Физиология и поведение животных в изменчивой среде
Содержание
Введение
1. Толерантность
2. Акклиматизация
3. Биологические часы
4. Репродуктивное поведение н физиология
5. Зимняя спячка
6. Миграция
7. Лунные и приливно-отливные ритмы
8. Циркадианные ритмы и суточное распределение активности
Введение
В этой работе описывается, как животные отвечают на физиологические требования, связанные с поддержанием жизни и воспроизведением в изменчивой среде.
1. Толерантность
Жизнь, весьма вероятно, возникла в море. По сравнению с другими биомами биомы морской среды сравнительно устойчивы. Флуктуации таких физических факторов, как температура и содержание кислорода, здесь невелики, поэтому внутренняя среда многих морских беспозвоночных мало подвержена нарушениям. Такие животные обычно являются конформерами в том смысле, что состояние их организма соответствует состоянию внешней среды и они не могут жить в изменчивых условиях. Так, например, соленость жидкостей тела у многих морских беспозвоночных такая же, как у морской воды. У других животных, называемых регуляторами, функции организма сравнительно независимы от флуктуации внешней среды. Эта способность послужила предпосылкой для заселения пресной воды и суши.
Каждый вид обладает своей характерной способностью переносить крайние значения внешних факторов, например температуры и влажности. На многих морских беспозвоночных действует изменение солености воды, потому что в жидкостях их тела в норме концентрация соли почти такая же, как в морской воде, и их ткани приспособлены хорошо функционировать именно в таких условиях. Если поместить их в менее соленую среду, вода извне будет осмотически поступать в их ткани. В более соленой среде произойдет обратное. Для животных, не способных контролировать проникновение воды в тело, условия существования соответствуют пределам переносимой солености.
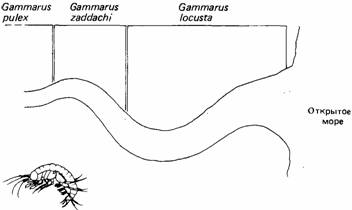
Среда обитания животного ограничена его толерантностью. Например, разные виды бокоплавов рода Gammarus приурочены к разным участкам эстуариев из-за своей разной солеустойчивости. Как показано на рис.1, Gammarus locusta весьма толерантен к соленой воде и встречается ближе к устью; у Gammarus zaddachi толерантность умеренная, и обычно он обитает в реке на участке от 13 до 20 км от моря. Gammarus pulex является настоящим пресноводным видом и полностью отсутствует в местах, где как-либо ощущается влияние прилива или соленой воды. В этом примере каждый вид переносит только ограниченные пределы солености и приспособлен отбором к условиям, в которых не живут другие представители этого рода.
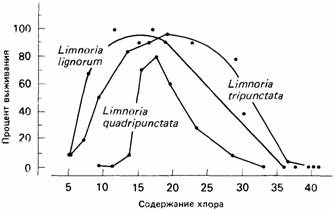
При построении графиков зависимости выживания, или приспособленности, от существенных переменных окружающей среды обычно получаются колоколообразные кривые наподобие приведенных на рис.2. Только немногие животные выдерживают крайне высокие или крайне низкие значения экологических параметров, а остальные вынуждены скапливаться в области их средних величин.
|
| Рис.1. Распределение но реке трех весьма близких друг к другу видов бокоплавов Gummarus в зависимости от концентрации соленой воды. Степень опреснения воды показана густотой точек. |
Такие кривые показывают не только пределы и диапазон толерантности вида, но также оптимальные значения переменных среды. Подобным образом можно представить толерантность животных применительно к любым градиентам среды.
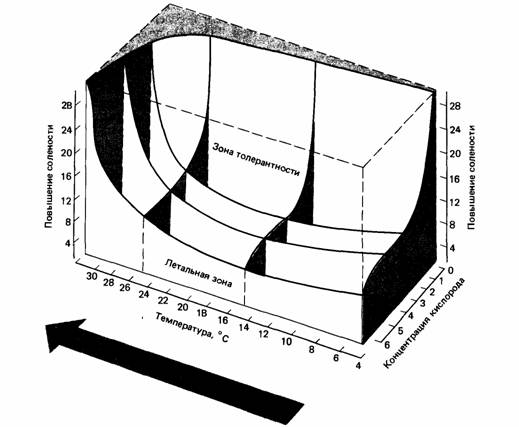
У конформеров толерантность часто непосредственно связана с физиологическим состоянием животного. Например, если окружающая температура 40°С для них смертельна, то это потому, что внутренняя температура около 40°С вызывает биохимический распад. Но на толерантность к данному фактору влияют значения и других переменных среды. Действуя в совокупности, экологические факторы могут убить животное при интенсивностях. которые, взятые по отдельности, не были бы губительны для него. Так, например, для американского омара Homarus americanus температура в 32°С смертельна при солености около 3% и содержании кислорода в воде 6,5 мг/л. Если содержание кислорода упадет до 2,9 мг/л, температурный предел выживания снизится до 29°С. В одной работе омаров подвергали действию 27 комбинаций температур, солености и содержания кислорода. Полученные результаты представлены в виде трехмерного графика, который показывает, как взаимодействие факторов влияет на пределы толерантности вида.
|
| Рис.2. Устойчивость трех видов сверлящих дерево морских изопод к разному постоянному содержанию хлора. |
|
|
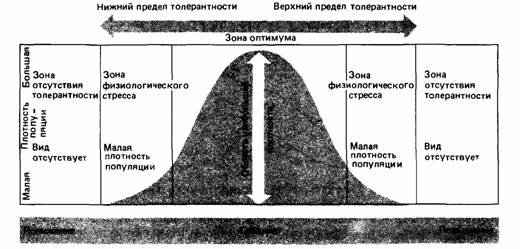
| Рис.3. Распределение плотности популяции вдоль экологического градиента. |
|
|
| Рис.4. Трехмерное изображение границ летальных условий для американского омара при разных сочетаниях температуры, солености и содержания кислорода. |
На изменение пределов толерантности, обычно называемое повышением сопротивляемости, сильно влияют длительность воздействия, степень изменения факторов среды и прошлая жизнь особи. Физиологические механизмы повышения сопротивляемости требуют определенного времени для приспособления к той или иной ситуации. Внезапное изменение среды может вызвать смерть, но если такое же изменение развивается постепенно, животное способно выжить. Повышение сопротивляемости включает процессы от очень быстрой регуляции до медленной акклиматизации. Таким образом, очень постепенные изменения среды позволяют особи приспособиться к новым условиям. Некоторые виды способны менять пределы своей толерантности в процессе акклиматизации. Так, например, мелкая древесная ящерица Urosaurus ornatus обычно выдерживает температуру до 43,1°С. После содержания этих животных в лаборатории в течение семи - девяти дней при температуре 35°С вместо более привычной для них температуры 22-26°С оказалось, что средняя летальная температура повысилась до 44.5° С.
2. Акклиматизация
Акклиматизация - это форма физиологической адаптации, которая позволяет животному изменить свою толерантность к факторам среды. Обычно термин акклимация применяют к экспериментам, в которых адаптация происходит по отношению к одному фактору, например к температуре. Термин акклиматизация обозначает комплекс адаптивных процессов, протекающих в естественных условиях.
Акклиматизация часто происходит в ответ на сезонные изменения климата. Так, сезонные изменения верхней летальной температуры у пресноводных рыб часто непосредственно коррелируют с изменениями температуры среды обитания. Поскольку поведенческие приспособления могут сделать акклиматизацию ненужной и наоборот, животные в природе используют самые разнообразные комбинации физиологических и поведенческих механизмов.
Температурные предпочтения у рыб часто связаны с уровнем их акклиматизации. У многих видов рыб, по-видимому, высокоразвита поведенческая терморегуляция, и они отвечают на температурный градиент выбором воды с определенной температурой. Золотых рыбок можно научить поддерживать температуру воды в аквариуме, приводя в действие клапан, который впускает холодную воду при повышении температуры. Эти рыбы с достаточной точностью сохраняют температуру аквариума близкой к 34°С.
Феномен выбора температуры в значительной мере объясняет распределение рыб в природе; обычно они предпочитают температуру, к которой акклиматизированы. Такое приспособление имеет биологический смысл. Если бы медленные изменения физиологического состояния, обусловленные акклиматизацией, не сопровождались соответствующими изменениями в поведенческих предпочтениях, то процессам акклиматизации могли бы противостоять механизмы поведения. Так, например, акклиматизации к холоду могла бы противодействовать тенденция к выбору более теплого климата. Если бы кратковременная возможность выбора теплой среды использовалась полностью, то значительная часть работы по акклиматизации к холоду пропадала бы впустую. Очевидная альтернатива для животных состоит в предпочтении условий, к которым они акклиматизированы. Однако картина усложняется феноменом предваряющего приспособления, которым животное отвечает на некоторые свойства среды или на свои собственные биологические часы, подготавливая себя к климатическим изменениям.
Акклиматизацию, как правило, считают относительно медленным процессом по сравнению с быстрыми физиологическими приспособлениями животных в ответ на внезапные изменения внешней среды. Однако обычно существует целый спектр адаптивных процессов от быстрых физиологических реакций до медленной акклиматизации.
|
| Рис.5. Акклиматизация к высоте. Адаптивные изменения у человека, дышавшего четыре дня разреженным воздухом, а затем находившегося шесть дней на уровне моря. В - легочная вентиляция; Э - эритропоэтин сыворотки; Г - скорость синтеза гемоглобина; ЭК - уровень эритроцитов в крови. |
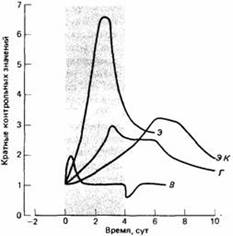
Так, если человека резко поднять на большую высоту над уровнем моря, то из-за разреженного воздуха содержание кислорода у него в крови снизится. Этому снижению вначале противодействует учащенное дыхание, но такая физиологическая реакция требует большой затраты энергии.
Этот быстрый физиологический ответ затем сменяется менее "дорогостоящими", хотя и более медленными формами физиологической адаптации, а те в свою очередь - более долговременными формами акклиматизации, например выработкой большего числа эритроцитов. Но и это требует затрат, поскольку для создания дополнительных клеток нужна энергия, а их присутствие в крови усиливает ее вязкость и работу, которую должно производить сердце, разгоняя кровь по всему телу. Когда человек возвращается на малую высоту, все адаптивные процессы протекают в обратном порядке. Такая обратимость характерна для физиологической адаптации и отличает ее от генетической.
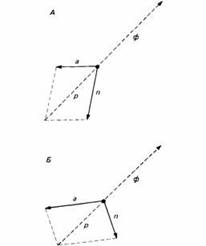
Как видно на рис.5, акклиматизация вместе с регуляцией образуют спектр адаптивных процессов - от быстрых гомеостатических реакций, восстанавливающих физиологическое равновесие в течение суток, до акклиматизации, при которой для достижения физиологически устойчивого состояния могут потребоваться дни или недели. Процессы акклиматизации и регуляции комплементарны. Например, при акклиматизации к высоте требуемое вначале учащенное дыхание замедляется по мере того, как наступает истинная физиологическая акклиматизация. Это значит, что воздействия регуляции и акклиматизации на физиологическое состояние суммируются как векторы, и именно их результирующая приводит к желаемому состоянию, причем вклад каждого вектора меняется с течением времени. Когда добавляется поведенческое приспособление, результат выражается векторной суммой трех процессов.
Когда процессы, представленные векторами-слагаемыми, протекают с очень разными скоростями, как показано на Рис.6, развитие медленных процессов можно рассматривать как "цель" более быстрых. Состояние акклиматизации при этом можно считать оптимальной точкой физиологической регуляции. То же самое верно по отношению к поведению. Так, например, если человек попал из холодного климата в жаркий, он располагает разными альтернативными возможностями приспособления. Он может сохранить свое нормальное поведение, подвергая себя действию солнца, полагаясь на потоотделение и другие физиологические реакции для сохранения нормальной температуры тела. Через несколько недель человек акклиматизируется в этих условиях и будет меньше потеть. Он может также изменить привычное поведение и искать тень, в меньшей степени полагаясь на физиологические механизмы, в частности потоотделение. При этом акклиматизация к новым условиям займет больше времени. Таким образом, как часто бывает, физиологическое и поведенческое решения непосредственной задачи альтернативны. Их эффекты дополняют друг друга и поэтому могут быть представлены векторной суммой.
Учитывая сказанное выше, можно ожидать, что акклиматизация изменяет цель поведения. Некоторые данные говорят о том, что такое изменение действительно происходит. Например, золотистый хомячок начинает готовиться к зимней спячке, когда окружающая температура падает ниже 15°С. Такая
подготовка включает ряд физиологических изменений, обеспечивающих акклиматизацию к холоду, и делает зимнюю спячку физиологически возможной при прочих благоприятных условиях, т.е. наличии материала для постройки гнезда и достаточного количества пищи для создания зимних запасов. Лабораторные опыты показывают, что у хомячков в период акклиматизации, предшествующий зимней спячке, развивается заметное предпочтение к низким температурам среды. Из трех вариантов: 8.19 и 24°С - они выбирают первый. При пробуждении от зимней спячки возникает обратная ситуация, и хомячки активно стремятся в более теплые условия. Как было показано на примере рыб, связь поведенческих, в частности температурных, предпочтений с акклиматизацией имеет биологический смысл.
Акклиматизационные изменения могут явиться прямой реакцией на изменения среды, однако они испытывают также влияние цирканнуальных ритмов. Так, морской полип Campanularia flexuosa обладает годичным циклом роста, развития и отмирания, который сохраняется и при постоянных лабораторных условиях. Известно, что у млекопитающих, в том числе и у человека, происходят сезонные изменения разных метаболических процессов, связанные, как полагают, с эндогенными цирканнуальными ритмами. Такие ритмы могли бы дать возможность животному адаптироваться физиологически еще до сезонных изменений среды.
3. Биологические часы
Животные обычно имеют дело с изменениями среды циклической природы - суточными, приливно-отливными, сезонными. Многие из них обладают некоторым внутренним ритмом, или часами, для предсказания периодических изменений и подготовки к ним.
Существуют три основных способа синхронизовать физиологию и поведение с циклическими изменениями среды:
1) прямой ответ на разные изменения во внешних геофизических стимулах;
|
| Рис.6. Векторно-аддитивные процессы при физиологической регуляции. Вектор акклиматизации а суммируясь с вектором регуляции п, дает результирующую р, которая препятствует сдвигу физиологического состояния ф. В образование результирующей может вносить большом вклад п и малый а, как на рис. А, или малый вклад п и большой - а, как на рис.Б. |
2) внутренний ритм, синхронизирующий поведение животного с периодическими экзогенными изменениями - особенно с суточными или годичными;
3) механизмы синхронизации могут представлять собой сочетание обоих названных способов.
Животное может использовать многие свойства внешней среды для получения информации о течении времени. Движение солнца, луны и звезд, видимое наземному наблюдателю, дает сведения о времени суток, времени года и т.п. Известно, что многие животные пользуются информацией такого типа. Так, медоносные пчелы, живущие в Бразилии, используют солнце как компас при фуражировке. Их можно научить отправляться за пищей по определенному азимуту. Когда таких пчел перевозят из одной местности в другую, они продолжают искать пищу в том же направлении независимо от времени дня. Таким образом, пчелы, родившиеся в Бразилии, способны делать поправку на движение солнца против часовой стрелки. Но пчелы Северного полушария, перемещенные в Бразилию, вначале к этому не способны. Дело в том, что в Северном полушарии солнце представляется движущимся по часовой стрелке, и пчелы должны сначала приспособиться к измененным условиям Бразилии.
Такие же способности известны у рыб и птиц. Имеются также данные, что некоторые животные реагируют на движение луны и звезд. Кроме того, возможно, что животные получают некоторые сведения о времени от таких факторов, как изменение температуры, барометрическое давление и явления магнетизма.
Циркадианными называют эндогенные ритмы, которые обычно короче суток, а цирканнуалъными - эндогенные ритмы с периодом, как правило, менее 365 дней. Многие животные сохраняют ритмическую активность и при изоляции в лаборатории, что говорит о наличии у них эндогенных часов. Однако при этом не исключено, что они реагируют на какой-то экзогенный фактор, еще не обнаруженный экспериментатором, и для проверки того, являются ли часы истинно эндогенными, нужны подходящие критерии. Здесь возможны разные пути. Во-первых, частота ритма может не точно совпадать с каким-либо известным периодическим фактором среды - освещением, температурой или иной геофизической переменной. Во-вторых, в постоянных лабораторных условиях период эндогенного ритма обычно отклоняется от наблюдаемого в естественных условиях. В-третьих, ритм может сохраниться, когда животное перемещают из одной части света в другую. Только при соответствии такого рода критериям можно говорить об эндогенности того или иного ритма.
Поскольку эндогенные ритмы имеют тенденцию постепенно отклоняться от экзогенного цикла, организм должен обладать способностью синхронизовать свой эндогенный ритм с периодическими внешними явлениями. Ашофф ввел термин Zeitgeber для агента среды, который согласует поведение организма с внешними ритмами. Так, например, когда ящериц Lacerta sicula выводят в инкубаторе при температурном и световом режимах, соответствующих 16 - или 36-часовым "суткам", они развиваются нормально и имеют нормальные циркадианные ритмы активности при проверке в постоянных лабораторных условиях. Следовательно, циркадианный ритм этих животных эндогенный и не зависит от индивидуального опыта жизни при том или ином цикле смены дня и ночи. Температурный цикл с 24-часовым периодом и амплитудой 0,6°С - достаточно эффективный времязадатель, захватывающий ритм активности ящериц.
В целом создается мнение, что у многих животных, от одноклеточных до сложно устроенных многоклеточных, в ходе эволюции развилось чувство времени, основанное на действии эндогенных часов, захватываемых экзогенными ритмами.
4. Репродуктивное поведение н физиология
Сезонные климатические изменения оказывают мощное влияние на успех размножения многих видов. У птиц наличие корма для птенцов, по-видимому, главный определяющий фактор такого успеха. В средних и северных широтах размножение птиц связано с временем года; обычно они откладывают яйца весной, что дает возможность птенцам достаточно созреть, чтобы противостоять зимним условиям или выдержать долгий миграционный перелет. Так, например, песочники, размножающиеся в арктических областях, строят гнезда и высиживают птенцов весной, когда земля еще покрыта снегом. Обычно птенцы вылупляются, когда снег тает и в изобилии появляются насекомые, которые служат для них кормом.
Репродуктивная физиология у сезонно размножающихся животных привязана к годичному циклу изменений среды таким образом, что появление детенышей предваряет пик изобилия корма или неблагоприятные климатические условия. Это осуществляется двумя способами. Во-первых, изменения внешней температуры, продолжительности светового дня или других факторов среды вызывают физиологические изменения в определенное время года. Во-вторых, сезонные физиологические изменения запрограммированы посредством эндогенных цирканнуальных часов.
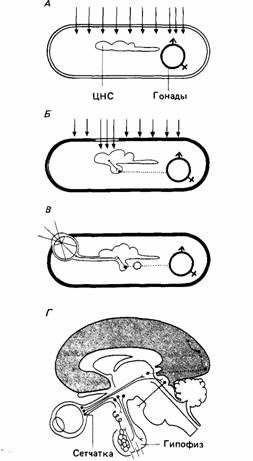
Самое регулярное и предсказуемое изменение среды связано с продолжительностью светового дня. У прозрачных организмов свет может прямо действовать на гонады, приводя их в репродуктивное состояние в надлежащее время. Некоторые другие животные имеют прозрачное "окно", которое пропускает свет в головной мозг. Нейросекреторные клетки превращают световые стимулы в химические сигналы. У ряда млекопитающих эпифиз, расположенный на дорсальной поверхности головного мозга, может действовать как преобразователь света. Но свет влияет на размножение млекопитающих, главным образом действуя через сетчатку на гипоталамус, как показывает схема Шаррера.
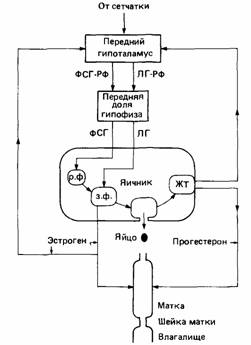
Регуляция физиологии размножения состоит в сложном взаимодействии ряда гормонов. У большинства позвоночных факторы среды стимулируют выработку гонадотропных гормонов гипофизом. Эти гормоны стимулируют рост и активность семенников и яичников, которые в свою очередь производят характерные половые гормоны. К концу сезона размножения активность гипофиза снижается, гонады также становятся неактивными и репродуктивное поведение затухает. Кроме сезонных циклов половой активности у многих млекопитающих имеется гораздо более короткий цикл - эструс, или "течка".
|
| Рис.7. Влияние света на гонады по Шарреру А. У прозрачных животных свет может оказывать прямое действие на внутренние органы. Б. Некоторые непрозрачные животные обладают прозрачным "окном", пропускающим свет к фоточувствительным областям головного мозга, который затем стимулирует гонады с помощью гормонов, выделяемых гипофизом. В. Система, в которой свет действует на сетчатку, посылающую нервные сигналы гипоталамусу. Эта область мозга стимулирует выделение гипофизом гонадотронных гормонов. Г. Система В применительно к головному мозгу человека. |
|
| Рис.8. Главные гормональные пути, участвующие в работе яичника у млекопитающего. ФСГ-РФ - фактор, стимулирующий высвобождение фолликулостимулирующего гормона; ЛГ-РФ фактор, высвобождающий лютеинизирующий гормон; р. ф. - растущий фолликул; з. ф. - зрелый фолликул; ЖТ - желтое тело. |
У одних животных, например у рыжей лисицы, течка бывает только раз в году; у других, например у домашних собак, - два раза; у третьих - чаще. У птиц число выводков в одном сезоне может зависеть от количества корма.
На характер размножения многих видов влияют как фотопериодические факторы, так и эндогенные цирканнуальные часы. Предполагается, что эти два механизма могут взаимодействовать. На виды, обитающие в высоких широтах, обычно сильнее всего влияет фотопериодичность. В низких широтах годичных фотопериодических изменений меньше, но тем не менее годичный репродуктивный цикл может иметь свои преимущества. Например, у животных, обитающих в безводной пустыне, начало процесса размножения может зависеть от выпадения дождя. Некоторые экваториальные виды обладают выраженным ритмом размножения, который, по-видимому, не связан с сезонными изменениями. Ряд морских птиц, в том числе коричневая олуша, темная крачка и тонкоклювая крачка, размножаются каждые 8-10 месяцев. Постепенное расхождение между такими циклами размножения и годичным циклом говорит о том, что в экваториальных условиях ни одно время года не является предпочтительным. Почему же в таком случае птицы не размножаются непрерывно? Возможно, это потребовало бы слишком много энергии, и оптимальным вариантом становится периодический отдых, во время которого может происходить линька.
Годичный цикл размножения сходен с акклиматизацией тем, что при нем также происходят медленные физиологические процессы, коренным образом меняющие физиологическое состояние животного. Репродуктивная деятельность, включающая в себя защиту территории, ухаживание, спаривание и заботу о потомстве, требует от животного дополнительной энергии и физиологических затрат, которые должен обслуживать весь его организм. Если в данный год эти затраты так велики, что животное не в состоянии сохранить физиологическую стабильность, оно вынуждено отказаться от размножения до более благоприятного года.
5. Зимняя спячка
В частях света со сменой времен года животным иногда приходится приспосабливаться к длительным периодам неблагоприятной погоды. Некоторые из них избегают таких условий путем миграции, а другие способны пережить их, впадая в продолжительный сон, который при высоких температурах называется эстивацией, а при низких - гибернацией. Некоторые пустынные грызуны, например суслики, летом впадают в оцепенение, при котором температура тела падает и происходит общее снижение физиологической активности. Это значит, что затраты энергии уменьшаются и животное может долго жить, не питаясь. Потери воды сокращаются вследствие меньшего потребления пищи, чему способствует уход в нору. Некоторые грызуны запасают корм в своих норах и сохраняют в них высокую влажность, закупоривая вход. Летняя спячка наиболее распространена среди обитателей пустыни, но не ограничивается ими. Так, например, многие европейские виды дождевых червей летом спят. Каждое животное вырывает небольшую нору глубоко в земле и сворачивается в ней в клубок. Наступление такой летней спячки связано с низкой влажностью, и ее можно предотвратить содержанием червей во влажной атмосфере.
В зимнюю спячку впадают многие виды в северных широтах, что позволяет им избегать зимних условий, требующих очень больших энергетических затрат. Подлинная зимняя спячка отличается от частичной спячки, в которую впадают европейский бурый и американский черный медведи. При частичной спячке температура тела медведя может упасть примерно от 38° до 30°С, хотя температуры тела ниже 15°С детальны. При истинной зимней спячке температура тела может упасть даже до 2°С. Истинная зимняя спячка характерна для мелких млекопитающих, хотя сходные с ней виды оцепенения бывают у енотов, барсуков и некоторых птиц. У многих видов колибри во время оцепенения температура тела снижается до температуры среды, хотя температуры ниже 8°С детальны. У птиц оцепенение длится обычно всего несколько часов, а сезонная спячка известна только в одном их семействе - у козодоев.
Период спячки, называемый диапаузой, как реакция на неблагоприятные климатические условия бывает также у насекомых. Диапауза часто связана с определенной стадией жизненного цикла. Насекомые, избегающие замерзания с помощью переохлаждения, могут пережить сильный холод в диапаузе, при которой температура замерзания жидкостей тела намного ниже 0°С. Глубина переохлаждения может зависеть от степени акклиматизации, которая постепенно меняет химический состав жидкостей тела. Канадская оса Bracon cephi может снизить точку их замерзания до - 46° С, повысив концентрацию глицерола в гемолимфе.
Истинная зимняя спячка наступает только у мелких млекопитающих, которые охлаждаются быстрее крупных из-за относительно большей поверхности тела. Они также быстрее согреваются благодаря своей малой теплоемкости. Зимняя спячка характеризуется сноподобным состоянием с замедленными дыханием и сердцебиением. При этом животные часто выбирают особое место и принимают позу, в которой обычно спят. При зимней спячке температура тела и затраты энергии падают ниже уровня, характерного для нормального сна. У таких животных часто откладывается специальный бурый жир. Его главная функция состоит в образовании тепла, а не энергии для обменных процессов, для которых мобилизуются резервы обыкновенного жира. Особенно важную роль бурый жир играет в период пробуждения от спячки, когда температура тела должна быстро повыситься.
Некоторые млекопитающие, в том числе золотистый суслик и североамериканский лесной сурок, обладают выраженным цирканнуальным ритмом, лежащим в основе их сезонной зимней спячки. Названные суслики встречаются в западной части Северной Америки на высотах от 1500 до 3600 м над уровнем моря от севера Британской Колумбии до юга Калифорнии. Обычно их зимняя спячка длится три-четыре месяца, и за это время масса их тела значительно снижается. После спячки у них быстро растет потребление пищи и вес тела достигает отмечавшегося перед началом спячки в октябре. При изоляции в лабораторных условиях ритм зимней спячки и связанных с ней изменений в массе тела может сохраняться ряд лет даже при постоянных освещении и температуре. Если сыворотку крови от суслика в состоянии зимней спячки ввести неспящему животному, то последний также впадает в спячку.
| Рис.9. Цирканнуальные ритмы изменения массы тела и периоды зимней спячки двух особей Citellus lateralis, содержавшихся в неизменных лабораторных условиях. |
|
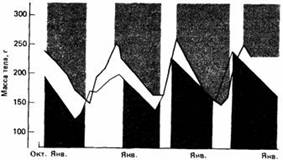
Этот факт служит веским доводом в пользу того, что основной физиологический механизм гибернации управляется эндогенными биологическими часами, захватываемыми ритмом внешних событий с помощью особого времязадателя. Это ясно показано на североамериканском лесном сурке, у которого также известен цирканнуальный ритм зимней спячки. Эти сурки были переправлены из восточной части США в Австралию - в новые для них условия освещения и температуры. Сурки за два года изменили нормальный ритм гибернации на обратный, приведя его в соответствие с местными условиями, несмотря на то, что в изобилии получали корм и воду и не страдали от понижения температуры.
Зимняя спячка похожа на акклиматизацию тем, что представляет собой длительное изменение физиологического состояния, позволяющее животному справляться с сезонными изменениями внешних условий. Цирканнуальные изменения физиологического состояния животного могут быть представлены в физиологическом пространстве. Годичные циклы размножения и зимней спячки являются в значительной степени врожденными, хотя фактически соответствуют общим сезонным изменениям в природе.
6. Миграция
Миграция из одного местообитания в другое на периодической или сезонной основе происходит у многих видов животных, в том числе у дневных бабочек, саранчи, лососей, птиц, летучих мышей и антилоп. Это не единственная форма миграции, поскольку у некоторых видов миграция составляет часть обследования и колонизации новых территорий. Так, популяционный взрыв у норвежских леммингов ведет к расселительной миграции, которая может охватить тысячи особей, обычно неполовозрелых самцов. Потоки леммингов устремляются вниз по склонам гор в долины. Многие тонут, пытаясь переплыть широкие водные пространства. Лемминги - хорошие пловцы и обычно не входят в воду, если не видят противоположного берега. Но порой давление множества скопившихся на берегу особей друг на друга таково, что некоторые из них вынуждены выплывать в открытое море.
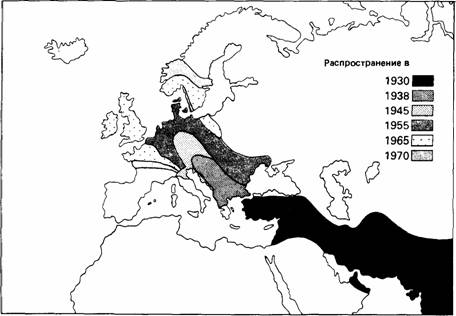
Заселение новых областей иногда достигается за счет постепенной миграции нескольких поколений, как, например, у кольчатой горлицы. Исследовательская миграция часто встречается у молодых позвоночных животных и при благоприятных условиях может обеспечить успешное расселение.
|
|
| Рис.10. Карта расширения ареала кольчатой горлицы в XX столетии. |
|
|
| Рис.11. Цирканнуальные ритмы изменения длины семенников у садовых славок, содержавшихся в течение трех лет в неизменных фотопериодических условиях. СТ соотношение света и темноты. |
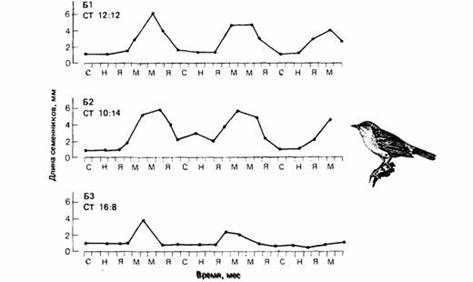
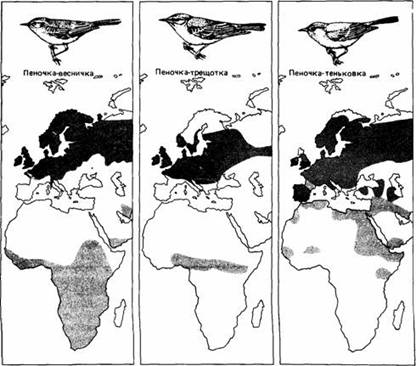
Рис.12. Места гнездовий, зимовок и их перекрывания у некоторых мигрирующих пеночек.
Периодические миграции возникают у многих видов в ответ на изменения внешних условий. Так, например, саранча Schistocerca gregaria заселяет в определенное время года безводные области.
В зависимости от степени скопления эти насекомые развиваются в одну из трех имагинальных форм. Относящиеся к стадной форме особи собираются в плотные стаи, которые переселяются по ветру в области с низким барометрическим давлением, где скорее всего может идти дождь. Они летят днем и останавливаются, попав во влажные условия. Здесь происходят половое созревание, спаривание и откладка яиц. Саранчевые стаи мигрируют по замкнутым сезонным маршрутам, но время генерации слишком коротко для того, чтобы отдельные особи могли завершить полный круг.
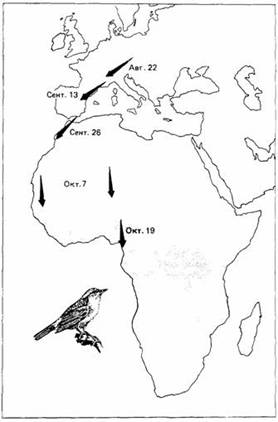
Некоторые сезонные миграции в отличие от периодических начинаются на основе цирканнуального ритма, а не в ответ на изменения внешней среды. У певчих птиц из родов Phylloscopus и Sylvia наблюдали разное миграционное поведение. У таких типичных мигрантов на дальние расстояния, как садовая славка, горная славка и пеночка-весничка, происходят значительные сезонные изменения в массе тела, характере линьки, размере семенников, ночном беспокойстве и пищевых предпочтениях. Европейские популяции этих видов зимуют в Африке и мигрируют через Сахару. Если же они помещены в постоянные лабораторные условия через несколько дней после вылупления, то процессы, которые у свободноживущих птиц являются сезонными, сохраняют сезонность у особей тех же видов в неволе, хотя период колебаний этих процессов у них несколько короче календарного года.
Мигранты на средние расстояния, такие, как славка-черноголовка и пеночка-теньковка, зимуют в Европе и Африке, причем масса тела и другие миграционные показатели у них меняются умеренно. В постоянных лабораторных условиях у этих видов тоже происходят сезонные изменения. Белоусая славка и провансальская славка являются частичными мигрантами, которые зимуют в средиземноморской области гнездования, a S. sarda balearica проводит весь год и эндемична на Балеарских и Питиузских островах в Средиземном море. У этих видов масса тела достаточно постоянна в течение всего года, а линька оперения на туловище продолжается долго и перемежается периодами ночного беспокойства. В лаборатории у них можно наблюдать некоторые проявления сезонных изменений, но с выраженными индивидуальными различиями.
Вероятно, у видов этих двух родов происходят сезонные изменения миграционных показателей, интенсивность проявления которых коррелирует с миграционным поведением вида, а распределение во времени говорит о том, что начало миграции, возможно, частично регулируется эндогенным цирканнуальным ритмом. Подобные же сезонные ритмы установлены у некоторых видов птиц для таких физиологических процессов, как линька, отложение жира, миграционное беспокойство и размножение.
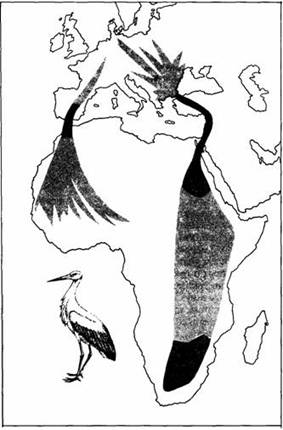
Получены также данные, что эндогенные факторы управляют не только началом миграционной активности, но и ее характером. У многих видов пути миграции характерны для определенных географических популяций. Так, например, белые аисты, гнездящиеся в Западной Европе, летят на места зимовок в Африку западным путем через Испанию и Гибралтар, а гнездящиеся в Восточной Европе избирают восточный путь, как показано на рис.13. Неопытные молодые аисты, выращенные в неволе в Восточной Европе, но выпущенные на свободу в Западной, летят в юго-восточном направлении, характерном для птиц из Восточной Европы. Такие же опыты с другими видами пернатых свидетельствуют о генетически обусловленном выборе направления миграции.
Чтобы достичь места зимовки, птица должна лететь не только в правильном направлении, но и на вполне определенное расстояние. Имеются данные о расстояниях, которые преодолевают при первом перелете молодые славки, не сопровождаемые взрослыми птицами. Эти данные говорят о том, что у птиц существуют эндогенные часы, указывающие, сколько времени нужно лететь на каждом этапе перелета. Наблюдается хорошая корреляция между числом часов миграционного беспокойства птиц в неволе и расстоянием, нормально пролетаемым ими во время миграции. Расстояние, эквивалентное одному часу такого беспокойства, может быть определено путем сравнения поведения птиц одной и той же популяции в неволе и во время перелета. Зная скорость полета мигрантов, можно вычислить расстояние, которое покрыли бы сидящие в клетке птицы, если бы летели в правильном направлении. Расчеты привели к результатам, близким к полученным при наблюдении над свободно мигрирующими птицами этой же популяции. Садовые славки, содержавшиеся в неволе во время первой осенней миграции, поворачивались на юго-запад в августе и сентябре и на юго-восток в октябре, ноябре и декабре. Эти данные совпадают с "расписанием" естественной свободной миграции, показанным на рис.14. Некоторые птицы перелетают много сотен миль над океаном, где ориентирами служат лишь несколько островков. Так, например, полярные крачки размножаются в Арктике, а осенью мигрируют к антарктическим паковым льдам. Весной они совершают обратный перелет, хотя, вероятно, другим путем. Известно, что некоторые особи пролетают 9000 миль. Подобным же образом тонкоклювый буревестник размножается в юго-восточной Австралии и мигрирует на Аляску через Японию. Обратное путешествие совершается вдоль западного берега Северной Америки.
7. Лунные и приливно-отливные ритмы
| Рис.13. Миграционные пути восточно- и западноевропейских популяций белых аистов. Интенсивность затенения - обилие птиц. |
|
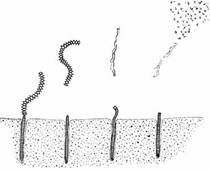
Как видно с земли, луна движется по такому же пути, как и солнце, и с каждым днем восходит на 50 мин позднее, из-за чего иногда видна среди дня, а иногда только ночью. Известно, что лунный цикл, равный 29,5 сут, влияет на самые разные стороны поведения животного. У тихоокеанского червя палоло репродуктивная активность приурочена только к квадратурным приливам последней четверти луны в октябре и ноябре. Как показано на рис.16, задний конец червя с генитальными органами отделяется от передней части и выплывает на поверхность, где в воду выбрасываются яйцеклетки и сперматозоиды. Эти черви дают обильную пищу акулам и другим рыбам, но, синхронизируя репродуктивную активность, добиваются того, что часть гамет всегда может выжить. В лабораторных условиях черви образуют гаметы в этот же срок, т.е. их репродуктивное поведение, вероятно, контролируется эндогенными часами.
|
| Рис.14. Предпочтения того или иного направления, наблюдавшиеся в лабораторных опытах по миграционному беспокойству у садовых славок. Направление стрелок предпочтение, наблюдавшееся в указанные даты. Положение стрелок на карте места, которых перелетные славки достигали бы в указанные даты при миграции к местам зимовки в южной Африке. |
Лунные ритмы известны и для наземных животных. Например, летучие мыши крыланы на Ямайке распределяют время своего кормления так, что в новолуние покидают дневные укрытия вечером и питаются в течение всей темной ночи. А в полнолуние они вылетают из дневных укрытий в это же время, но возвращаются туда, когда луна поднимается высоко, даже если ее заволакивают тучи. Это говорит о том, что для распределения фуражировки во времени они пользуются эндогенными лунными часами.
Приливы и отливы происходят от изменений совместного притяжения Солнца и Луны. Приливно-отливный цикл повторяется дважды в течение одного лунного месяца. У многих морских животных ритмы поведения совпадают с этим циклом и, как установлено, управляются эндогенными часами. Так, например, суточный ритм активности прибрежного краба Carcinus maenas накладывается на ритм приливов и отливов: активность этого животного синхронизирована с высокой водой. Такой ритм сохраняется в постоянных лабораторных условиях около недели, а затем исчезает, но может быть восстановлен приблизительно 6-часовым охлаждением краба до температуры, близкой к точке замерзания.
| Рис.15. Места размножения и миграционные пути тонкоклювых буревестников, установленные по пойманным окольцованным птицам. |
|

Этот холодовой шок, по-видимому, вновь запускает "приливные" часы, может быть, потому, что он сходен с воздействием прилива, случайно достигающего выброшенного на берег краба. В одном опыте таких крабов выращивали в лаборатории от яйца до взрослого состояния в режиме день-ночь без влияний приливов. У этих крабов появлялся только суточный ритм активности. Однако после одного холодового шока возник приливно-отливный ритм. По-видимому, эндогенные "приливные" часы не действовали, пока холодовой шок не запустил их.
Подобные явления наблюдаются и у других крабов. У манящих крабов известны приливно-отливные ритмы активности, сохраняющиеся до пяти недель в постоянных лабораторных условиях. Крабы выползают из нор при отливе и активно кормятся, ухаживают и т.п. При наступлении прилива они возвращаются в свои норы. У зеленых крабов ритм противоположный, они активизируются во время прилива. У этих и многих других видов крабов падение температуры, вызванное приливными водами, действует как времязадатель, устанавливающий фазу ритма активности.
|
| Рис. Размножение червя палоло. |
|
|
| Рис.17. Изменения высоты прилива в Ла-Холья в Калифорнии в сравнении с плавательной активностью бокоплава Synchelidium в лабораторных условиях, |
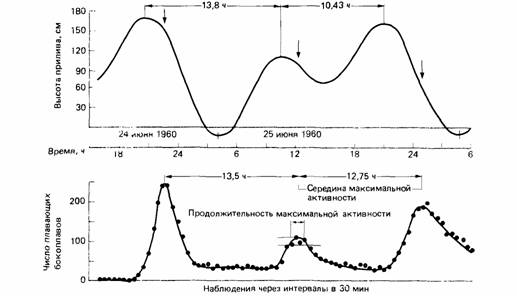
Наложение друг на друга циркадианного и приливно-отливного ритмов позволяет животному приспособиться к нерегулярным изменениям воды, происходящим в некоторых частях света. Например, на побережье Калифорнии за периодом 13.8 ч соответствующим сильному отливу, следует период 10.43 ч с небольшим отливом. У литорального ракообразного Synchelidium плавательная активность здесь почти точно следует за этим ритмом. Такой плавательный ритм сохраняется в течение нескольких дней в постоянных лабораторных условиях.
8. Циркадианные ритмы и суточное распределение активности
Большинство животных сталкивается с различными условиями днем и ночью. Такие изменения действуют на них как прямо, так и косвенно. Например, под влиянием колебаний температуры, освещенности и т.п. могут меняться доступность пищи и число хищников.
Приспосабливаясь к различиям условий днем и ночью, животное производит суточное распределение активности, представленной многими видами поведения, которые в совокупности образуют распорядок, повторяющийся с небольшими отклонениями изо дня в день. Можно заметить, что каждый вид - активности имеет тенденцию следовать типичному суточному ритму. "Режим дня" в целом у животных изучен сравнительно мало, но суточные ритмы привлекли к себе значительное внимание специалистов по циркадианным часам.
Самые важные суточные изменения во внешней среде - это изменения освещенности и температуры. Животные, специализированные к дневному зрению, ночью могут оказаться в неблагоприятном положении, потому что легко станут жертвой хищников и не смогут эффективно добывать себе пищу. В холодном климате мелким млекопитающим может быть выгодна ночная активность при низкой температуре. Период наибольшей теплопродукции придется у них в таком случае на самую холодную часть 24-часового цикла, когда активность способствует терморегуляции.
|
|
| Рис.18. Частота прятания и извлечения спрятанной добычи пустельгой. |
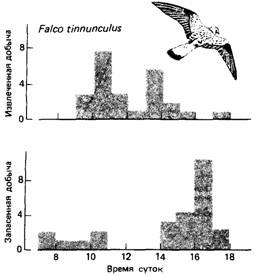
В то же время мелкие птицы холодными ночами сберегают энергию, становясь неактивными и давая температуре тела снизиться. В жарком климате мелким млекопитающим выгодно быть ночными животными, избегая дневной жары.
Неудивительно, что ритмы покоя и активности широко распространены в животном мире. Если ночная активность невыгодна, лучше всего сидеть неподвижно в надежном месте и сохранять как можно больше энергии. Было высказано предположение, что это одна из главных функций сна. Ночные виды могут затаиваться днем, если им грозит нападение. Если же они сами являются ночными хищниками, то днем могут прятаться и бездействовать, чтобы не спугнуть добычу. Таким образом, суточные ритмы физической внешней среды делают некоторые виды активности выгодными в одно время и невыгодными в другое. Многое зависит от общей экологии данного вида.
|
|
| Рис. 19. Передвижения пустельги, наблюдавшееся в Нидерландах 12, 13, 14 и 16 августа 1977 г. Цифры на маршрутах полета - время суток. Суша обозначена темным, море - светлым. |
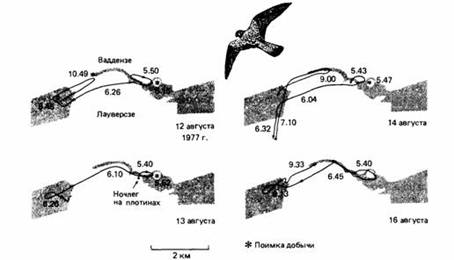
Животное, адаптированное к среде, придерживается такого суточного распорядка, который максимально увеличивает ценность разных видов его активности для выживания. Отчасти это означает наилучшее использование предоставляющихся возможностей. Например, распространенная в Европе пустельга, дневной хищник, нападает главным образом на мелких млекопитающих. Эта птица обнаруживает добычу с помощью зрения и успешнее всего охотится при хорошем освещении. В полевых исследованиях установлено, что пустельга хватает добычу весь день, но не всегда тут же поедает ее. Она прячет избыток пищи в случайно выбранных местах своего охотничьего участка. Это происходит в течение всего дня. Спрятанная пища обычно извлекается в сумерках, что позволяет пустельге максимально использовать для охоты светлую часть суток, не тратя слишком много времени на еду. Кроме того, если бы птица сразу съедала всю добычу, она становилась бы слишком тяжелой и, вероятно, эффективность ее охоты была бы снижена.
В дополнение к типичному для вида суточному распределению активности отдельные животные могут приобретать свои собственные привычки. Так, пустельги, нашедшие пищу в определенное время и на определенном месте, стремятся повторить на следующий день тот же охотничий маршрут. Такая стратегия годится там, где виды-жертвы тоже имеют свой собственный типичный суточный распорядок.
Похожие работы
... акты всегда отличаются наличием такой ориентировочной основы действия, такой стратегии и тактики. В этом случае А.Р. Лурия обращался к некоторым классическим опытам, в которых изучалось интеллектуальное поведение животных. Эти опыты были проделаны Кёлером, и получили известность как элементарнейшие опыты с употреблением орудий. Употребление орудий есть всегда типичное интеллектуальное действие. ...
... решает задачу, логически рассуждая, делая необходимые выводы, животные находят правильное решение случайно, часто действуя методом проб и ошибок. 1.2 Предпосылки интеллектуального поведения животных Предпосылкой для инстинктивного поведения является отражение отдельных свойств внешней среды, которое действует на механизм, пускающий в ход врожденный инстинктивный акт. Предпосылкой для ...

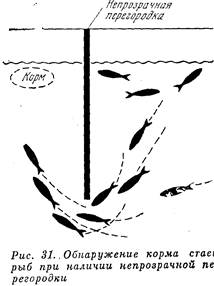
... отличаются обычно высокой численностью и большой экологической устойчивостью. Можно вслед за другими исследователями установить два основных типа несемейных группировок животных, которые различаются по особенностям взаимоотношении и поведения животных, составляющих группу: 1) с доминированием одних особей над другими (такие группы называются иерархическими); 2) стаи без доминирования. В ...

... результаты были получены благодаря широкому использованию в лабораторных тестах генетически охарактеризованных животных, а также применению основных методов анализа генетических различий. В настоящее время исследование когнитивных способностей животных в этом тесте является одним из ведущих подходов в оценке особенностей поведения трансгенных животных и мышей-нокаутов. Детальнее с этими вопросами ...
0 комментариев