Сигналы нервных клеток
Сигналы нервных клеток
Для анализа событий во внешнем мире или внутри нашего тела, для передачи информации от клетки к клетке нейроны используют электрические и химические сигналы. Расстояние передачи сигнала может быть большим: от кончиков пальцев на ногах до спинного мозга. Различные сигналы прекрасно представлены все в той же сетчатке. В то время, когда Рамон и Кахаль рисовал стрелки почти не было информации об этих сигналах, что делает его достижения еще более примечательными
Ступени переработки информации можно последовательно проследить: свет падает на фоторецепторы и генерирует электрические сигналы, которые воздействуют на биполярные клетки. От биполярных клеток сигналы передаются к ганглиозным клеткам и от них к высшим центрам мозга, которые и осуществляют восприятие внешнего мира. В следующих разделах рассматриваются свойства сигналов и пути переработки информации.
Классы электрических сигналовЭлектрические сигналы нервных клеток могут быть разделены на два основных класса. Во-первых, это локальные градуальные потенциалы, которые вызываются такими внешними стимулами, как свет, падающий на фоторецепторы глаза, звуковая волна, деформирующая волосковые клетки уха, или прикосновение, механически смещающее отросток сенсорной клетки в коже. Сходны по характеристикам, но существенно отличаются по происхождению сигналы, генерируемые в синапсах – соединениях между клетками, которые мы обсудим позже. Все эти сигналы градуальны и привязаны к месту возникновения, а их распространение зависит от пассивных характеристик нервных клеток.
Потенциалы действия составляют вторую основную категорию Потенциалы действия вызываются локальными градуальными потенциалами. В отличие от локальных потенциалов, они быстро распространяются на большие расстояния – например, от глаза до высших центров по волокнам ганглиозных клеток, составляющих оптический нерв, или от моторных клеток в спинном мозге к мышцам ноги. Второе отличие потенциалов действия состоит в том, что они фиксированы по амплитуде и длительности, как точки в азбуке Морзе. Крайне существенно понимать, что потенциалы действия, путешествующие по волокнам оптического нерва не являются эпифеноменами, присутствующими лишь в наших представлениях о работе мозга. Они являются единственной формой сигнализации, которая снабжает мозг информацией о внешнем мире.
Передача сигнала от сетчатки может быть представлена следующей упрощенной схемой:
Важным свойством электрических сигналов является то, что они фактически идентичны во всех нервных клетках организма независимо от того, запускают ли они движение, передают ли информацию о цветах, формах или болезненных стимулах, или соединяют различные области мозга. Вторым важным свойством сигналов является то, что они настолько одинаковы у разных животных, что даже умудренный опытом исследователь не способен точно отличить запись потенциала действия от нервного волокна кита, мыши, обезьяны или профессора. В этом смысле потенциалы действия могут считаться стереотипными единицами. Они являются универсальным эталоном для обмена информацией во всех исследованных нервных системах. В мозге не типы сигналов, а огромное количество клеток (от 1010 до 1012 нейронов) и разнообразие связей обеспечивают сложность выполняемых задач.
Эта идея была высказана в 1868 году немецким физиком и биологом Германом фон Гельмгольцем. Беря за основу гипотетические принципы, задолго до обнаружения известных сейчас фактов, он писал: "Нервные волокна часто сравнивают с телеграфными проводами, пересекающими местность, и это сравнение хорошо приспособлено для иллюстрации удивительных и важных особенностей их образа действия. В телеграфной сети везде мы обнаруживаем те же медные или стальные провода, несущие только один вид движения, поток электричества, но вызывающие самые разные результаты на разных станциях в соответствии с дополнительной аппаратурой, с которой провода соединены. На одной станции эффект состоит в звонке колокольчика, на другой сигнал просто передается дальше, на третьей вступает в работу записывающий аппарат….Говоря коротко, каждое из… различных действий, вызываемых электричеством, может быть вызвано и передана проводом в любую необходимую точку. При этом β проводе происходит один и тот же процесс, приводящий к самым разным последствиям….Та разница, которую мы видим при возбуждении различных нервов, заключается только в рознице самих органов, к которым присоединен нерв и которым передается состояние возбуждения".
На самом деле, как будет показано в главе 6, небольшая разница в амплитуде и длительности очевидна в потенциалах действия разных нейронов. Утверждение, что все потенциалы действия одинаковы, равносильно утверждению, что все дубы одинаковы.
Техника записи сигналов от нейронов с помощью электродовДля решения некоторых задач существенно регистрировать активность одного нейрона или даже одного ионного канала, тогда как для других задач необходима суммарная активность многих нейронов. Ниже коротко суммируются основные приемы для записи активности нейронов, используемые для обсуждения в следующих главах.
Впервые запись потенциалов действия от нерва была сделана от периферических нервов экстра клеточным и электродами. Пропускание тока между парой серебряных проводников вызывало потенциал действия, тогда как вторая пара таких же электродов на некотором расстоянии регистрировала ответ. В центральной нервной системе регистрация от нейрона или группы нейронов производится экстраклеточным электродом, который состоит из проводника в изолирующей оболочке или из стеклянного капилляра, заполненного проводящим солевым раствором
С помощью внутриклеточного микроэлектрода мы можем прямо измерять разницу потенциала между наружной и внутренней средой клетки, так же как возбуждение, торможение и возникновение импульсов. Стеклянный микроэлектрод, заполненный солевым раствором и с кончиком менее 0,1 мм в диаметре, вводится в клетку с помощью микроманипулятора Микроэлектроды также используют для пропускания тока через мембрану или внутриклеточной инъекции молекул в цитоплазму.
Часто используется прием измерения мембранного потенциала, известный как пэтч-кламп целой клетки. Стеклянная пипетка со сравнительно большим полированным кончиком придвигается к поверхности клетки, где она прилипает к мембране и образует прочное соединение. После нарушения целостности мембраны внутри пипетки жидкость в пипетке прямо контактирует с внутриклеточной жидкостью.
Неинваэивные методы регистрации нейронной активностиИспользуя метод оптической регистрации, можно проследить передачу информации в некоторых препаратах мозга без использования электродов. Специально созданные красители, которые связываются с клеточной мембраной, изменяют абсорбцию проходящего света или флуоресценцию при изменениях мембранного потенциала клетки, что можно объективно регистрировать. Существуют и такие неинвазивные методы, как позитронно-эмиссионная томография и магнитно-резонансная томография (МРТ), которые позволяют определить, какие области мозга бодрствующего человека активизируются при предъявлении стимулов или при движении. Получаемое с помощью МРТ изображение показывает области, активируемые при предъявлении зрительного стимула.
Ретинограмма отражает суммарную активность сетчатки, электроэнцефалограмма – суммарную активность мозга. Эти методы в основном используются для диагностики нарушений функций мозга.
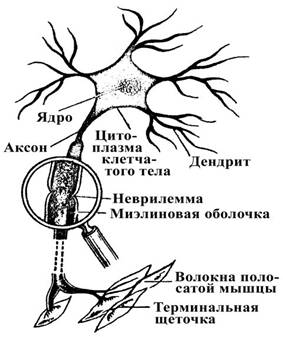
Распределение локальных градуальных потенциалов и пассивные электрические свойства нейронов.В схемах Рамона-и-Кахаля, отображающих клеточное строение мозга, просвечивает идея о том, что освещение сетчатки изменяет активность фоторецепторов и эти изменения отражаются в активности нервных волокон, выходящих из глаза. Для такой передачи информации сигналы должны распространяться не только от клетки к клетке, но и вдоль клетки, от одного ее конца до другого. Как, например, электрический сигнал, генерируемый на контактирующем с фоторецептором конце биполярной клетки, распространяется вдоль нейрона и достигает терминали, которая расположена около ганглиозной клетки?
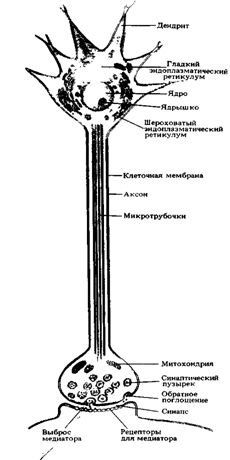
Для того чтобы ответить на этот вопрос, полезно рассмотреть соответствующие структуры, которые передают сигналы. Биполярную клетку можно рассматривать как длинный цилиндр, наполненный водным раствором солей (диссоциированных на положительно и отрицательно заряженные ионы) и белков, отделенный от экстраклеточного раствора мембраной. Внутриклеточный и экстраклеточный растворы осмотически одинаковы, но имеют разный ионный состав. Ионы двигаются по специальным ионным каналам, которые образованы белковыми молекулами, пронизывающими мембрану. Электрические и химические стимулы вызывают открытие или закрытие каналов для ионов кальция, натрия, калия и хлора.
В результате различий в концентрации ионов по обе стороны мембраны и из-за избирательности каналов для определенных ионов образуется потенциал покоя клетки. В покое внутреннее содержимое клетки отрицательно заряжено по отношению к наружной среде. Строение и свойства нейрона определяют способность проведения электрических сигналов. Во-первых, внутриклеточная жидкость, цитоплазма (аксоплазма в отростке клетки, аксоне) примерно в 107 раз хуже проводит электричество, чем металлический проводник. Одной из причин является то, что плотность переносчиков заряда, ионов, в несколько раз меньше, чем электронов в металле; кроме того, подвижность ионов невелика. Во-вторых, протекание тока вдоль аксона на большое расстояние осложняется тем, что мембрана не является идеальным изолятором. Соответственно, величина тока, текущего вдоль волокна, быстро уменьшается из-за утечки через ионные каналы мембраны. Тот факт, что нервные волокна очень малы (обычно не более 20 микрон (мкм) в диаметре у позвоночных), еще больше уменьшает количество проводимого тока. Алан Ходжкин дал интересную иллюстрацию этих свойств распространения электрического сигнала.
Если специалист по электричеству посмотрит на нервную систему, то сразу увидит, что передача сигнала под нервным волокном является огромной проблемой. Диаметр аксона в нерве варьирует от 0,1 до 20 микрон. Внутреннее содержимое содержит ионы и является неплохим проводником электричества. Однако, волокно невелико и его продольное сопротивление очень высоко. Простой расчет показывает, что в волокне диаметром 1 микрон и сопротивлением 100 Ом/см удельное сопротивление составит около 1010 Ом/см. Это означает, что электрическое сопротивление маленького нервного волокна длиной в 1 метр равно сопротивлению 1010 миль 0,2 мм медной проволоки, то есть проволоки длиной в десять раз больше, чем от Земли до планеты Сатурн.
Таким образом, пассивное проведение электрических сигналов затруднено и ограничено расстоянием 1–2 мм. Кроме того, когда такой сигнал короток, его форма может быть сильно искажена и его амплитуда еще уменьшена емкостью клеточной мембраны. Тем не менее, локальные потенциалы очень важны для вызова и проведения распространяющегося сигнала.
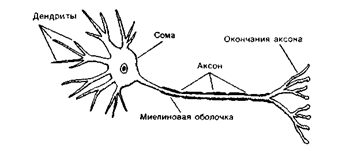
Распространение изменений потенциала в биполярных клетках и фоторецепторахФоторецепторы и биполярные клетки невелики по длине, поэтому локальный градуальный сигнал может эффективно распространяться от одного конца клетки до другого. Электрический сигнал, который отражает попадание света на фоторецептор, генерируется в наружном сегменте палочек или колбочек. Оттуда сигнал пассивно распространяется вдоль клетки до терминали на биполярной клетке. Если бы рецептор или биполярная клетка были длиннее (несколько миллиметров в длину), то локальный потенциал из-за сильного ослабления не достиг бы терминали и не смог влиять на следующую в цепи клетку. Биполярные клетки и фоторецепторы представляют собой исключение из общего правила, которое гласит, что для переноса информации вдоль нейрона необходимы потенциалы действия. Ганглиозные клетки обладают длинным (несколько сантиметров) аксоном и поэтому должны генерировать потенциалы действия для эффективного распространения сигнала в оптический нерв. Записи активности, сделаны от тел клеток. Локальные потенциалы возникают на дендритах в результате синаптических воздействий и пассивно распространяются к месту отведения.
Свойства потенциалов действияОдним из основных свойств потенциала действия является то, что это взрывное, пороговое событие, возникающее по закону «все или ничего». Потенциал действия возникает в ганглиозной клетке в тех случаях, когда приходящие от биполярных и амакриновых клеток сигналы достигают некоторого критического уровня (порога) мембранного потенциала. У потенциала действия (ПД) есть четко определенный порог, после достижения которого амплитуда и длительность ПД не зависят от параметров стимуляции. Большие по амплитуде стимулы не вызывают большие по амплитуде ПД, равно как и длинные стимулы не приводят к появлению более длинных ПД. Все фазы ПД должны быть полностью закончены до начала возникновения следующего ПД. После каждого ПД существует период вынужденного молчания (рефрактерный период), во время которого инициация ПД невозможна. Частота ритмических ПД определяется рефрактерным периодом.
Каждый импульс вызывает электрические токи, распространяющиеся пассивно перед ним по аксону. Хотя результирующий локальный потенциал быстро угасает с расстоянием, он все же превышает порог. Таким образом, ПД производит электрический стимул области аксома, в которую будет распространяться. Наиболее быстрые ПД распространяются по волокнам большого диаметра со скоростью около 120 метров в секунду (430 км/час), что и определяет возможность быстрой передачи информации на большие по сравнению с размером тела клетки расстояния.
ПД как нейронный кодУчитывая, что каждый ПД имеет фиксированную амплитуду, неясно, в чем же отражается величина стимула. Интенсивность кодируется частотой ПД. Более эффективный зрительный стимул вызывает большую деполяризацию и, как следствие, более высокую частоту генерации ПД в ганглиозной клетке Такое обобщение впервые было сделано Е. Эдрианом, который показал, что частота ПД в чувствительном окончании кожного нерва зависит от интенсивности стимула. Кроме того, он обнаружил, что более сильный стимул активирует большее количество чувствительных волокон.
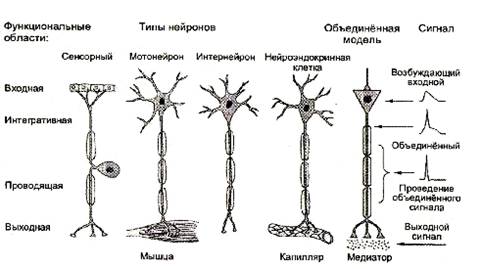
Синапсы: области межклеточной коммуникацииФоторецепторы влияют на биполярные клетки, которые влияют на ганглионарные клетки и так далее, что приводит к восприятию зрительного образа. Структура, через которую одна клетка передает информацию другой, известна как синапс. Механизм синаптической передачи представляет собой основную тему исследований в современной нейробиологии. Через синаптические взаимодействия нейроны, подобные ганглиозной клетке, интегрируют информацию о сигналах во многих фоторецепторах, производя на выходе новый собственный информационный сигнал для нервной сети.
Химически опосредованная синаптическая передачаПресинаптическая терминаль фоторецептора отделена от биполярной клетки щелью, заполненной экстраклеточной жидкостью. Это пространство слишком велико для прохождения токов, генерируемых фоторецептором. Вместо этого терминаль фоторецептора выделяет медиатор (иначе, трансмиттер или нейропередатчик), который хранится в пресинаптических пузырьках. Медиатор (в данном случае глутамат) диффундирует через синаптическую щель и реагирует со специфическими молекулами белка (рецепторами), которые находятся в постсинаптической мембране биполярной клетки. Следует отличать «хеморецепторы», реагирующие на молекулы, и «сенсорные рецепторы», реагирующие на внешние стимулы, например, фоторецептор. Медиаторы, синтезируемые и выделяемые нейроном, и рецепторы мембраны могут быть идентифицированы и визуализованы некоторыми методиками, включающими мечение антителами.
Активация молекул рецепторов биполярной клетки глутаматом приводит к появлению градуального локального потенциала, который распространяется по нейрону. Чем больше медиатора выделяется, тем выше его концентрация в щели, тем больше рецепторов активируется и тем больше локальный потенциал. Все эти события происходят быстро, примерно за 1 мс. Основные принципы синаптической передачи были впервые описаны Катцом, Куффлером и соавторами, которые использовали ответы мышечных рецепторов как очень чувствительную биомодель с хорошим разрешением по времени для измерения выделения медиатора.
Возбуждение и торможениеОсобенностью синаптической передачи, продемонстрированной на примере взаимодействия между фоторецептором и биполярной клеткой, является возможность торможения или возбуждения в зависимости от набора рецепторов в постсинаптической клетке. Например, один из видов глутаматных рецепторов на биполярной клетке реагирует на глутамат возбуждением (деполяризацией), которое распространяется до терминалей на другом конце клетки и приводит к высвобождению медиатора. Другой класс биполярных клеток содержит глутаматные рецепторы другого вида, которые реагируют торможением. В этом случае события происходят в той же последовательности, но приводят к уменьшению выброса медиатора.
Во всех нейронах нервной системы соотношение возбудительных и тормозных входов определяет возможность достижения порога инициации потенциала действия. Например, ганглиозная клетка получает и возбудительные, и тормозные входы. Если порог преодолен, то новый сигнал в виде ПД будет послан к высшим центрам, если нет, то сигнала не будет. В моторных клетках спинного мозга, например, возбудительные и тормозные влияния от разных волокон определяют, будет или нет произведено движение, контролируемое данными мотонейронами. Подобные мотонейроны получают около 10000 входов от волокон. Эти волокна выделяют медиаторы, которые приближают или отдаляют от порога возникновения ПД мембранный потенциал. Отдельные клетки в мозжечке получают более чем 100000 входов.
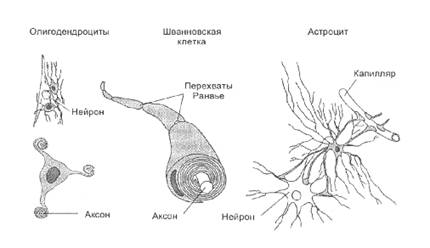
Электрическая передачаХотя основной способ передачи информации осуществляется через химическую передачу, некоторые клетки в сетчатке и других областях нервной системы связаны специализированными соединениями, в которых происходит электрическая передача информации. Пре- и постсинаптические мембраны в таких соединениях близко расположены и связаны каналами, которые соединяют внутриклеточное содержимое двух клеток. Такое соединение позволяет локальным потенциалам и даже потенциалам действия прямо распространяться от клетки к клетке без химического передатчика. Продукты метаболизма и красители также могут распространяться от клетки к клетке. В сетчатке есть так называемые горизонтальные клетки, которые электрически связаны таким способом. Благодаря этому свойству градуальные потенциалы могут распространяться от одной к другой горизонтальной клетке, сильно влияя на процесс переработки зрительной информации в сетчатке. Электрические синапсы обнаружены и между другими клетками тела, например между эпителиальными клетками, мышечными волокнами кишечника и сердца.
Химически опосредованная синаптическая передача информации очень лабильна. Основные изменения происходят в количестве медиатора, выделяемого при достижении пресинаптической терминали потенциалом действия или градуальным потенциалом. Фоторецептор сетчатки может служить примером: количество медиатора глутамата, выделяемого палочкой или колбочкой в ответ на стандартный световой стимул, может быть увеличено или уменьшено в зависимости от обратной связи на терминаль от горизонтальных клеток, которые получают входы от фоторецепторов. Эта цепь обратной связи играет критическую роль в адаптации глаза к различным уровням освещения.
Другие механизмы, влияющие на величину выброса медиатора, зависят от предыстории импульсной активности. Во время или после залпа импульсов в нейроне количество выделяемого им медиатора может существенно увеличиваться или уменьшаться в зависимости от частоты и длительности предшествующей импульсной активности. Модуляция эффективности может происходить и в постсинапсе. Долговременная и кратковременная пластичность находится в фокусе внимания нейробиологов.
Интегративные механизмыКаждый нейрон в центральной нервной системе учитывает все приходящие влияния и на их основе создает свое импульсное «послание» с новым значением. Термин интеграция впервые был применен Ч. Шеррингтоном, который также ввел в обиход термин «синапс».
Ганглиозные клетки сетчатки опять же могут служить примером способности к интеграции. С. Куффлер впервые показал, что ганглиозные клетки отвечают наиболее сильно на небольшое световое или темновое пятно размером в несколько рецепторов в определенной области сетчатки. Такое пятно вызывает отчетливый залп потенциалов действия Большое пятно, освещающее ту же область сетчатки, менее эффективно. Это происходит потому, что другая группа фоторецепторов, расположенная вокруг активированных, также реагирует на свет. Действие этих фоторецепторов тормозит активность ганглиозных клеток Суммация возбуждающего действия маленького пятна и тормозный эффект расположенных вокруг рецепторов приводят к тому, что ганглиозные клетки относительно слабо чувствительны к диффузному свету.
Таким образом, значение сигнала ганглиозной клетки не просто отражает «свет» или «темноту», но и соотносится с паттерном кон трастности светового стимула в поле зрения. Такой сложный сигнал возникает из-за того, что на каждую ганглиозную клетку приходят сигналы от многих фоторецепторов. Специфические связи, опосредованные биполярными, горизонтальными и амакриновыми клетками, определяют специфический паттерн светового стимула, оптимальным образом активирующий каждую конкретную ганглиозную клетку.
Сложность информации, передаваемой потенциалами действияЕще более сложную информацию о зрительных стимулах несут ПД нервных клеток новой коры, получающих сигнал через три переключения после сетчатки. Появление ПД в нейронах коры зависит от паттерна освещения сетчатки, который может быть специфичным для разных клеток. Например, один тип клеток избирательно отвечает на полоску света специфической ориентации (вертикальная, горизонтальная или наклонная), которая движется в определенном направлении в определенной части поля зрения. Параметры разрядов такой клетки не зависят от диффузного освещения или от появления полоски неоптимальной ориентации, или от движения в неправильном направлении. Таким образом, ПД в таком нейроне дают точную информацию о зрительном стимуле в высшие центры мозга. Подобная детализация значения, передаваемая стереотипными ПД, может быть объяснена точностью образования связей между клетками низкого порядка с кортикальными клетками и способом интеграции входных сигналов (суммация градуальных потенциалов).
Переработка информации может быть представлена в следующем виде:
сигнал фоторецептора несет информацию об изменении интенсивности освещения в данной области поля зрения;
сигнал ганглиозной клетки несет информацию о контрасте;
сигнал кортикального нейрона несет информацию о наличии ориентированной полоски света.
Сложная интеграция информации происходит в других сенсорных системах. Например, направление и локализация механического стимула кончиков пальцев служат избирательным стимулом для конкретных нейронов той области неокортекса, в которой происходит переработка информации о направлении тактильной стимуляции.
Выводы
Можно сделать два важных заключения о принципах переработки информации в нервной системе: (1) нервные клетки играют роль составляющих элементов для «построения» восприятия; (2) значение сигнала нейрона может быть очень сложным и зависеть от входных сигналов.
Литература
Шлегель Г.Г. История микробиологии.
Лотова Л.И. Морфология и анатомия высших растений.
Алексеев В.И., Каминский В.А. Прикладная молекулярная биология.
Галимов Э.М. Феномен жизни. Происхождение и принципы эволюции.
Блюменфелъд Л.А. Решаемые и нерешаемые проблемы биологической физики.
Барышников А.Ю., Шишкин Ю.В. Иммунологические проблемы апоптоза.
Похожие работы
... плотно смыкаются друг с другом и практически полностью обвертывают кровеносный сосуд. Подобная изоляция сосуда является одним из способов формирования гематоэнцефалического барьера - граници между кровью и нервной тканью, закрытой для многих находящихся в крови веществ. Другие отростки астроцита почти целиком обвертывают тела нейронов. Если нейрон возбуждается длительно, вокруг него повышается ...
... нервами. БИЛЕТ № 12 Общая морфология заднего мозга Задний мозг, включающий продолговатый мозг и Варолиев мост представляют собой филогенетически древнюю область центральной нервной системы, сохраняя черты сегментарного строения. В заднем мозге локализованы ядра и проводящие восходящие и нисходящие пути. По проводящим путям в задний мозг поступают афферентные волокна от вестибулярных и ...
... мозгу (1), где через вставочный нейрон передаются на эфферентные волокна (эфф. нерв), по которым доходят до эффектора. Пунктирные линии - распространение возбуждения от низших отделов центральной нервной системы на ее вышерасположенные отделы (2, 3,4) до коры мозга (5) включительно. Наступающее вследствие этого изменение состояния высших отделов мозга в свою очередь воздействует (см. стрелки) на ...




... подход к оценке механизмов работы высших отделов головного мозга с использованием условных рефлексов был столь успешным, что позволил Павлову создать новый раздел физиологии — «Физиологию высшей нервной деятельности», науку о механизмах работы больших полушарий головного мозга. БЕЗУСЛОВНЫЕ И УСЛОВНЫЕ РЕФЛЕКСЫ Поведение животных и человека представляет собой сложную систему взаимосвязанных ...
0 комментариев