Гормоны, плохо проникающие в клетку (белково-пептидные, катехоламины), соединяются с рецепторами на клеточной мембране и вызывают внутриклеточные эффекты через вторые посредники.
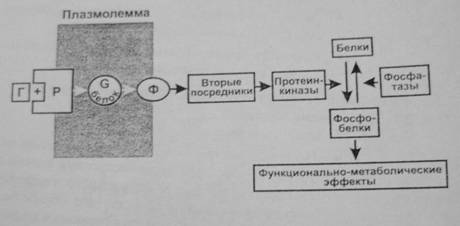
Большинство гормонов после присоединения к рецепторам активируют регулируемый гуанилнуклеотидами G-белок мембраны. Активация состоит в обмене ГДФ на ГТФ в α-субъединице G-белка. При этом происходит его распад на α-субъединицу и βγ-димер, имеющие самостоятельные регуляторные эффекты. Более изучено действие α-субъединицы, являющейся ферментом ГТФазой. Гидролиз ГТФ до ГДФ освобождает энергию, используемую для передачи регуляторного сигнала. ГДФ-α-субъединица соединяется с βγ-димером, вновь образуя ГДФ—G-белок. Имеется несколько видов G-белков (GS, Gi ,Gq ,GO), которые влияют на ферменты (например, аденилатциклазу, фосфолипазу С), образующие различные вторые посредники (например, цАМФ, инозитолтрифосфат, диацилглицерол). При этом G-белки могут как активировать (GS), так и ингибировать (Gi) образование вторых посредников. Вторые посредники, в свою очередь, активируют соответствующие им протеинкиназы, фосфорилирующие различные клеточные белки по остаткам серина и треонина, что изменяет их функциональную активность (чаще увеличивает). Наряду с этим активированные G-белки оказывают выраженное влияние на проницаемость кальциевых и калиевых каналов мембраны.
В действии гормонов, плохо проникающих в клетки, можно выделить 4 системы: аденилатциклазную, фосфоинозитидную, гуанилатциклазную и тирозинкиназную.
Аденилатциклазная система. Эту систему используют гормоны, осуществляющие свой эффект через аденилатциклазу и синтез второго посредника—циклического АМФ (3,5-цАМФ). Действие гормона может, как активировать, так и тормозить его образование.
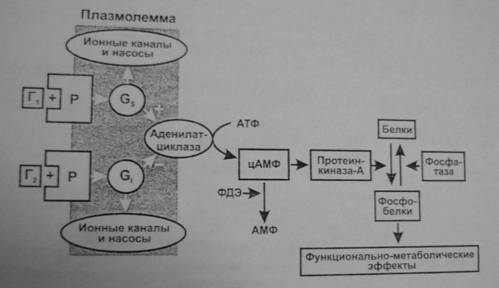
- Комплекс «гормон + рецептор» активирует GS–белок, который активирует аденилатциклазу и образование цАМФ (например, так действуют адреналин через β1/β2-рецепторы, АДГ через V2-рецепторы, глюкагон, АКТГ, ТТГ, ФСГ, ЛГ, МСГ, кортиколиберин, соматолиберин, кальцитонин, хориогонадотропин человека, паратгормон).
- Комплекс «гормон + рецептор» активирует Gi-белок, который тормозит аденилатциклазу и образование цАМФ (например, так действуют адреналин через α2-рецепторы, ангиотензин II через АТ-рецепторы, аденозин через А1-рецепторы, соматостатин, опиоиды через μ-, δ- и κ-рецепторы, ацетилхолин через М2-холинорецепторы гладкомышечных клеток).
- цАМФ влияет на функции клетки через изменение активности протеинфосфокиназы-А, фосфорилирующей различные клеточные белки: мембранные (изменяется ее проницаемость), ферменты (изменяется метаболизм клетки), ядерные (изменение транскрипции), рибосом (изменение трансляции), микротрубочек (двигательные эффекты, изменение формы клетки) и др.
Фосфоинозитидная система. Эту систему используют гормоны, которые, действуя через Gq-белок, активируют в мембране фосфолипазу С.
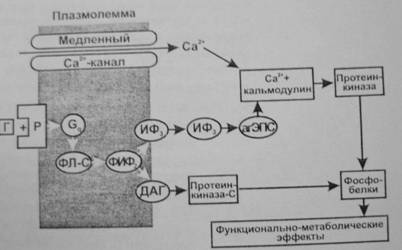
Она образует из фосфолипида мембраны (фосфатидилинозитолдифосфата) два посредника—инозитол—1,4,5—трифосфат (ИФ3) и 1,2—диацилглицерол (ДАГ).
- ИФ3, переходя в цитоплазму, действует на рецепторы гладкой ЭПС, что приводит к открыванию Са2+-каналов и выходу Са2+ в цитозоль. Са2+ при этом выступает в качестве третьего посредника, который связывается с белком кальмодулином и активирует Са2+-кальмодулинзависимую протеинкиназу, фосфорилирующую эффекторные белки, влияющие на функции клеток, их пролиферацию и дифференцировку (регулирует активность аденилатциклазы и синтез цАМФ, фосфолипазы—А2 и синтез простагландинов, тромбоксана, простациклина, гуанилатциклазы и синтез цГМФ, фосфодиэстеразы и разрушение цАМФ и цГМФ; участвует в регуляции секреции инсулина, гормонов щитовидной железы, гипофиза и надпочечников, кишечной секреции и секреции нейромедиаторов). (Роль Са2+ как посредника действия гормонов обусловлена его сильным влиянием при низких концентрациях 10-8—10-6 М на метаболизм и функции клетки. Гормоны могут увеличивать концентрацию Са2+ в цитозоле как в результате открывания Са2+-каналов клеточной мембраны, так выхода Са2+ из гладкой ЭПС. В клетке есть специальные белки, связывающие Са2+ при низкой концентрации—кальмодулин, гуанилатциклаза, тропонин С и др. Присоединение Са2+ к этим белкам изменяет их конфигурацию и свойства. Это приводит к изменению многочисленных функций клеток—метаболической, секреторной, сократительной, биоэлектрической и др.)
- ДАГ, оставаясь на внутренней поверхности мембраны, активирует протеинкиназу С, фосфорилирующую эффекторные белки (например, активирует киназу легкой цепи миозина), а также синтез простагландинов из арахидоновой кислоты, что изменяет функции клетки.
- Через ИФ3 и ДАГ действуют адреналин (α-рецепторы), вазопрессин (V1), ацетилхолин (М1), ангиотензин II (AT1), гистамин (Н1), серотонин (5-НТ2), холецистокинин, окситоцин, ВИП и др.
Гуанилатциклазная система. Некоторые гормоны (например, ацетилхолин, цитокины) действуют через гуанилатциклазу, образующую второй посредник—циклический ГМФ (цГМФ). Гуанилатциклаза существует в двух формах—мембраносвязанной и растворимой в цитозоле.
- Мембраносвязанная гуанилатциклаза является частью рецептора плазмолеммы, ее активация гормоном (например, Na+-уретическим) увеличивает образования цГМФ, который стимулирует протеинкиназу G, фосфорилирующую белки.
- Растворимая в цитозоле гуанилатциклаза содержит в своем составе гемм, с которым связывается сигнальная молекула – NO (оксид азота образуется из аргининав ряде клеток—эндотелии, нейронах и др. – под действием ацетилхолина, механических раздражителей и др.). Активированная оксидом азота гуанилатциклаза через протеинкиназу G фосфорилирует белки (например, так действует ацетилхолин, вызывая расширение сосудов).
Тирозинкиназная система характеризуется особыми рецепторами, которые путем фосфорилирования и дефосфорилирования сигнальных белков по тирозиновым остаткам, регулируют процессы роста, дифференцировки, выживания, метаболизма и смерти клеток. Эту систему используют некоторые гормоны—инсулин, инсулиноподобные факторы роста (или соматомедины), факторы роста эпидермиса, тромбоцитов и др. Многие рецепторы к таким гормонам содержат в своём составе тирозинкиназу, которая при действии гормона аутофосфорилирует внутреннюю часть рецептора по остаткам тирозина. Активированный таким образом рецептор передает через сложный и недостаточно изученный комплекс посредников фосфорилирующее и дефосфорилирующее влияние по остаткам тирозина на многие сигнальные молекулы цитоплазмы и ядра. Фосфорилирование и дефосфорилирование белков может приводить как к их активации, так и инактивации.
Похожие работы
... . И.П. Павлову, первому из русских ученых, 7 октября 1904 г. была присуждена Нобелевская премия в знак признания его работ по физиологии пищеварения. Потребность организма в пище проявляется в виде физиологической реакции голода. У человека голод приобретает выраженную субъективную окраску — от относительного безразличия к пище до яркой эмоциональной реакции. Физиологической основой голода ...
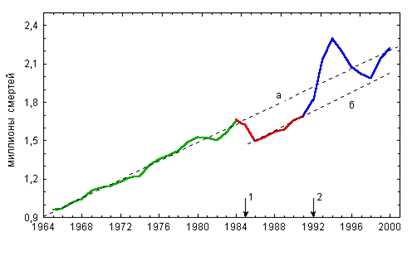
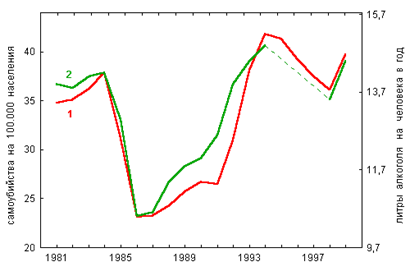
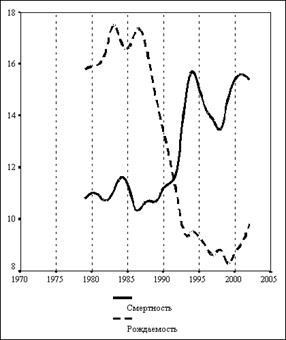
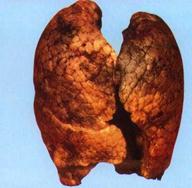
... в состоянии продолжать жить, несмотря на указания совести о неправильности жизни…». На этом тезисе мы закончили предыдущую главу книги. Не развивая его, поскольку это не входит в достаточно узкую заявленную тематику оружия геноцида, обратимся к внутренним механизмам, которые включаются у людей после принятия психотропов. Мы знаем от чего люди убегают (от своей совести). Но за чем же люди гонятся? ...
табилизирует мембрану тучных клеток препятствует процессу дегрануляции тучных клеток применяется для профилактики приступов бронхиальной астмы снижает давление в малом кругу кровообращения улучшает кровоток в почках улучшает кровоток в сердце применяется для купирования приступов бронхиальной астмы 80.Укажите 2 нейротропных средства, ослабляющих сократительную активность миометрия 4 2 2 1 2 ...
... механизмов, формирующих иммунные барьеры против чужеродных веществ; синтезирует некоторые витамины, частично используемые организмом: В3, В9, В5, В6, В8, Н и К; и другие функции. Микроорганизмы толстого кишечника для своего питания используют непереваренные остатки пищи человека, вызывая глубокое расщепление органических веществ и образуя разнообразные продукты, в том числе токсические и ...
0 комментариев