заболочуваних локалітетах виникають пухирчастоосокові угруповання складнішої структурної організації. Зміни, яких вони при цьому зазнають, спричинені переважно господарською діяльністю людини, зокрема систематичним сінокосінням, випасанням на болотних угіддях великої рогатої худоби, через що руйнується дернина тощо.
Внаслідок розрідженості травостою, руйнування дернини, зникнення мохового покриву тощо під покрив Carex vesicaria проникають Інші менш вологолюбні види, особливо зі злаків та лучно-болотного різнотрав'я. Зі збільшенням участі цих видів формуються різноманітні пухирчасто-осокові угруповання. В результаті чисто пухирчастоосокові угруповання змінюються осоково-злаковими, осоково-злаково-різнотравними і осоково-різнотравними, які становлять гідрофільний екологічний ряд асоціацій, що відображають зміни флористичного складу та ценотичних взаємозв'язків ценоелементів із тенденцією до мезофітизації та оліготрофізації болотних екосистем.
3.4 Трав'янисто-мохові болотаНа болотах помітне місце в рослинному покриві займають трав'яно-мохові ценози. Характерною їх ознакою є відсутність будь-якого вираженого деревостану або чагарникового ярусу. Трапляються окремі дерева або біогрупи, окремі кущі чи їх невеликі плями. їх особливістю є безлісність, відкритість і непорушність фітоценозів.
Порівняно з трав'яними, трав'яно-мохові угруповання відрізняються рядом еколого-ценотичних і господарсько цінних ознак:
1. На трав'янисто-мохових болотах поширені тільки трав'янисто-гіп-нові болотні угруповання. Трав'янисто-сфагнові угруповання трапляються невеликими ділянками на староруслових болотах (долина р. Супій). Із мохів тут відмічено ряд сфагнів евтрофної екології – Sphagnum subsecundum, S. palustre, S. contortum, S. teres, але їх проективне покриття (15–50%) неоднорідне.
2. У травостої переважають ті ж види злаків і осок, що і в трав'янистих рослинних угрупованнях. У цьому виявляється генетична спорідненість двох класів формацій рослинних угруповань. Трав'янисто-мохові угруповання мають і певні ценотичні відмінності. Трав'янисті види домінують у структурі травостою, а їх участь у проективному покритті фітоценозів зменшується. Знижуються фітоценотична роль цих видів від едифікатора до субедифікатора, а в деяких ценозах до рівня асектатора.
3. Основними домінантами трав'янисто-гіпнових боліт є такі купинні види осок: Carex elata, C. appropinquata, а в північних районах на межі з поліськими – С. caespitosa. Перші два види властиві надмірно зволоженим або обводненим ділянкам, а останній – періодично надмірно або постійно зволоженим екотопам. Із кореневищних видів на дуже обводнених місцезростаннях ростуть Carex diandra, C. rostrata, C. lasiorarpa, а на періодично підсихаючих – С. nigra, С. сіпегеа, С. рапісеа та ін. Рідше в складі цих угруповань субдомінантами виступають види болотного різнотрав'я.
4. Флористичний склад трав'янисто-гіпнових болотних угруповань бідніший і одноманітніший, ніж у трав'янистих угрупованнях. Особливо зменшується кількість трав'янистих видів, насамперед видів різнотравного високотрав'я, таких як Valeriana officinalis, Peucedanum palustre, Filipendula denudata, Oenanthe aquatica та інші менш поширені види. Тому в окремих ценозах налічується не більше 15–20 видів вищих спорових і квіткових рослин.
5. Види трав'янисто-мохових, а фактично трав'янисто-гіпнових і навіть осоково-гіпнових угруповань гірше ростуть і розвиваються в разі погіршення азотного та мінерального живлення. їх висота знижується до 60–80 см, а проективне покриття – до 40–60%, у зв'язку з чим помітно послаблюється фітоценотична роль основних фітоценотипів квіткових і вищих спорових рослин.
6. У складі трав'янисто-гіпнових угруповань все більшу едифікаторну роль відіграють не трав'янисті види, з ппнові мохи, зокрема Drеpanocladus sendtneri, D. vernicocus, Calliergonella cuspidata, Calliergon trifarium, C. giganteum, Bryum ventricosum, Brachythecium mildeanum, Mnium medium, M. affine, Tomenthypnum nitens, Scorpidium scorpioides. На періодично підсихаючих болотах зростає участь у моховому покриві таких видів гіпнів, як Aulacomnium palustre, Climacium dendroides та деяких інших. Значна участь гіпнових мохів у фітоценозах, їх ценотична активність і структурно-екологічні властивості – ось фактори, під впливом яких послаблюється едифікаторна роль трав'янистих видів. Це у свою чергу призводить до спрощення та збіднення флористичного складу рослинних угруповань.
7. У результаті осушення та впровадження інтенсивних технологій освоєння й експлуатації торф'яного фонду регіону в трав'яному покриві порушених болотних масивів сталися істотні зміни флористичного складу та ценотичної структурованості. В структурі рослинного покриву багатьох боліт замість осоково-гіпнових сформувалися злаково-осоково-гіпнові, рідше злаково-гіпнові угруповання, що помітно позначилося на продуктивності та якості травостою кормових угідь меліорованих боліт і перезволожених земель.
Висновки
На теренах України переважають низинні болота з евтрофною рослинністю. Вони надзвичайно різноманітні за своєю природою, що пояснюється умовами водно-мінерального живлення.
Серед евтрофних боліт виділяємо три групи формацій. З них – найбільш поширеними і досить різноманітними трав'яні болота, в меншій мірі – лісові, фрагментарно – чагарникові. Основу трав'яного покриву евтрофних боліт складають: лепешняк великий ((llyceria maxima), очерет звичайний (Phragmites australis), різні види осок (Care.x acutifonths, С. omskiana, C. vesicaria, C, acutd) та інші види болотного різнотрав'я (хвощ річковий (Equisetum fluviatile), плакун верболистий (Lyihrum sahcaria), вербо чіл ця звичайне (Lysimachia vulgaris))
Лісові евтрофні болота на Чернігівщині представлені двома формаціями – вільхи клейкої (Alneta (glutinosae) paludosa) і берези болотної (Betulela (pubescentis) pahulosa), з яких більш поширеною (перша. Класичне місце зростання лісових бони і домінуванням вільхи клейкої (Alnus giutinosci) – притерасна частина заплави, пере (воложена, багата мінеральними речовинами. Характерною особливістю вільшинкових боліт мозаїчність рельєфу – наявність п'ядесталів та міжстовбурових знижень У зниженнях переважають осоки (о.побережна (Carex riparia), о.пухирчаста (С. vesicaria), о госіровидна (С. acutiformis)) та види гідрофільного різнотрав'я (образки болотні (Colla palustris), бобівник трилистий (Menyanlhes trifoliata), кизляк болотний (Naumburgia thyrsijlora) На пристовбурових підвищеннях трапляються різні види гідрофільних папоротей (тсліптерисі болотний (Thelypteris palustris), nut шик шартрський (Drypteris carthusiana), безщитник жіночий (Athyrium filix-femina) щитник гребінчастий (Drypteris cristata)), види лучно-і болотного різнотрав'я.
Найбільш представленою серед евтрофних боліт є група асоціацій трав'яних боліт, їх І ценози звичайно розвиваються в умовах надмірного зволоження і вкриванні, більшу частину площі долинних, заплавних та частково притерасних боліт. Рослинний покрив трав'яних евтрофних боліт представлений осоковими, злаковими та лепешняковими ценозами формацій осоки омської (Caned omskianae), осоки гострої (С. acutae), осоки пухірчастої (С. vesicariae), лепешняку великого (Glycerieta maximae), очерету звичайного (Vhragmiteta austrsalis).
Сучасні трав'янисто-мохові болота та їх рослинний покрив під впливом господарської діяльності людини зазнали істотних змін і потребують, з одного боку, реструктуризації меліоративного фонду, а з іншого, – удосконалення методів поліпшення природних угідь і торфовищ з метою раціонального використання рослинницької і торфової сировини.
Список використаних джерел
1) Александрова В.Д Изучение смен растительного покрова // Полевая геоботаника. – М.-Л.: Наука. – 1964. – Т. 3. – С. 300–447.
2) Александрова В.Д. Классификация растительности. Обзор принципов классификации и классификационных систем в разных геоботанических школах. – П.: Наука, 1969. – 275 с.
3) Білявський Г.О. та інші. Основи екологічних знань. – К.: Либідь, 2004. – 408 с.
4) Бойчук Ю.Д., Солошенко Е.М., Бугай О.В. Екологія і охорона навколишнього середовища. – Суми, 2002. – 284 с.
5) Брадіс Є.М. Рослинний покрив боліт УРСР /7 Рослинність УРСР Болота. – К.: Наук, думка, 1969. – С 34–135.
6) Брадіс Є.М., Бачуріна Г.Ф. Болота УРСР. – К: Наук, думка, 1969. – 242 с
7) Гейнріх Д., Гергт М. Екологія. – К.: Знання-Прес, 2001. – 287 с.
8) Григора И.М. Происхождение и динамика лесных болот Украинского Полесья: Автореф. дис… д-ра биол. наук. – Киев, 1988. – 44 с.
9) Григора І.М., Соломаха В.А. Лісові болота Українського Полісся (походження, динаміка, класифікація рослинності). – К.: Фітосоціоцентр, 2005. – 415 с.
10) Григора І.М., Соломаха В.А. Рослинність України (еколого-ценотичний, флористичний та географічний нарис). – К.: Фітосоціоцентр, 2005. – 452 с.
11) Загальна гідрологія. /За ред. С.М. Лисогора. – К.: Фітосоціоцентр, 2000. – 264 с.
12) Збереження і невиснажливе використання біорізноманіття України: стан та перспективи. / За ред. Ю.Р. Шелег-Сосонко. – К.: Хімужест, 2003. – 248 с.
13) Зеленая книга Украинской ССР. / Под общ. ред. Ю.Р. Шелег-Сосонко. – К.: Наук. думка, 1987.
14) Карпенко Ю.О. та інші. Дивосвіт природи Чернігівщини: Кн. 2. Луків сіверських різнотрав`я. Світ імлистих боліт і водойм: Навч. посібн. – Чернігів, 2001. – 170 с.
15) Карпенко Ю.О. та інші. Дивосвіт природи Чернігівщини: Кн. 3. Раритети мальовничої Чернігівщини. Живий світ поряд з людиною: Навч. посібн. – Чернігів, 2001. – 163 с.
16) Кокин К.А. Экология высших водных растейний. – М.: Изд-во МГУ, 1982.
17) Кузьмичев А.И. Гидрофильная флора юго-запада Русской равнины и ее генезис. – СПб.: Гидрометеоиздат, 1992. – 215 с.
18) Левина Ф.Я. Болота Черноговского Полесья. // Бот. журнал СССР, ХХІІ, 1937. – №1.
19) Макрофиты – индикаторы изменений природной среды./ Дублена Д.В., Гейне С., Гроудова З. И др. – К.: Наукова думка, 1993. – 435 с.
20) Мулярчук С.О. Рослинність Чернігівщини. – К.: Вища школа, 1970. – 132 с.
21) Мусієнко М.М., Ольгович О.П. Методи дослідження вищих водних рослин. Навч.посібник до лабораторних занять з фізіології водних рослин. – К.: Фітосоціоцентр, 2005.
22) Мусієнко М.М., Серебряков В.В., Брайон О.В. Екологія. Охорона природи: Словник-довідник. – К.: Т-во Знання, КОО, 2002. – 550 с.
23) Определиетль высших растений Украины. / Доброчаева Д.Н., Котов М.И., Прокудин Ю.Н. и др. – К.: Наукова думка, 1987. – 548 с.
24) Парчук Г.В. Проблема дифтрофирования водоемов. // Гидробиологический журнал. – 1994. – №3. – С. 30.
25) Романенко В.Д. Основи гідроекології: Підручник. – К.: Обереги, 2001. – 728 с.
26) Рослинність УРСР. Болота. – К.: Наук думка: 1969. – 243 с.
27) Рычин Ю.В. Флора гигрофитов. – М.: Сов. наука, 1948.
28) Семенихіна К.А. Водна рослинність р. Десни та водойми її заплави в межах УРСР // Укр. бот. журнал. – 1982. – №2. – С. 39.
29) Соломаха В.А. Синтаксономія рослинності України // Укр. фітоцен. зб.-К., 1996.-Сер. А, вип. 4 (5). – 120 с.
30) Управління водно-болотними угіддями міжнародного значення. / За ред. В.А. Костушин. – К.: Національний екологічний центр України, 2005. – 194 с.
31) Федоров А.А. Жизнь растений. – М.: Просвещение, 1974.
32) Хржановский В.Г., Исаин В.Н. Практический курс ботаніки. – М: Высшая школа, 1963. – 299 с.
33) Чорна Г.А. Рослини наших водойм (Атлас-довідник). – К.: Фітосоціоцентр, 2001. – 134 с.
34) Шеляг-Сосонко Ю.Р, Осычнюк В.В., Андриенко Т.Л. География растительного покрова Украины. – Киев: Наук, думка, 1982. – 285 с.
Похожие работы
... з цього питання. Так, Міжнародна конференція у Ріо-де-Жанейро (1991 р.) звернулась до світового співтовариства із заявою про необхідність переходу на шлях сталого розвитку, який передбачає збалансований підхід до розв'язання соціально-економічних проблем і збереження довкілля для майбутніх поколінь. Під водоресурсною складовою сталого розвитку розуміють такий стан водних об'єктів та систем їх ...
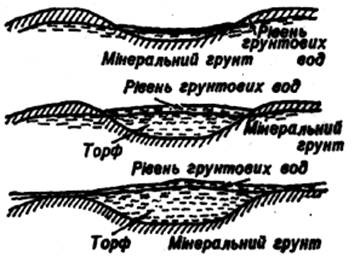
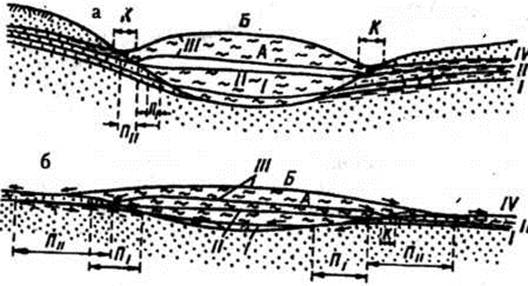
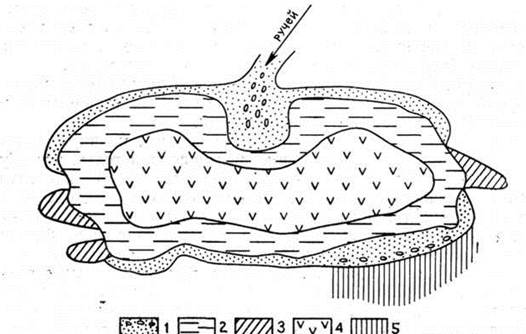
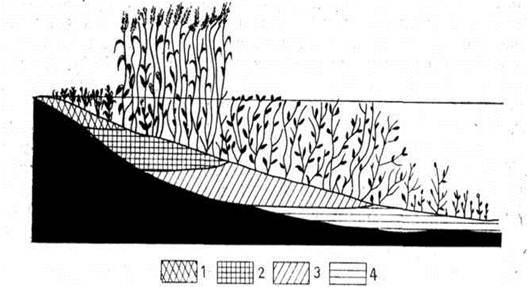
... боліт залежить від кліматичних умов, товщини мерзлого ґрунту і снігового покриву, тому його строки будуть різні в окремих болотних мікроландшафтах. 2. Геологічна діяльність озер і боліт 2.1 Умови формування тирогенних відкладень Вода в озерах має різне походження. Озерні западини можуть бути заповнені річковою водою, водою льодів, що тануть, і снігів, атмосферною або водою підземних ...
... мешкають представники 124 видів тварин, що підлягають особливій охороні згідно з Бернською конвенцією, 15 видів, віднесених до "червоного" списку Міжнародного союзу охорони природи (МСОП), 29 видів вважаються регіонально рідкісними. Фауна заповідника потребує подальшого вивчення. 2.2 Національні природні парки 2.2.1 Швацький національний природний парк Перший національний природний парк на ...
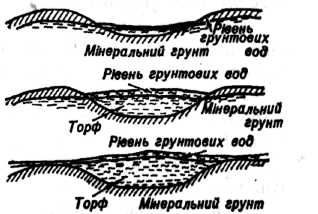

... боліт. Зустрічаються вони в заплавах річок і на заростаючих водоймах. У місцях, де виходять на поверхню підземні води, болота можна зустріти в будь-якій географічній зоні, а також у горах і на гірських схилах. Болото може з'явитись у найрізноманітніших умовах. Часто воно утворюється в ялинковому лісі, де внаслідок вилуговування грунтів з'являється мохова рослинність, яка спричиняє відмирання лісу ...
0 комментариев