Для того чтобы вычислить точное значение мембранного потенциала для модели идеальной клетки, необходимо принять во внимание отдельные ионные токи, протекающие через мембрану. Входящий натриевый ток (iNa) зависит от величины движущей силы для ионов натрия (Vm-ENa), а также от натриевой проводимости мембраны (gNa). Проводимость пропорциональна среднему количеству натриевых каналов, находящихся в открытом состоянии при потенциале покоя: чем больше открытых каналов, тем выше проводимость. Таким образом, натриевый ток равен:
![]()
Тот же подход справедлив и для калия и хлора:
![]()
Если принять iCl, = 0 (т.е. ионы хлора находятся в равновесии), то, для сохранения неизменного значения мембранного потенциала, калиевый и натриевый токи должны быть равны по величине и противоположны по направлению:
![]()
Рассмотрим это уравнение более подробно. Предположим, что gK значительно больше gNa. Тогда, для того чтобы токи были равны, движущая сила для выхода калия должна быть значительно меньше движущей силы для натриевого входа. Другими словами, мембранный потенциал должен быть ближе к ЕK, чем к ENa. Соответственно, если значение gNa велико, то мембранный потенциал будет ближе к ENa.
Преобразовав выражение, получим формулу для мембранного потенциала:
![]()
Если по той или иной причине ионы хлора не находятся в равновесии, то формула приобретает несколько более сложный вид:
![]()
Вышеприведенные расчеты были первоначально проведены Голдманом) и, независимо от него, Ходжкином и Катцем), с той лишь разницей, что вместо равновесных потенциалов и проводимостей они оперировали ионными концентрациями снаружи ([Na] 0,. .) и внутри ([Na] i,. .) клетки, а также проницаемостью мембраны для каждого из ионов (pNa···):
![]()
Так же как и ранее, хлорные компоненты выражения не рассматриваются, если ионы хлора находятся в равновесии. Уравнение называется "уравнением ГХК" по фамилиям авторов, или уравнением постоянного поля, поскольку одним из допущений при выводе уравнения является равномерное распределение градиента напряжения (т.е. "поля") на всем протяжении мембраны. Уравнение ГХК полностью аналогично уравнению проводимостей, на его основании можно сделать те же предсказания: когда проницаемости для натрия и хлора малы по сравнению с калиевой проницаемостью, хлорный и натриевый компоненты уравнения становятся пренебрежимо малы, и мембранный потенциал стремится к значению равновесного потенциала для калия: Vm = 58 log ([K] 0/ [K] 1). При увеличении натриевой проницаемости мембранный потенциал сдвигается в сторону натриевого равновесного потенциала.
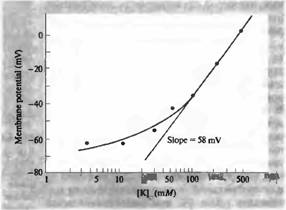
Рис.1. Зависимость мембранного потенциала от внеклеточной концентрации калия на аксоне кальмара (полулогарифмическая шкала). Прямая линия проведена под углом наклона, равным 58 мВ на десятикратное изменение внеклеточного уровня калия, в соответствии с уравнением Нернста. Благодаря наличию мембранной проницаемости для натрия, экспериментальные данные расходятся с прямой линий, особенно при низких концентрациях калия.
Из уравнения постоянного поля следует один важный принцип. Мембранный потенциал зависит от проводимости (или проницаемости) мембраны для важнейших ионов, а также от их равновесных потенциалов. В реальных клетках проводимости для калия и хлора сравнительно· высоки, поэтому потенциал покоя клетки близок к равновесным потенциалам для этих двух ионов. При увеличении натриевой проводимости, например, во время потенциала действия или возбуждающего постсинаптического потенциала, мембранный потенциал сдвигается в сторону натриевого равновесного потенциала.
Потенциал покоя
Несмотря на то, что во многих случаях уравнение ГХК весьма полезно, оно не дает возможности правильно вычислить потенциал покоя. Дело в том, что условие равенства суммарного тока нулю неправомочно: третье условие равновесия, а именно требование, чтобы каждый отдельный ионный ток равнялся нулю, не выполняется. В результате клетка будет постепенно терять калий и наполняться натрием и хлором. В реальных клетках концентрации натрия и калия поддерживаются на постоянном уровне при помощи натрий-калиевых насосов. В противовес непрерывному потоку натрия внутрь клетки и калия наружу, эти насосы переносят оба эти иона в определенной пропорции в направлении, противоположном их пассивному движению (рис.1). Следовательно, для поддержания клетки в стабильном состоянии затрачивается метаболическая энергия.
Чтобы получить более полную формулу потенциала покоя, необходимо принять во внимание как пассивное движение ионов, так и их активный транспорт, осуществляемый насосами. Рассмотрим сначала пассивное движение ионов натрия и калия через мембрану:
![]()
Отказавшись от предположения о том, что токи эти равны по величине и противоположны по направлению, но зная соотношение между ними, можно получить уравнение мембранного потенциала, исходя из равновесных потенциалов для натрия и калия, а также их относительных проводимостей. Здесь уже нужно учитывать влияние насоса. Поскольку насос поддерживает внутриклеточные концентрации натрия и калия на постоянном уровне, перенося ионы в соотношении 3: 2, то и пассивные токи должны находиться в том же соотношении: iNa/iK = 3/2. Имеем следующее выражение:
![]()
Соотношение имеет отрицательную величину, потому что направления натриевого и калиевого токов противоположны. Преобразовав выражение, получим:
![]()
Это уравнение похоже на полученное выше для модели идеальной клетки, и на его основании можно сделать то же предсказание: значение мембранного потенциала будет определяться относительными величинами gNa и gК. Отличие настоящей формулы состоит в том, что члены, относящиеся к калию, умножены на 1,5. Благодаря этому фактору мембранный потенциал будет ближе к EK. Таким образом, движущая сила для входа натрия возрастает, а для выхода калия, наоборот, уменьшается. В результате и пассивные токи натрия и калия будут находиться в соотношении 3: 2, а не 1: 1.4,Обобщая вышесказанное, можно сделать вывод, что потенциал покоя - это такое значение мембранного потенциала, при котором входящий натриевый ток не равен по величине выходящему калиевому, а превосходит его в 1,5 раза. Пассивные токи обоих ионов определяются их равновесными потенциалами и проводимостями, в то время как соотношение токов 3: 2 продиктовано стехиометрическими характеристиками насоса.
Задачу по выводу формулы потенциала покоя для реальных клеток впервые взяли на себя Муллинс и Нода. С помощью внутриклеточных микроэлектродов они исследовали влияние изменения ионных концентраций на мембранный потенциал мышечной клетки. Подобно Голдману, Ходжкину и Катцу, они получили выражение, связывающее мембранный потенциал с проницаемостями и концентрациями отдельных ионов. Результат аналогичен только что полученному нами уравнению, основанному на проводимостях и равновесных потенциалах:
![]()
где r - абсолютное значение соотношения ионного транспорта (3: 2). Уравнение точно описывает потенциал покоя при условии, что все остальные ионы, проникающие сквозь мембрану (такие как хлор), находятся в равновесии.
Распределение хлора
Насколько применимы подобные рассуждения для хлора? Как и для других ионов, суммарный ток хлора должен быть равен нулю. Как показано выше, равновесие ионов хлора достигается путем простого изменения его внутриклеточной концентрации, без изменения при этом мембранного потенциала. Тем не менее, во многих клетках существуют транспортные системы для хлора. В аксоне кальмара и в мышце активные транспортные системы переносят ионы хлора внутрь клетки; во многих нервных клетках, напротив, хлор выводится из клетки (см. рис.1). В результате активного транспорта хлора в клетку, его внутриклеточная концентрация возрастает, что приводит к усилению выходящего пассивного тока утечки хлора, равного по величине активному входящему току. Транспорт хлора из клетки наружу производит обратное действие.
Электрическая модель мембраны
Потенциалы ENa, ЕK и ЕCl показаны в виде батарей, а проводимости для натрия, калия и хлора изображены в виде сопротивлений (резисторов). Пассивные токи через сопротивления равны и противоположны токам, генерируемым соответствующими насосами, поэтому суммарный ток через мембрану для каждого иона равен нулю.
Ожидаемые значения мембранного потенциала
Для аксона кальмара соотношение констант проницаемости для натрия и калия приблизительно равно 0,04: 1,0. Воспользовавшись этими данными, а также значениями ионных концентраций, получим потенциал покоя в морской воде:
![]()
Становится очевидным, почему при изменении внеклеточной концентрации калия мембранный потенциал не строго подчиняется уравнению Нернста для калия. Если сравнить слагаемые в знаменателе, относящиеся к калию (1,5 x 10 = 15) и к натрию (0,04 x 460 = 18,4), то получится, что вклад калия составляет всего около 45%. По этой причине, увеличение внеклеточной концентрации калия вдвое не приведет к удвоению числителя (как было быв случае полного соответствия с уравнением Нернста), и, следовательно, эффект изменения уровня калия на мембранный потенциал меньше, чем в идеальной ситуации, когда калий является единственным проводящим ионом. Когда внеклеточная концентрация калия возрастает до существенных значений (100 ммоль), то роль калиевого компонента в знаменателе становится подавляющей, и кривая зависимости приближается к своему теоретическому углу наклона (58 мВ на 10-кратное увеличение концентрации калия). Многие калиевые каналы обладают чувствительностью к мембранному потенциалу и открываются при деполяризации, вызванной увеличением внеклеточной концентрации калия. В результате подобного увеличения проницаемости для калия вклад натрия в формирование мембранного потенциала дополнительно снижается.
Вообще, потенциал покоя нервных клеток приблизительно равен - 70 мВ. В некоторых клетках, например в скелетной мышце позвоночных, потенциал покоя может быть - 90 мВ или даже больше, что объясняется значительным превосходством калиевой проводимости над натриевой. Натриевая проводимость глиальных клеток настолько мала, что их потенциал покоя практически равен калиевому равновесному потенциалу. В таких клетках, как, например, нейроны ганглиев пиявки или рецепторы сетчатки, натриевая проводимость достаточно велика, а потенциал покоя составляет всего - 40 мВВклад натрий-калиевого насоса в мембранный потенциал.
Натрий-калиевый насос является электрогенным, потому что за каждый цикл его работы из клетки выводится один суммарный положительный заряд, тем самым увеличивая отрицательный заряд на внутренней поверхности мембраны клетки. Насколько велик этот вклад в заряд на мембране? Чтобы ответить на этот вопрос, представим себе, что насос не электрогенен, а нейтрален, то есть соотношение ионов, переносимых насосом внутрь и наружу клетки, равно единице. С учетом этого условия проведем вычисления:
![]()
Поскольку полученный потенциал покоя на 6 мВ менее отрицателен, то можно сделать вывод о том, что вклад насоса в потенциал покоя равен - 6 мВ. При условии транспорта ионов в соотношении 3: 2 вклад насоса в потенциал покоя не может превысить - 11 мВ. При остановке ионообмена его влияние на величину потенциала покоя прекращается, и потенциал покоя постепенно снижается по мере накопления в клетке ионов натрия и выхода из нее калия.
Ионные каналы, участвующие в формировании потенциала покоя
Наличие проводимости для натрия, калия и хлора в состоянии покоя было показано во многих клетках. Интересно отметить, что ионные каналы, обеспечивающие эти проводимости, до сих пор не были определены ни в одном типе клеток. Кандидаты на роль калиевых каналов, активных в диапазоне потенциала покоя, различны в зависимости от типа клеток. Среди них есть каналы, активируемые внутриклеточными катионами: натрий-активируемые и кальций--активируемые калиевые каналы. Кроме того, многие нервные клетки обладают так называемым калиевым каналом типа "М", которые открыты при потенциале покоя и закрываются при участии внутриклеточных посредников. Активация большого количества потенциал-зависимых калиевых каналов (то есть "замедленного выпрямителя" - delayed rectifier, и каналов типа "А") при потенциале покоя маловероятна. Однако, даже 0,1-1% от их общего числа было бы достаточно, чтобы создать проводимость, сравнимую с проводимостью в покое.
Конкретные носители натриевой проводимости в нервных клетках также неизвестны. В некоторой степени, вход натрия может происходить через калиевые каналы, для большинства из которых проницаемость для натрия составляет от 1 до 3% от калиевой проницаемости. Кроме того, открытые катионные каналы могут способствовать как выходу калия, так и входу натрия, поскольку большая часть этих каналов не обладает высокой селективностью к калию по сравнению с натриевой. Дополнительный источник входящего в клетку натрия - это натрий--зависимые вторичные активные транспортные системы. Наконец, потенциал--чувствительные натриевые каналы также вносят свой вклад в натриевую проводимость в покое, поскольку эта проводимость блокируется тетродотоксином, хотя и в небольшой степени).
Хлорные каналы семейства CLC широко распространены в нервных и мышечных клетках. Наличие хлорных каналов обеспечивает стабилизацию мембранного потенциала. Эти каналы также участвуют в установлении внутриклеточной концентрации хлора, взаимодействуя с системами хлорного транспорта. При низком уровне экспрессии каналов CLC, например, в нейронах эмбрионального гиппокампа, значение ЕCl менее отрицательно по сравнению с потенциалом покоя, благодаря вкладу активного транспорта ионов хлора внутрь клетки и их накоплению в цитоплазме. В нейронах взрослого животного экспрессия каналов CLC более выражена, и мембранная проводимость для хлора выше, поэтому накопление внутриклеточного хлора невозможно, и ЕСl равен мембранному потенциалу. В клетках центральной нервной системы до 10% проводимости мембраны при потенциале покоя обеспечивается за счет хлорных каналов.
Изменения мембранного потенциалаРавновесный мембранный потенциал представляет собой тот фоновый уровень, на котором происходят все изменения мембранного потенциала. Как они происходят? Обычно кратковременные изменения мембранного потенциала (подобные тем, которые наблюдаются при взаимодействии нейронов между собой) происходят за счет временных изменений проницаемости мембраны. Как мы знаем из уравнения постоянного поля, увеличение натриевой проницаемости (также как и снижение калиевой) приводит к деполяризации, то есть к смещению мембранного потенциала в сторону натриевого равновесного потенциала. Напротив, увеличение калиевой проницаемости вызывает гиперполяризацию. Еше один важный для сигнальных процессов клетки ион - это кальций. Внутриклеточная концентрация кальция очень низка, а ЕСа в большинстве клеток превышает +150 мВ. Следовательно, увеличение кальциевой проницаемости должно приводить к входу кальция в клетку и деполяризации мембраны.
Особый интерес представляет влияние хлорной проницаемости на мембранный потенциал. Как было отмечено выше, хлор вносит незначительный вклад в потенциал покоя. Внутриклеточная концентрация хлора подстраивается под новые условия за счет работы существующих хлорных транспортных механизмов. Влияние кратковременного увеличения хлорной проницаемости может выражаться как в гиперполяризации, так и в деполяризации, в зависимости от того, положителен или отрицателен ЕС1 по отношению к потенциалу покоя. Это, в свою очередь, зависит от работы транспортного механизма, либо наполняющего клетку хлором, либо выводящего его наружу. В любом случае, влияние хлора на мембранный потенциал обычно невелико. Тем не менее, увеличение хлорной проницаемости может иметь важное значение для регулирования сигнальных процессов, поскольку оно поддерживает мембранный потенциал в диапазоне ЕCl и за счет этого ослабляет способность других процессов изменить мембранный потенциал.
Поддержание мембранного потенциала на неизменном уровне играет важную роль в регуляции возбудимости таких клеток, как скелетная мышца, хлорная проводимость которой в покое достаточно велика. В таких клетках кратковременный вход положительных ионов вызовет меньшую деполяризацию в присутствии хлорной проницаемости, нежели в ее отсутствии, поскольку их вход отчасти балансируется входом хлора через уже открытые каналы. Мутации хлорных каналов, снижающие хлорную проницаемость мембраны, приводят к некоторым мышечным заболеваниям. Потерявшие нормальную высокую хлорную проводимость мышцы находятся в состоянии повышенной возбудимости (myotonic.
Выводы
Нервные клетки обладают высокой внутриклеточной концентрацией калия и низкой концентрацией натрия и хлора, так что калий стремится выйти из клетки, а натрий и хлор стремятся войти в нее. Стремлению калия и хлора проникнуть через мембрану в направлении концентрационного градиента противостоит мембранный потенциал.
В модели идеальной клетки, обладающей только калиевой и натриевой проводимостями, мембранный потенциал может в точности уравновесить концентрационные градиенты таким образом, чтобы суммарный ток обоих ионов равнялся нулю. В этом случае мембранный потенциал соответствует равновесному потенциалу для калия и хлора.
Изменение внеклеточного уровня калия приводит к изменению его равновесного потенциала и, следовательно, мембранного потенциала. Напротив, изменение внеклеточной концентрации хлора приводит в конечном итоге к изменению его внутриклеточного уровня, так что сдвиг хлорного равновесного потенциала и мембранного потенциала от своих первоначальных значений носит кратковременный характер.
Мембрана реальных клеток обладает также натриевой проводимостью. В результате в состоянии покоя происходит постоянная диффузия натрия внутрь, а калия наружу клетки. Эти токи в точности уравновешиваются активным ионным транспортным механизмом - натрий-калиевым насосом, работающим по схеме 3 иона натрия к 2 ионам калия. В этих условиях, величина мембранного потенциала зависит от натриевого и калиевого равновесных потенциалов, относительных проводимостей для этих ионов, а также от соотношения ионов в цикле насоса.
Благодаря тому, что натрий-калиевый насос переносит больше положительных ионов наружу, чем внутрь клетки, он вносит непосредственный вклад в мембранный потенциал в размере нескольких мВ.
В зависимости от особенностей хлорных транспортных механизмов, равновесный потенциал для хлора может быть как более, так и менее отрицательным по отношению к потенциалу покоя. Несмотря на незначительное влияние распределения хлора на потенциал покоя, наличие высокой хлорной проводимости необходимо для поддержания электрической стабильности клетки.
Литература
1. Блюменфелъд Л.А. Решаемые и не решаемые проблемы биологической физики.
2. Барышников А. Ю, Шишкин Ю.В. Иммунологические проблемы апоптоза.
3. Крутько В.Н., Славин М.Б., Смирнова Т.М. Математические основания геронтологии.
Похожие работы
... Шоттки, и механизма их функционирования на основе разработанной математической модели двойного электрического слоя на границе мембрана/раствор. 2. Основные закономерности переноса ионов в трехслойной мембранной системе при интенсивных токовых режимах, а именно: а) утверждение, что одновременный учет трех факторов: диссоциации воды, пространственного заряда и сопряженной конвекции объясняет ...
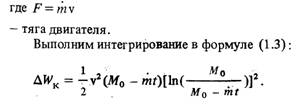
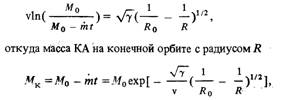
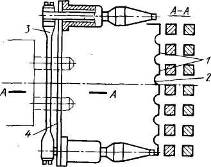
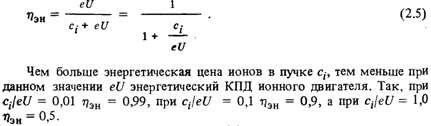
... ионные источники Как уже говорилось, в контактных ионных источниках образование ионов происходит в результате поверхностной ионизации. В ионных электрических ракетных двигателях применяются контактные ионные источники с пористым ионизатором. Пористые ионизаторы изготавливаются различными способами, например, тонкие каналы-поры могут быть пробиты лазерным или электрическим лучом. В качестве ...
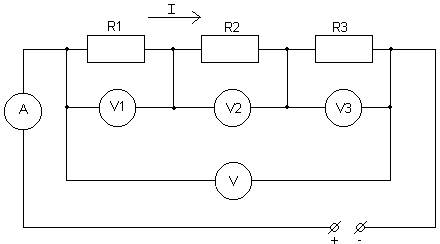
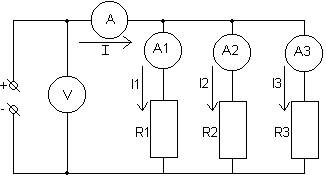
... неровностей на поверхности анода, т.е. происходит его полировка. 2 Расчётная часть 2.1Задание на курсовую работу Расчет разветвлённой электрической цепи постоянного тока. Для заданной электрической цепи необходимо: 1) Записать систему уравнений по законам Кирхгофа (без расчетов); 2) Определить все токи и ...
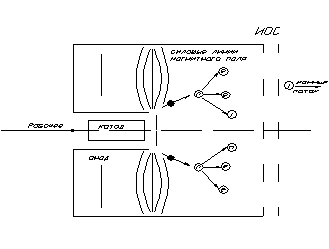
... и описание теоретического чертежа двигателя Плазменный ионный движитель представляет собой устройство, в котором создание тяги основано на принципе ускорения заряженных частиц. Общий вид плазменного-ионного двигателя представлен на чертеже ХАИ.06.441п.11.TЧ.04. Заряженные частицы образуются в части движителя, которая называется газоразрядной камерой (ГРК) (6). В состав ГРК входят катодный ...
0 комментариев