Покровы тела
Способность живых существ к жизни в различных условиях является результатом эволюции не только их самих, но и их систем органов. Ниже мы рассмотрим это заключение на примерах эволюции систем органов животных, начиная с простейших и заканчивая млекопитающими.
Внешние покровы животных (кожа или интегумент) выполняют защитную функцию, предохраняют тело от излишней потери воды и помогают регулировать температуру тела. Кроме того, у ряда организмов они являются дополнительным органом, принимающим участие в обмене веществ (дыхании и выделении).
У беспозвоночных покровы тела не достигают значительного развития, будучи представленными эктодермой и ее производными. Эволюция покровов у них шла в направлении развития мерцательного эпителия в плоский эпителий, что отмечается у турбеллярий и других плоских червей, а также у круглых и кольчатых червей. Мерцательный эпителий служит примитивным органом движения, тогда как плоские этой способностью не обладают.
У членистоногих поверхностный слой эпителия превращается в хитинизированную кутикулу, которая развивается в панцирь у ракообразных. У моллюсков поверхностный слой эпителия развивается в раковину, содержащую известь. У животных отдельных групп (кольчатые черви, членистоногие и моллюски) в эпидермисе локализованы одно- или многоклеточные железы (слюнные, паутинные, ядовитые и другие).
У хордовых эволюция кожных покровов шла в направлении замены однослойного эпителия многослойным, в частности, в формировании двух слоев и в превалирующем развитии собственно кожи (кориума).
Кожа хордовых состоит из поверхностного слоя эктодермального происхождения, или эпидермиса, иногда называемого кутикулой, и нижнего слоя мезодермального происхождения — собственно кожи (кориума), или дермы. У туникат (асцидий, пиросом и других морских животных) наружний покров представляет собой студенистую или хрящевую оболочку, являющуюся продуктом эктодермального эпителия и пронизанную мезодермальными клетками и сосудами. Покровы ланцетников имеют двуслойное строение. Первый слой представлен цилиндрическим однослойным эпителием эктодермального происхождения, второй — студенистой соединительной тканью, выполняющей роль кориума.
Покровы у позвоночных достигли большого развития и представляют собой кожу, которая построена из двух различных слоев. Один из них является поверхностным и состоит из многослойного эпителиального эпидермиса эктодермального происхождения, второй — нижним, собственно кожей, которая прилегает к подкожной ткани. Последняя, в свою очередь, прилегает к мышцам и костям.
Поскольку у земноводных кожа используется в качестве дополнительного органа дыхания, но при этом необходима водная пленка, то это привело к развитию в коже этих животных многослойных желез, продуцирующих слизь и увлажняющих ее. Кожа земноводных содержит также ядовитые железы, выполняющие защитную функцию.
У млекопитающих развились производные кожи (поверхностный каратин, волосы, копыта и рога), которые выполняют важные функции в жизни организмов. Большое значение в терморегуляции имеют кожные потовые железы.
СкелетСкелет выполняет опорную и защитную функции. У беспозвоночных скелетом являются ригидные структуры, имеющие самый разнообразный характер и подразделяемые на эндоскелет и экзоскелет
Эндоскелет построен из органических фибрилл или кристаллов неорганических соединений. У простейших эндоскелетной структурой является аксостиль ряда жгутиковых, который имеет фиб-риллярную структуру, у радиолярий и фероминифер эндоскелет представлен кристаллами неорганического происхождения, у губок — спикулами (иглами, состоящими из отложений карбоната кальция или силикона вокруг клеточных фибрилл), у морских звезд — различными хрящеподобными тканями. Некоторые членистоногие имеют хорошо развитый эндоскелет, к которому прикрепляются мышцы. Известковые пластинки и иглы иг-локожих имеют мезодермаль-ное происхождение.
Экзоскелет имеется у многих беспозвоночных организмов и характеризуется чрезвычайным разнообразием, начиная от пелликулы (утолщенной клеточной стенки) и заканчивая такими образованиями, как крылья бабочек. Импрегнация эк-зоскелета неорганическими солями повышает его ригидность. Гибкость экзоскелета, которая необходима для его мобильности, достигается у многих нематод вследствие гибкости кутикулы, а у коловраток и особенно у членистоногих благодаря наличию гибких районов между ригидными участками кутикулы. У членистоногих кутикула является результатом секреции клеточного эпидермиса и состоит из тонкой наружной эпикутикулы и толстой эндокутикулы, в состав которой входят хитин и белок.
У низших хордовых развит осевой скелет, которым является хорда. У бесчерепных эндоскелетом является хорда, а экзоскелетом — стержневидные образования, состоящие из плотного студенистого вещества (непарные плавники и опора жаберного аппарата). У круглоротых хорда также сохраняется на протяжении всего онтогенеза. Однако у них под хордой уже появляются парные закладки позвонков в виде хрящевидных образований (верхние дуги). У беспозвоночных кальциевый скелет представлен кальцитом.
У позвоночных эндоскелет достигает высокого развития, являясь хрящевым и (или) костным скелетом. У этих организмов различают осевой скелет, скелет головы (черепная коробка и висцеральный скелет) и скелет конечностей. Экзоскелет представлен волосами, перьями, чешуей, рогами, отростками рогов, копытами, когтями, косточками в коже.
У примитивных рыб наряду с верхними дугами появляются закладки низших дуг. У высших рыб из тканей, окружающих хорду, и оснований верхних и низших дуг, развиваются позвонки. Позвоночный столб состоит из туловищного и хвостового отделов. Благодаря сращению концов верхних дуг образуется канал для спинного мозга. К низшим дугам прикрепляются ребра.
У земноводных процесс замены хорды позвонками углубляется еще в большей степени и происходит в ранние периоды развития. Развиваются четыре отдела позвоночника — шейный, грудной, крестцовый и хвостовой. Шейный отдел очень короток (один позвонок), грудной отдел насчитывает пять позвонков, к которым прикрепляются свободно заканчивающиеся ребра (не доходящие до грудины), крестцовый отдел также состоит из одного позвонка, к которому прикрепляются кости таза. Хвост хвостатых амфибий состоит из нескольких позвонков, бесхвостых — из одного позвонка, называемого уростилем.
У амниот (рептилий, птиц и млекопитающих) различают осевой скелет, скелет головы (череп) и скелет конечностей. Осевой скелет состоит из шейного, грудного, поясничного, крестцового и хвостового отделов.
Количество шейных позвонков варьирует от 1 (у современных земноводных) до 76 (у ископаемых форм — морских плезиозавров). У мниот и лабиринтодонтов первые два шейных позвонка специализированны, обеспечивая поддержание и движение головы. Количество шейных позвонков зависит от длины шеи. У некоторых рептилий их количество достигает 8, у птиц — до 25. Современные млекопитающие имеют 7 шейных позвонков.
Количество грудных позвонков различно у разных групп позвоночных. У черепах 10 грудных позвонков слиты со связками и костями и формируют панцирь, у пресмыкающихся к грудным позвонкам прикрепляются ребра, что формирует грудную клетку. Птицы имеют по 4—6 грудных позвонков. У млекопитающих чаще развивается 10-20 грудных позвонков. У человека грудную клетку формируют также ребра (12 пар) и грудина.
Поясничный отдел развит по-разному. У рептилий поясничный отдел выражен плохо. У птиц поясничные позвонки обычно слиты с грудными, крестцовыми и урокрестцовыми, формируя сложный крестец. У млекопитающих количество поясничных позвонков составляет от 3 до 24 в зависимости от вида.
Количество хвостовых позвонков зависит от длины хвоста. У лягушек, например, хвостовые позвонки сливаются в одиночную кость (уростиль), у птиц имеется 4-6 хвостовых позвонков, которые сливаются также в уростиль. Некоторые ископаемые птицы (археоптерикс) имели длинный хвост, состоящий из 20 позвонков. У млекопитающих количество позвонков в хвосте зависит от вида. Прямохождение человека вело к искривлению позвоночника, т. е. это оказалось «платой» за Прямохождение
Скелет головы состоит из черепа и висцерального (лицевого) скелета.
Черепная коробка различается не только по форме, но и по количеству входящих в нее костей, которые очень колеблются в зависимости от принадлежности животных к тому или иному классу. У костных рыб, например, количество костей черепа превышает 100, в то время как у земноводных оно равно 15-19, у пресмыкающихся — от 40 до 70, у птиц — 20, у примитивных млекопитающих (Didelphis) — 48, у шимпанзе — 22, у человека — 27 Снижение количества костей черепа в эволюции от рыб до млекопитающих является результатом потери индивидуальных костей или их слияния с другими костями.
Эволюция висцерального скелета шла в направлении его редукции. Он очень уменьшен у четвероногих позвоночных, дышащих легкими.
Первоначальная функция висцеральных дуг заключалась в поддержке жабер. Это подтверждается и наличием жаберных щелей у низших рыб (кроссоптериг, осетровых и др.). У рептилий и птиц первая жаберная дуга развилась в скелет гортани. Жаберная крышка, представлявшая собой костный клапан, покрывающий жабры, и большинство жабер были утеряны наземными позвоночными. Многие кости мозгового черепа, которые разделены у низших четвероногих, слились у млекопитающих. Млекопитающие отличаются от других животных наличием трех небольших костей в полости среднего уха. Одна из этих костей — стремечко — гомологична второй висцеральной дуге рыб, другая — наковальня — гомологична кости нижней челюсти других позвоночных, а третья — молоточек — гомологична кости верхней челюсти других позвоночных.
Скелет конечностей имеет особенности, причем ключевым является вопрос о переходе плавников в конечности. Конечности классифицируют на непарные и парные. Примером непарных конечностей является спинной и хвостовой плавники у рыб, парных — свободные конечности, роль которых выполняют плавники у рыб и пятипалые конечности у наземных животных. Парные плавники кистеперых рыб содержат гомологи плечевой, локтевой и лучевой костей пятипалой конечности наземных позвоночных, что свидетельствует о происхождении пятипалой конечности от плавника кистеперых рыб. Свободные конечности четвероногих обычно состоят из трех сегментов, которые организованы по-разному у разных животных. Главные вариации отмечаются в строении костей плеча и бедра. В частности, плечевая и бедренная кости укорочены у дельфинов и некоторых ископаемых форм (ихтиозавров).
Существенные колебания отмечаются в количестве костей запястья и плюсны. Некоторые из примитивных ископаемых земноводных имели от 20 до 30 костей в этих частях скелета, но современные земноводные имеют лишь от 3 до 9 в запястье и от 3 до 12 в плюсне. Число костей в каждом пальце также колеблется у разных позвоночных. Фаланговая формула (количество костей от первого пальца до последнего) у современных амфибий, у которых имеется обычно четыре пальца, составляет 2—2-3—2 (саламандры), 2-2—3-3 (лягушки). Ихтиозавры имели от трех до восьми пальцев. Крылья современных птиц имеют по три пальца (пониженное число фаланг).
Общее количество костей у человека составляет 206. В ходе эволюции сформировался механизм остеофикации скелета эмбрионов, в котором существенное значение имеют кальций, фосфор, витамины Д и ряд ферментов.
У позвоночных кальциевый скелет представлен не кальцитом, а гидроксиапатитом.
В ходе эволюции существенное развитие приобрела и скелетная мускулатура. Выработался также механизм использования химической энергии пищи для сокращения мышц (20-35% от общего количества энергии) и для использования остальной энергии в качестве тепла, обеспечивающего тепловой режим организма.
Пищеварительная системаУ простейших органами пищеварения являются пищеварительные вакуоли. Для этих животных характерно внутриклеточное пищеварение, которое является самым древним.
У кишечнополостных и червей из эмбрионального бластопора развивается ротовое отверстие. У кишечнополостных рот открывается в гастральную полость, которая является полостью первичной кишки и заканчивается слепо (анального отверстия нет). В этой полости наряду с внутриклеточным пищеварением появляется внеклеточное. На более высоких уровнях эволюции внутриклеточное пищеварение теряется полностью, уступая место внеклеточному (полостному) как более прогрессивному способу. Однако внутриклеточное пищеварение все же сохраняется у организмов на всех ступенях развития, но лишь в виде фагоцитоза, выполняющего защитную роль.
У ресничных и плоских червей строение пищеварительной системы еще сохраняет сходство с пищеварительной системой кишеч-нополостных. Анального отверстия нет, поэтому выброс пищеварительных остатков происходит через рот. Но у круглых червей пищеварительная система уже несколько усложняется, поскольку пищеварительная трубка заметно подразделяется на передний, средний и задний отделы, из которых передний и задний отделы имеют эктодермальное происхождение, средний — энтодермальное. Впервые развивается анальное отверстие за счет впячивания эктодермы в каудальную часть тела и соединения ее с полостью первичной кишки. У кольчатых червей пищеварительный тракт уже представлен глоткой, пищеводом, зобом, желудком, средней и задней кишками.
Дальнейшее усложнение пищеварительного тракта продолжается у членистоногих и моллюсков, у которых развиваются ротовые придатки вокруг рта, которые служат для измельчения пищи, в кишечнике развиваются отростки, что увеличивает его поверхность. Развиваются пищеварительные железы, которые являются аналогом печени позвоночных.
У хордовых в ходе эволюции продолжается дифференциация кишечной трубки, причем более глубокая, в результате чего происходит увеличение кишечной поверхности для всасывания продуктов пищеварения. Возникают железы, участвующие в пищеварении. Пищеварительная система всех хордовых в основном имеет энтодермальное происхождение (кроме ротового отдела и заднего кишечника, имеющих эктодермальное происхождение). У позвоночных кишечная трубка дифференцируется на ротовую полость, глотку, пищевод, желудок, тонкий и толстый кишечники, которые, однако, не у всех организмов (классов) дифференцированы полностью. Начиная с круглоротых, у позвоночных развивается печень, а начиная с рыб — поджелудочная железа.
У рыб развиваются костные пластинки и зубы. У земноводных впервые появляются слюнные железы, заканчивается обособление желудка, тонкой и толстой кишок, клоаки, развиваются однорядные зубы. Очень интенсивно развивается язык. У рептилий намечается некоторая дифференцировка зубов, часть слюнных желез преобразуется в ядовитые, углубляется обособление желудка, в кишечнике развивается зачаток слепой кишки. У птиц способность к полету сопровождается изменениями в пищеварительной системе. Вместо челюстей и зубов у них появляется клюв, а в пищеводе образуется растяжимое расширение, называемое зобом. Желудок разделен на железистый и мускульный отделы, в которых происходит хжлическая и механическая переработка пищи (соответственно). Увеличивается в длину толстый отдел кишечника, но редуцируется язык.
У млекопитающих развитие пищеварительной системы достигает верхнего предела. Имея значительную длину, она характеризуется большей расчлененностью. Дифференцируются зубы, развивается пищевод. Желудок, состоит из нескольких слоев и содержит много пищеварительных желез. У некоторых травоядных (жвачных копытных) желудок является многокамерным. Кишечник дифференцирован на тонкий, толстый и прямой отделы. Значительной дифференцировки достигают печень, поджелудочная железа, их протоки впадают в передний отдел тонкого кишечника. Происходит удлинение толстого кишечника, появляются слепая кишка, и аппендикс. У многих млекопитающих получила значительное развитие слепая кишка, достигая '/g длины всего кишечника. Толстая кишка, заканчивается анусом, отделенным от мочеполового отверстия промежностью. У яйцекладущих толстая кишка заканчивается клоакой.
У человека в состав пищеварительной системы входит рот, глотка, пищевод, желудок, кишечник, слюнные железы, печень, желчный пузырь и панкреатическая железа Все отделы желудочно-кишечного тракта построены из четырех слоев.
Человек является всеядным существом, поскольку его пищеварительная система обеспечивает метаболизирование как растительной, так и животной пищи (1).
Главными углеводами, содержащимися в растительной пище, являются крахмал и целлюлоза, а в животной пище —гликоген.
Таблица 1
Функции отделов пищеварительной системы человека
| Отделы пищеварительной системы | Ротовая полость | Пищевод | Желудок | Тонкий кишечник | Толстый кишечник |
| Основные функции | Поступление пищи, частичное переваривание крахмала | Переход пищи в желудок | Переваривание белков | Переваривание и всасывание | Реадсорбция солей и воды, формирование каловых масс |
| Ферменты | Амилаза | Пепсин | Трипсин, липаза | ||
| Источники | Слюнные железы | Желудочные железы | Панкреас, тонкий кишечник | ||
| Образуемые | Сложные сахара | Крупномолекулярные фрагменты белков, некоторые аминокислоты | Жирные кислоты, глице-рол, простые сахара, аминокислоты, фрагменты белков |
Переваривание углеводов начинается в полости рта, где слюнная амилаза, (птиалин) действует на крахмал и гликоген, превращая их в смесь Д-глюкозы, мальтозы и олигосахаридов. Переваривание углеводов завершается в тонком кишечнике, где переваренный под действием птиалина крахмал подвергается дальнейшему воздействию панкреатической амилазы, конвертируется в мальтозу. В конечном итоге вся мальтоза под действием маль-тазы конвертируется в глюкозу.
Все реакции происходят с участием воды (гидролиз) и ферментов:
Рот, желудок:
![]()
Тонкий кишечник:
![]()
![]()
![]()
![]()
Непереваренная целлюлоза способствует нормальной перистальтике кишечника у млекопитающих.
Переваривание жиров начинается в тонком отделе кишечника, где панкреатическая липаза с помощью желчных кислот и белка колипазы конвертирует жиры в глицерол и жирные кислоты. Одна молекула жира дает одну молекулу глицерола и три молекулы жирной кислоты:
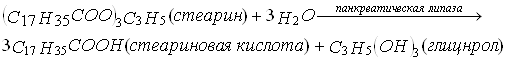
Перевариванию липидов в тонком отделе кишечника способствуют соли желчных кислот, которые являются производными холевой кислоты. Эти соли поступают из печени в желчь, которая затем изливается в переднюю часть тонкого кишечника.
Белки начинают перевариваться (гидролизуются) в желудке под воздействием пепсина и соляной кислоты, выделение которой клетками желудка происходит под действием гормона — гастри-на. Образованные продукты подвергаются повторному гидролизу трипсином и хемотрипсином, предшественники которых вырабатываются в клетках поджелудочной железы, до полипептидов, после чего последние гидролизуются пептидазами до аминокислот.
Однако в желудочно-кишечном тракте человека не все белки перевариваются полностью. Например, такой белок, как кератин, переваривается лишь частично. Многие белки злаковых также перевариваются не полностью, поскольку они покрыты непереваривающейся целлюлозой.
Дыхательная системаВ ходе эволюции у организмов развилась высокоорганизованная система органов дыхания.
У многих беспозвоночных животных, например, у кишечно-полостных, плоских и круглых червей еще нет специальных органов дыхания. У них газообмен со средой осуществляется всей поверхностью тела. Впервые органы дыхания встречаются у морских кольчатых червей и водных членистоногих в виде пористых жабер, располагающихся по обеим сторонам тела и обильно снабжаемыми кровью. У моллюсков в мантийной полости развиваются пластинчатые жабры. Однако уже у наземных членистоногих в углублениях тела имеются трахеи или листовидные легкие.
У водных хордовых дыхательная система связана с кишечником, причем наиболее просто она организована у ланцетника, у которого стенка глотки (передний отдел кишечника) пронизана жаберными щелями, пронизывающими весь передний отдел кишечника и открывающимися в околожаберную полость. У наземных хордовых жаберные щели развиваются лишь в эмбриональный период, после чего исчезают. Дыхание у них осуществляется легкими, которые развиваются из выпячиваний кишечной стенки.
Жаберный аппарат у хордовых эволюционировал в направлении образования жаберных лепестков. В частности, у рыб развилось 4-7 жаберных мешков, являющихся щелями между жаберными дужками и содержащими большое количество лепестков, которые пронизаны капиллярами. У рыб в дыхании участвует также воздушный пузырь.
Легкие эволюционировали в направлении увеличения дыхательной поверхности, что привело к образованию бронхов и бронхиол. Впервые легкие появляются у земноводных, представляя собой полые мешки. Однако у них в дыхании еще участвует кожа. У рептилий происходит дальнейшая дифференцировка органов дыхания, В частности, строение легких усложняется, в результате чего в них образуются ячеистые структуры (перекладины). Появляются бронхи. У птиц легкие представляют собой губчатые образования, развиваются разветвления бронхов.
У млекопитающих развитие дыхательной системы достигает вершины Наряду с дальнейшим совершенствованием легких чрезвычайному развитию подвергаются воздухоносные пути.
Развиваются бронхи второго, третьего и четвертого порядков, а также бронхиолы и альвеолы. Развивается диафрагма, которая отделяет грудную полость от брюшной. Наличие высокоспециализированных органов дыхания обеспечивает очень эффективный газовый обмен в легких (внешнее дыхание) и в тканях (внутреннее дыхание). Появляются гортанные хрящи.
Эволюция дыхательной системы зависима от колебаний в содержании атмосферного кислорода и углекислоты, поскольку жабры не чувствительны к изменению газового состава воздуха, а кожное дыхание не компенсирует этого недостатка жабер. Начиная с позднего палеозоя, содержание атмосферного кислорода колебалось в разные времена. Хотя фактические величины этих колебаний не определены, тем не менее, предполагают, что максимум содержания кислорода в атмосфере достигал 35%, а затем падал до 15% (при 21% в настоящее время).
Повышенное содержание кислорода и одновременное уменьшение углекислоты повышало эффективность легочного дыхания, помогая позвоночным в завоевании суши. Более того, есть основания к допущению влияния кислородного ритма даже на эволюцию основных групп животных.
Кровеносная и лимфатическая системы
У многих беспозвоночных животных сосудистой системы как таковой нет. В частности, у губок, кишечнополостных и плоских червей транспорт питательных веществ и кислорода в разные части тела осуществляется путем диффузии их тканевых жидкостей. Однако у организмов других групп появляются сосуды, обеспечивающие циркуляцию тканевой жидкости в определенных направлениях. В начале развиваются примитивные сосуды, но в результате развития в сосудах мышечной ткани они в дальнейшем оказываются способными к сокращению. Параллельно развивается в кровь жидкость, заполняющая сосуды.
Кровеносная система впервые встречается (развивается) у кольчатых червей, у которых она является замкнутой, представляя собой систему, состоящую из верхнего спинного и нижнего брюшного сосудов, объединенных кольцевыми сосудами, опоясывающими кишечник. Кроме того, от спинного и брюшного сосудов отходят более мелкие сосуды к стенкам тела. К головному отделу кровь идет по спинной стороне, а к заднему — по брюшной.
У членистоногих, кровеносная система не замкнута, т. к. сосуды открываются в полость тела. Спинной сосуд разделяется перегородками (клапанами) на отдельные камеры — сердца, сокращения которых заставляют проходить кровь в артерии, а из последних — в пространства между органами. Из этих пространств кровь затем поступает в околосердечную полость.
У моллюсков кровеносная система тоже не замкнута и представлена сердцем, состоящим из нескольких предсердий и желудочка, а также артериальными и венозными сосудами. Вены впадают в предсердие, тогда как артерии отходят от желудочка.
Кровь у беспозвоночных несет кислород к тканям, удаляет двуокись углерода и транспортирует питательные вещества, гормоны, а также шлаковые продукты азотистого обмена. Кровь беспозвоночных по сравнению с кровью позвоночных содержит меньшее количество белков и форменных элементов. Единственным форменным элементом крови большинства беспозвоночных являются лейкоциты. Поэтому очень простой состав крови у беспозвоночных компенсируется разными способами транспорта кислорода, при которых используются гемоглобин, гемоцианин или другие дыхательные пигменты.
Предполагают, что развитие гемоглобина беспозвоночных шло независимо от развития гемоглобина позвоночных, поскольку этот пигмент у них обладает несколько отличной функцией. В частности, у беспозвоночных он выполняет запасательную функцию, обеспечивая защиту против недостатка кислорода в неблагоприятной среде.
По мере усложнения организации живых существ отмечается повышение количества кислорода в крови, т. е. повышается кислородная способность дыхательных пигментов (2).
Считают, что гемоглобин имеет древний возраст. Поскольку некоторые бактерии синтезируют гемопротеин, содержащий участки с аминокислотным составом, близким к последовательностям глобинов высших эукариотов, то предполагают, что гемоглобин впервые возник еще у прокариот. Однако не исключено, что у эукариот и прокариот имела место конвергентная эволюция.
Гемоглобин встречается у плоских, круглых и кольчатых червей, членистоногих, моллюсков, иглокожих, рыб, земноводных, рептилий, птиц, млекопитающих. Молекулярная масса гемоглобина беспозвоночных колеблется в пределах 17 000-2 750 000. У многих членистоногих и моллюсков встречается гемопианин, представляющий собой медьсодержащий белок молекулярной массой 400 000—5 000 000. Дыхательный пигмент обеспечивает у беспозвоночных буферную способность крови. Кроме того он ответственен и за осмотический баланс кровяной жидкости. Большинство насекомых не содержит совсем дыхательного пигмента, имея в крови растворенный кислород.
Таблица 2
Кислородная способность различных дыхательных пигментов
| Животные | Пигмент | Цвет пигмента | Метал-лоэлемент | Локализация пигмента | Количество кислорода (в мл на 100 мл крови) |
| Млекопитающие | Гемоглобин | Красный | Железо | Эритроциты | 25 |
| Птицы | 18,5 | ||||
| Рептилии | 9 | ||||
| Земноводные | 12 | ||||
| Рыбы | 9 | ||||
| Аннелиды | Плазма | 6,5 | |||
| Моллюски | 1,5 | ||||
| Моллюски | Гемоцианин | Голубой | Медь | Плазма | 2,8 |
| Аннелиды | Хлоро-круорин | Зеленый | Железо | Плазма | 9 |
| Аннелиды | Гемоэрит-рин | Красный | Железо | Эритроциты | 2 |
У полухордовых кровеносные сосуды еще не выстланы эндотелием. Сердце урохордовых представляет собой мускульное выпячивание около желудка. У хордовых кровеносная система характеризуется дальнейшим совершенствованием. У них развиваются сердце, сосуды, кровь.
У ланцетника кровеносная система является замкнутой, но сердца еще нет; его функцию выполняет передний отдел крупного сосуда в виде брюшной аорты. Впервые сердце появляется у водных позвоночных. В частности, у рыб сердце двухкамерное с предсердием и желудочком.
Круг кровообращения один, в котором артериальная и венозная кровь не смешиваются. Из сердца венозная кровь идет к жабрам, где, окисляясь, она становится артериальной, после чего расходится по артериям ко всем частям тела. К сердцу кровь вновь доставляется венами. У пресмыкающихся желудочек разделен неполной перегородкой на правую (венозную) и левую (артериальную) половины. У крокодилов желудочек разделен полностью. Однако у пресмыкающихся разделения артериального и венозного тока крови еще не происходит.
У наземных позвоночных развивается трех-, а затем четырехкамерное сердце, что явилось результатом очень крупного ароморфоза. У птиц и млекопитающих сердце построено из мышц и разделено на четыре камеры в виде двух предсердий и двух желудочков Имеются два круга кровообращения, один из которых большой, второй малый. Благодаря этим кругам артериальная и венозная кровь не смешиваются. Артериальная кровь выходит из сердца, венозная вступает в сердце.
Сердце позвоночных относят к миогенному типу, поскольку его сокращения начинаются внутри за счет сократительных элементов, называемых миофибриллами. У рыб, земноводных и пресмыкающихся сокращения сердца начинаются в тонкой мышечной стенке венозного синуса и продолжаются посредством внутренней проводящей системы через предсердие, потом к желудочкам. У птиц и млекопитающих венозного синуса нет.
У позвоночных усложняются строение и функции крови. Кровь состоит из форменных элементов и плазмы, увеличивается их масса. Например, в крови млекопитающих клеточная масса крови составляет 45% , плазменная — 55% . Гемоглобин содержится в эритроцитах, которые у всех позвоночных, кроме млекопитающих, содержат ядра. У земноводных клетки крови очень крупные по сравнению с пресмыкающимися, птицами и млекопитающими. В 1 мл крови человека содержится 5 млн эритроцитов и 8—10 тыс лейкоцитов. Группы крови современных людей также являются результатом их эволюции В плазме крови содержатся ферменты, гормоны и другие соединения, необходимые для метаболизма клеток.
В ходе эволюции у высших животных взаимосвязи обмена веществ разных органов, обеспечиваемые кровью, получили дальнейшее усложнение. Одна из важнейших функций крови заключается в транспорте кислорода от легких к тканям и в транспорте СОд, являющимся конечным продуктом дыхания, от тканей к легким, причем эта функция крови развивалась в направлении способности транспортировать большие объемы кислорода. Основная часть кислорода, потребляемого организмом человека, транспортируется гемоглобином эритроцитов, масса которого составляет примерно треть массы эритроцитов. Как неоднократно отмечалось ранее, гемоглобин представляет собой очень сложный белок, состоящий из двух а-полипептидных цепей и двух b-полипептид-ных цепей, каждый из которых присоединяет по одной гемовой группе. Его происхождение является очень древним
Эритроциты участвуют также в транспорте СО2 из тканей в легкие, откуда он выводится в процессе выдыхания. Эволюционным приобретением млекопитающих, в том числе человека, является то, что их гемоглобин приспособлен как для транспорта кислорода, так и для транспорта СОд, причем оба эти транспорта взаимоусиливаемы.
Кровь ответственна и за перенос питательных веществ из тонкого кишечника в печень и другие органы, а также за выведение шлаков из тканей в почки. Эти функции тоже развивались в направлении увеличения количественной способности в связи с увеличивающейся интенсификацией метаболизма по мере усложнения организации живых существ.
Диффузия кислорода, двуокиси углерода, питательных веществ и метаболитов происходит лишь в капиллярах.
Параллельно с развитием кровеносной системы шло развитие лимфатической системы, которая впервые появилась у хордовых. Эта система в ходе эволюции обособилась из венозной системы.
Лимфатическая система состоит из лимфатических сосудов, лимфатических узлов и лимфы.
Лимфатические сосуды построены из эластичной и мышечной тканей и выстланы внутри эндотелием. Для них характерен такой же внутренний объем, как и у кровеносных капилляров. Лимфатические сосуды очень разветвлены. Они в виде очень тонких капилляров начинаются с межклеточных пространств, формируя затем более крупные сосуды. С помощью лимфатических сосудов лимфа проходит от тканей в венозное русло. Все лимфатические сосуды сливаются в один общий сборный сосуд (млечный путь). Одиночные лимфатические узлы обнаруживаются в толще слизистой оболочки и подслизистой основы кишечника у рыб, земноводных, пресмыкающихся, птиц и млекопитающих, но групповые лимфатические узлы развиваются лишь у млекопитающих.
У позвоночных по мере усложнения их организации наступает приближение лимфатических капилляров к клеткам и лимфоид-ным узелкам. Например, расстояние между лимфатическими капиллярами, кишечными эпителиоцитами и лимфоидными узелками у рыб составляет 150-180 мкм, но затем прогрессивно уменьшается у земноводных, пресмыкающихся и птиц, составляя у млекопитающих всего лишь 50-30 мкм.
Выделительная системаУ простейших выделение продуктов обмена осуществляется через всю поверхность тела путем диффузии. Этот же механизм выделения действует также у губок и иглокожих, однако у некоторых из них освобождение от продуктов обмена осуществляется с помощью фагоцитоза.
У плоских червей, ротифер и некоторых немертин развивается примитивная выделительная система в виде протонефридий, которые представляют собой слепые канальцы, разветвленные по телу и снабженные на конце пламенной клеткой, проникающей в канадец (рис. 193). Продукты выделения поступают в цитоплазму пламенных клеток, а затем в канальцы, объединяющиеся и открывающиеся наружу в виде пор. Через эти поры продукты выделения и выходят в окружающую среду.
У круглых червей выделительная система в виде протонефридий представлена внутрицитоплазматическими каналами, проникающими в полость тела.
У кольчатых червей, у которых появляется вторичная полость (целом), развивается выделительная система в виде метанефридий, представляющих собой извитые канальцы, располагающиеся по два в каждом сегменте тела Эти канальцы открываются во вторичную полость тела одним концом (нефро-стомом), снабженным мерцательными ресничками, а наружу (на боковой стороне гельминта) — другим (нефри-диопорой).
У кольчатых червей метанефридий очень многочисленны. У насекомых и других членистоногих выделительная система представлена слепыми канальцами (мальпигиевыми сосудами), включенными в целом и открывающимися в анальное отверстие. Эти канальцы выстланы полусферическим эпителием.
У моллюсков также имеются метанефридий, но они представлены всего лишь одной парой, каждая из которых открывается нефростомом в околосердечную полость, а другим концом в бронхиальную полость. Таким образом, выделительную функцию осуществляет структура, формируемая канальцами каждого метанефридия.
У хордовых выделительная система характеризуется дальнейшим усложнением и разнообразием, определяемым переходом от нефридий у низших хордовых к почкам трех «поколений» у позвоночных. У бесчерепных, в частности у ланцетника, выделительная система состоит из располагающихся в районе жаберной щели сегментированных канальцев, в которые жидкость поступает из це-лома через нефростомы и выводится в околожаберную полость через не-фридиоспоры. У ланцетника количество таких нефридий достигает до 100 пар.
У позвоночных в период эмбриогенеза из мезодермы развивается три «поколения» почек — предпочка, или головная почка (пронефрос) у круглоротых (миксин), первичная, или туловищная почка (мезонефрос) у круглоротых (миног), рыб и земноводных и вторичная, или тазовая почка (метанефрос) у пресмыкающихся, птиц и млекопитающих.
Усложнение выделительной системы отмечается в первую очередь в строении почечных канальцев. В предпочке почечные канальцы еще похожи на метанефридий, т. е. они являются канальцами, открывающимися мерцательным нефростомом в целом, а количество канальцев составляет 4-10. У миксин пара предпочечных протоков тянется от шейной части до клоаки.
Дальнейшее усложнение связано с развитием большей эффективности фильтрующей системы внутри канальцев. Для первичной почки характерно увеличение извилистости капилляр внутри канальцев, которых может быть около 100, концентрация их в клубочки и локализация в особых камерах (капсулах), образуемых стенкой канальцев, что приводит к образованию так называемых мальпигиевых телец. Первичные почки являются овальными телами.
Вторичная почка представляет собой уже дольчатый орган, канальцы которого (их количество доходит до 1 млн) открываются в мочеточник, переходящий в мочевой пузырь (у птиц последнего нет). У земноводных, пресмыкающихся и птиц протоки выделительной системы открываются непосредственно в клоаку, тогда как у млекопитающих мочевой пузырь переходит в мочеиспускательный канал, открывающийся наружу.
Почки разных поколений развиваются из мезодермы. Пронефрос закладывается из парных зачатков. Разрастаясь, они сливаются в так называемый предпочечный проток. Закладка мезонефроса происходит сзади пронефроса. В процессе развития проток мезонефроса расщепляется на так называемые вольфов и мюллеров каналы.
Метанефрос развивается позади мезонефроса. Канальцы этой почки открываются в мочеточник, который отделяется от вольфова канала, развивающегося в семяпровод у самцов, тогда как мюллеров канал сохраняется, выполняя роль яйцевода у самок.
У взрослых форм круглоротых, рыб и земноводных функционируют первичные почки, но у миксин еще сохраняются некоторые особенности структуры предпочки, у личиночных форм рыб и земноводных предпочки сохраняются полностью. У взрослых форм пресмыкающихся функционируют только вторичные почки. Они представляют собой парные сплошные или дольчатые тела, располагающиеся под позвоночником.
У млекопитающих выделительная система помимо почек представлена мочеточниками, мочевым пузырем и мочеиспускательным каналом. Резко возрастает «производительность» выделительной системы. Почки здорового человека образуют ежедневно около 2 л мочи. Половина твердых веществ суточного количества мочи составляет мочевина (около 30 г), которая является главным шлаковым продуктом белкового метаболизма.
Нервная системаУ простейших нервной системы в той форме, как ее обычно понимают, не существует, однако в их цитоплазме имеется аппарат возбудимости и движения. В частности, у парамеций центральная нейромоторная масса цитоплазмы связывается фибриллами с основанием каждой реснички, благодаря чему образуется примитивный координирующий центр и проводящая система.
Начало нервной системы простейшего типа отмечается у ки-шечнополостных. Причем первым этапом в ее возникновении является дифференциация эпителиальных клеток в нейро-сенсорные элементы. Поверхность этих клеток выполняет роль рецептора, в то время как от их основания разветвляются нервные нити в сторону подлежащих мышечных клеток. Истинные нервные клетки, обладающие функцией проводимости, наблюдаются впервые также у кишечнополостных, у которых впервые появляются нейроны, секретирующие катехоламины.
Дальнейшее развитие нервной системы оказалось связанным с переходом организмов от радиальной симметрии к билатеральной и заключалось в концентрации нервных клеток в разных частях тела.
У плоских червей концентрация нервных клеток завершилась формированием парных головных нервных узлов с отходящими от них нервными волокнами к органам чувств и парных нервных стволов, проходящих по телу от головной части к каудальной. У круглых червей уже встречается окологлоточное нервное кольцо, образуемое за счет слияния головных нервных узлов, а у кольчатых червей даже развивается нервная цепочка за счет образования парных нервных узлов в сегментах тела.
У членистоногих отмечается дальнейшая концентрация нервных клеток, в результате чего обособляются нервные центры, развиваются органы чувств.
Эволюция центральной нервной системы у беспозвоночных и позвоночных проходила, главным образом, в направлении топографической и ультраструктурной перестроек базисных нервных и нервно-эндокринных структур. Она генерирована из одного источника — нейрогенного эпителия, который формирует вначале нервную пластинку, а затем и нервную трубку.
У низших хордовых центральная нервная система представлена трубкой, в головной части которой развивается расширение, представляющее собой зачатки мозга, а полость в расширении (невроцель) — желудочек. Периферическая нервная система представлена отходящими нервами. Нейросекреторные клетки осуществляют сенсорную, проводящую и секреторную функции.
По мере усложнения организмов нервная система эволюционировала в направлении цефализации и повышения ассоциации нейронов У позвоночных нервная система представлена головным и спинным мозгом и периферическими нервами. Головной и спинной мозг составляют центральную нервную систему. Нервы составляют периферическую нервную систему (соматическую и автономную).
У позвоночных центральная нервная система имеет существенные вариации в зависимости от филогенетического положения организма.
Головной мозг состоит из пяти отделов — переднего, промежуточного, среднего, мозжечка и продолговатого, которые у животных, принадлежащих к разным классам, характеризуются различной степенью развития. Для позвоночных характерна повышающаяся дифференциация серого и белого вещества. Основная функция спинного мозга заключается в интеграции различных сенсомоторных механизмов.
Наименее развит головной мозг у круглоротых, у которых все отделы этого мозга располагаются один за другим.
У рыб центральная нервная система имеет вид трубки, расширяясь в головной части. Дифференциация головного мозга более выражена. В частности, увеличен передний мозг, хорошо выражены зрительные доли среднего мозга, развит мозжечок. Имеется 10 пар черепномозговых нервов.
У земноводных в связи с выходом их на сушу нервная система характеризуется более сложным строением по сравнению с рыбами, в частности, большим развитием и полным разделением мозга на полушария. Передний мозг (полушария) содержит серое вещество. Дно желудочка, бока и крыша также содержат серое вещество, что свидетельствует о появлении у земноводных первичного мозгового свода (архепаллиума).
Из головного мозга как и у рыб выходят 10 пар черепно-мозговых нервов. Симпатическая нервная система хорошо развита. Более совершенное зрение. Наряду с внутренним ухом, развитым у рыб, у них появляется среднее ухо. Большего развития достигает орган обоняния.
У рептилий особенностью нервной системы является прогрессивное развитие всех отделов головного мозга, характерное для наземных животных. В частности, значительно увеличены полушария мозга и мозговой свод. Увеличивается архепаллиум, дифференцируются закладки вторичного мозгового свода (неопаллиума). На поверхности полушарий впервые появляется кора, увеличивается мозжечок, становясь более выпуклым, продолговатый мозг формирует изгиб. Из головного мозга выходят 12 пар черепно-мозговых нервов. Усложняются нейронные сети. Еще в большей мере развиваются органы чувств.
У птиц прогрессивное развитие головного мозга заключалось в увеличении полушарий и зрительной доли, дальнейшем развитии мозжечка.
У млекопитающих увеличение полушарий головного мозга сопровождалось развитием коры, образованием в ней извилин и борозд, завершением развития вторичного мозгового свода (неопаллиума), прогрессивным развитием мозжечка Основной структурой и функциональной единицей нервной системы является нейрон, развитие которого у человека достигает наивысшего уровня. Происходит дифференциация нейронов на сенсорные, моторные и соединительные. Нейроны синтезируют большое количество нейропептидов, ответственных за коммуникацию клеток и другие важные функции.
Высокого уровня достигает развитие сенсорной системы, в которой наиболее сложными являются органы зрения и слуха. В ходе эволюции зрение впервые появляется у членистоногих. У них оно представлено парой сложных фасеточных глаз, разделенных на омматидии, каждая из которых может различать лишь часть объекта. Насекомые обладают цветовым и объемным зрением. Дальнейшее совершенствование органа зрения характерно для рыб и земноводных.
У рептилий уже отмечается возможность изменения кривизны хрусталика, что ведет к улучшению зрения. В ресничном теле уже развита поперечно-полосатая мускулатура. У земноводных в среднем ухе содержится слуховая косточка, а у рептилий уже увеличилась улитка. Органы зрения и слуха достигают совершенства у млекопитающих, особенно у человека.
Предполагают, что высокая скорость эволюции млекопитающих по сравнению с другими организмами объясняется наличием у них мозга и крупных полушарий. Особенно быстро шло развитие головного мозга у гоминид в плейстоцене, в течение которого объем мозга возрос у них в три раза.
Это является примером наиболее быстрых макроэволюционных явлений. Появление нервных клеток означало качественно новый этап эволюции, позволивший высшим животным и человеку регулировать условия среды обитания, а с этим и выживание.
Эндокринная системаЭндокринной системы в том виде, как ее понимают в применении к высшим организмам, у беспозвоночных нет. Однако у членистоногих в особых железах синтезируются гормоны, которые влияют на линьку (у ракообразных), на созревание яиц и протекание диапаузы, линьку и развитие взрослых форм (у насекомых) и другие свойства.
Таблица 3
Эндокриннная система человека
| Железы | Гормоны | Гормональные эффекты |
| Гипофиз | Соматотропин, адренокорти-костероидный гормон | Рост |
| Гонадотропный гормон | Половые железы | |
| Тиротропный гормон | Щитовидная железа | |
| Вазопрессин | Почки | |
| Щитовидная железа | Тироксин | Метаболизм |
| Паращитовидная железа | Паращитовидный гормон | Кальций крови |
| Надпочечники | Разные гормоны (более 30) | Метаболизм |
| Островки Лалгерганса | Инсулин | Метаболизм углеводов |
| Яичники | Эстрогены, прогестероны | Гениталии и половые признаки |
| Тестисы | Тестостерон |
Относительно совершенная система появляется у позвоночных. Уже у земноводных имеется гипофиз, надпочечники, щитовидная, поджелудочная и половые железы.
Чрезвычайного развития эндокринная система достигает у млекопитающих, у которых она представлена совокупностью желез внутренней секреции, выделяющих в кровь гормоны — химические регуляторы
Гипофиз вырабатывает гормоны, которые не только влияют на свойства организма, но и на синтез гормонов Другими железами внутренней секреции. В частности, гипофизарный гормон со-матотропин повышает частоту роста клеток, стимулирует рост и развитие тканей. Вазонрессин оказывает влияние на реабсорбцию воды почечными канальцами. В то же время гипофиз стимулирует секрецию тироксина, который повышает частоту метаболизма в клетках и тканях, а гипофизарный гонадотропин влияет на синтез половых гормонов.
Надпочечники секретируют ряд гормонов, среди которых наиболее важными являются кортизол, альдостерон и эпинефрин (адреналин). Кортизол регулирует метаболизм белков, жиров и углеводов, влияет на беременность, альдостерон — метаболизм натрия и калия, а эпинефрин стимулирует распад гликогена печени и повышает кровяное давление. Гормоны, получившие название эстро-генов, синтезируются в яичниках и тестисах из холестерола в ответ на сигналы, поступающие из мозга и других органов. Попадая в кровь, они разносятся затем к молочным железам и репродуктивным органам. Поскольку эстрогены растворимы в липидах, обычно содержащихся в мембранах, то они легко преодолевают мембранные барьеры и входят в ядро, где связываются с присутствующим в ядерном содержимом белком-рецептором. В дальнейшем комплекс эстроген + белковый рецептор связывается с регуляторными районами ряда генов и изменяют их экспрессию путем активирова-ния или ингибирования действия генов. Комплекс эстроген + рецептор регулирует также экспрессию гена, кодирующего синтез белка-рецептора для прогестерона.
Эндокринная система у высших организмов ответственна за гуморальную регуляцию роста и развития. Все гормоны представляют собой химические соединения, обладающие функциями сигналов, связывающих внутренние коммуникационные системы организма, чем помогают клеткам «чувствовать» и реагировать на изменяющиеся физиологические условия. Различают пептидные гормоны (все гормоны гипоталамуса, гипофиза, инсулин и глюкагон), стероидные (гормоны коры надпочечников, андрогены и эстрогены) и гормоны типа аминов (эпинефрин и тиреоидные гормоны).
В ходе эволюции у высших организмов сформировалась сложная система регуляторных взаимосвязей между железами, клетки которых продуцируют гормоны, и клетками (органами)-мишенями. Главным центром развитой эндокринной системы является гипоталамус. В ответ на сигналы, которые получает гипоталамус из центральной нервной системы, его клетки секретируют ряд гипоталамических регуляторных гормонов, каждый из которых в свою очередь регулирует секрецию одного из гипофизарных гормонов. Последние, оказавшись в крови, подходят к надпочечникам, щитовидной железе и стимулируют выработку разных гормонов, которые, попав в кровь, транспортируются к рецепторам гормонов, расположенных на поверхности клеток или тканей-мишеней этих гормонов. В клетках существуют внутриклеточные сигнальные посредники (передатчики), которые передают затем сигнал с клеточного (тканевого) рецептора гормона на какую-либо из специфических внутриклеточных структур или какой-либо фермент.
Репродуктивная системаУ простейших специальной репродуктивной системы нет. У губок и кишечнополостных такой системы тоже нет, их половые клетки выходят наружу через стенки тела. Репродуктивная система в виде половых желез и протоков для выведения половых клеток впервые развивается у плоских червей. Вопреки гермафродитизму, у плоских червей формируются семяпроводы и семяизвергательные каналы для мужских половых клеток, а также яйцеводы, матка и влагалище для женских половых клеток. Половые клетки развиваются в семенниках и яичниках.
У круглых червей уже возникает раздельнополость. Отмечается развитие копулятивного аппарата, необходимого для внутреннего оплодотворения.
У хордовых прогрессивное усложнение репродуктивной системы шло в направлении специализации женских и мужских половых желез и формирований приспособлений для внутреннего осеменения. Для ланцетника характерно наличие большого количества половых желез (26 пар), располагающихся в стенках околожаберной щели. Эти железы еще не имеют протоков для выведения половых клеток. Поэтому последние благодаря разрывам стенки желез вначале попадают в околожаберную щель, а затем наружу через атриопор. У круглоротых уже имеется непарная половая железа, но протоков еще нет. После созревания половые клетки оказываются в полости тела, откуда через половые отверстия выводятся наружу.
У рыб характерно увеличение половых желез. Яичник обычно непарный, а семенники часто парные. Выводные протоки половых желез у рыб уже связаны с выделительными канальцами почек. Сложность оплодотворения яйцеклеток в водной среде компенсируется выработкой огромного количества половых клеток (икры и молоки).
У земноводных половые железы являются парными как у самцов (семенники), так и у самок (яичники). У части амфибий (бесхвостых) еще сохраняется наружное оплодотворение, тогда как у хвостатых оплодотворение является внутренним. Их развитие происходит с метаморфозами.
У пресмыкающихся женские и мужские половые железы сходны по строению с такими железами земноводных, но у них сильнее развит правый яичник. Крокодилы продуцируют большое количество яиц, но мелких.
У птиц репродуктивная система тоже асимметрична, но больше развит левый яичник. Яйцевод является очень большим, причем делится на передний и задний отделы. Яичник очень увеличен в период кладки яиц.
После завершения откладывания яиц он уменьшается. Яйца относительно крупные.
Наибольшего усложнения репродуктивная система достигает у млекопитающих, для которых характерно только внутреннее оплодотворение. Происходит развитие совокупительных органов, приспособлений для развития плода в организме матери. Развиваются придаточные железы. Характерным является также сложное половое поведение организмов этого класса
Исключительное совершенствование отмечается в организации половых клеток, особенно мужских. У большинства беспозвоночных половые клетки обладают структурными компонентами, характерными для одноклеточных простейших и соматических клеток многоклеточных. В частности, акросома является первичной лизосомой. Имеются центриоли, митохондрии и жгутик с аксонемой. У организмов, приспособившихся к жизни на суше, после развития внутреннего оплодотворения сперматозоиды развились в вытянутые структуры, а в их хвосте появились дополнительные цитоскелетные элементы. У высших наземных организмов развиваются разные формы подвижности сперматозоидов и разные типы акросом, что зависит от строения яйцевых оболочек.
Список литературы:
· Биология с основами экологии. Серия «Учебники для вузов. Специальная литература» — СПб.: Издательство «Лань», 2000.
· Биология. (Учебник) Чебышев Н.В. и др. (2000
· Наглядная биохимия. Кольман Я., Рём К.-Г. (2000
Похожие работы
... рассказано в книге Хукера "Эти удивительные ледниковые эпохи". Приведенные примеры еще раз подтверждают модель экранированной Земли. В прошлом растительная и животная жизнь существовала от полюса и до полюса. 2.3 ВОЗНИКНОВЕНИЕ ЖИЗНИ НА ПЛАНЕТЕ Существуют и другие представления о возникновении нашей планеты и жизни на ней. Вот одно из них. Возникновению живого вещества на Земле (и, как можно ...
... весьма любопытную теорию Бергсона. Согласно мнению французского философа, сущность жизни заключается именно в совершенно непредсказуемых, спонтанных действиях, так что ни о каких целях живых существ быть не может , поскольку цель предполагает представление о будущем результате, жизнь заключается именно в активности вопреки предвидениям – предвидеть можно лишь действия механизмов в неживой материи ...
рганов и систем его организма. Так как темпы научно-технического прогресса сейчас приобретают стремительный характер и предъявляют серьезные требования к человеку и другим живым существам, проблема актуальности биоритмов является сегодня самой важнейшей. Бездумное отношение человека к самому себе, как и к окружающей природе, часто является следствием незнания биологических законов, эволюционных ...
... . Таким образом в середине XX века был экспериментально осуществлен абиогенный синтез белковоподобных и др. органических веществ в условиях, воспроизводящих условия первобытной Земли. Гипотеза Опарина о возникновении жизни на Земле опирается на представление о постепенном усложнении химической структуры и морфологического облика предшественников жизни (пробионтов) на пути к живым организмам. На ...
0 комментариев