Всі процеси життєдіяльності організмів супроводжуються появою в клітинах і тканинах електрорушійних сил. Електричні явища відіграють велику роль у найважливіших фізіологічних процесах: збудженні клітин і проведенні збудження по клітинам.
Завдяки безпосередньому зв'язку біопотенціалів з метаболічними процесами й фізіологічним станом клітин вони є чутливими й точно вимірними показниками різних змін у клітинах у нормі й при патології. Для більш ефективного й плідного використання електрофізичних методів у медицині необхідне з'ясування механізмів виникнення біоелектричних потенціалів.
Всі клітини збудливих тканин при дії різних подразників достатньої сили здатні переходити в стан збудження. Обов'язковою ознакою збудження є зміна електричного стану клітинної мембрани. Досвід показує, що збуджена ділянка клітини стає електронегативною стосовно не збудженої, що показує на перерозподіл іонів у збудженій ділянці. При збудженні він має тимчасовий характер, і після закінчення збудження знову відновлюється вихідний потенціал спокою. Загальна зміна різниці потенціалів між клітиною й середовищем, що відбувається при граничному й надпороговому збудженні клітин, називається потенціалом дії. Потенціали дії забезпечують проведення збудження по нервових волокнах й ініціюють процеси скорочення м'язових і секреції залізистих клітин.
Фаза швидкої деполяризації
Виникнення потенціалу дії (ПД) пов'язане зі збільшенням проникності мембрани для іонів натрію (в 20 разів у порівнянні із проникністю для К+, і в 500 разів у порівнянні з вихідною проникністю Na+) і наступним посиленням дифузії цих іонів по концентраційному градієнті усередину клітини, що приводить до зміни (зменшенню) мембранного потенціалу. Зменшення мембранного потенціалу приводить до збільшення проникності мембрани для натрію, а збільшення проникності супроводжується посиленням дифузії натрію в цитоплазму, що викликає ще більш значну деполяризацію мембрани. Завдяки наявності позитивного зворотного зв'язку деполяризація мембрани при збудженні відбувається із прискоренням і потік іонів натрію в клітину увесь час зростає. Інтенсивність же потоку іонів калію, спрямованого із клітини назовні, у перші моменти збудження залишається як і на початку. Посилений потік позитивно заряджених іонів натрію усередину клітини викликає спочатку зникнення надлишкового негативного заряду на внутрішній поверхні мембрани, а потім приводить до перезарядження мембрани.
Надходження іонів натрію відбувається доти, поки внутрішня поверхня мембрани не придбає позитивний заряд, достатній для зрівноважування градієнта концентрації натрію й припинення його подальшого переходу усередину клітини.
Натрієвий механізм виникнення ПД підтверджують досвіди зі зміною зовнішньою й внутрішньою концентрацією цих іонів. Було показано, що десятикратній зміні концентрації іонів натрію в зовнішнім або внутрішнім середовищі клітини відповідає зміна ПД на 58 мВ. При повному видаленні іонів натрію з оточуючої клітину рідини ПД не виникали.
Таким чином, установлено, що ПД виникають у результаті надлишкової в порівнянні зі спокоєм дифузії іонів натрію з оточуючої рідини усередину клітини.
Період, протягом якого проникність мембрани для іонів натрію при порушенні клітини зростає, є невеликим (0,5-1 мс); слідом за цим спостерігається підвищення проникності мембрани для іонів калію й, отже, посилення дифузії цих іонів із клітини назовні.
Фаза реполяризації
Збільшення іонного потоку калію, спрямованого із клітини назовні, призводить до зменшення мембранного потенціалу, що у свою чергу обумовлює зменшення проникності мембрани для іонів натрію, що, як указувалося, є функцією мембранного потенціалу.
Таким чином, другий етап порушення характеризується тим, що потік іонів калію із клітини назовні зростає, а зустрічний потік іонів натрію зменшується. Це триває, поки не відбудеться відновлення потенціалу спокою - реполяризація мембрани. Після цього проникність для іонів калію також падає до вихідної величини. Зовнішня поверхня мембрани за рахунок позитивно заряджених іонів калію, що вийшли в середовище, знову здобуває позитивний потенціал стосовно внутрішнього.
Фаза слідових потенціалів
У кінцевій фазі порушення відбувається уповільнення відновлення мембранного потенціалу спокою, і при цьому реєструються слідові реакції у вигляді слідової деполяризації й гіперполяризації, обумовлені повільним відновленням вихідної проникності для іонів К+.
Таким чином, формування ПД обумовлене двома іонними потоками через мембрану: потік іонів натрію усередину клітини призводить до перезарядження мембрани, а протилежно спрямований потік іонів калію спричиняється відновлення вихідного потенціалу спокою. Потоки приблизно рівні по величині, але зрушені за часом. Завдяки цьому зрушенню в часі й можлива поява потенціалу дії. Якби потоки натрію й калію через мембрану збігалися в часі, то вони б компенсували один одного, і ніякої зміни мембранного потенціалу не могло б відбуватися.
Узагальнимо вищесказане. Потенціал дії є результатом швидкого й значного збільшення натрієвої провідності мембрани. Вхід великої кількості іонів натрію й акумуляція позитивного заряду на внутрішній поверхні мембрани клітини збуджує мембранний потенціал у напрямку ENa. Реполяризація мембрани відбувається в результаті наступного збільшення калієвої провідності й зменшення внутрішньоклітинного позитивного заряду через вихід іонів калію із клітини. Мембранний потенціал при цьому знову наближається до ЕК.
Яка кількість іонів входить у клітину й виходить з неї під час потенціалу дії
Головною властивістю натрієвої й калієвої провідністю, що визначає характер струмів під час потенціалу дії, є їх потенціал-залежність: імовірність відкриття іонних каналів збільшується з деполяризацією мембранного потенціалу. Деполяризація збільшує натрієву провідність, а також, з деякою затримкою, калієву. Вплив деполяризації на натрієву провідність носить регенеративний характер: спочатку невелика деполяризація збільшує кількість відкритих каналів; іони натрію, що входять у клітину по напрямку свого електрохімічного градієнта, роблять подальшу деполяризацію мембрани, відкриття більшої кількості каналів, що спричиняє вхід ще більшого числа іонів, і так далі. Такий процес, який самопідсилюється, характеризується позитивним зворотним зв'язком. Потенціал-залежність калієвих каналів, навпаки, характеризується наявністю негативного зворотного зв'язку. При деполяризації кількість відкритих калієвих каналів зростає, і іони калію входять у клітину в напрямку електрохімічного градієнта. Однак, вхід калію не підсилює деполяризацію, а веде до реполяризації й повернення калієвої провідності в стан спокою.
Якщо врахувати, що на фазі росту потенціалу дії в клітину входить велика кількість натрію й велика кількість калію виходить із неї на фазі спаду, то стає очевидним, що концентрації цих іонів у цитоплазмі повинні змінитися. Величину цих змін можна визначити або експериментально, або шляхом розрахунків.
При потенціалі - 67 мВ на внутрішній поверхні мембрани перебуває приблизно 4 • 1011 негативних зарядів на см2. На піку потенціалу дії (+40 мВ) замість цього негативного заряду усередині клітини накопичується близько 2,4 • 1011 позитивних зарядів, що відбувається в результаті входу в клітину 6,4 • 1011 іонів натрію на см2. Це відповідає приблизно 10-12 моль/см2. Експериментальні виміри входу радіоактивного натрію й виходу радіоактивного калію дали значення між 3 • 10-12 й 4 • 10-12 моль/см2. Розходження між теоретично передказаним й експериментальним значеннями порозуміваються головним чином тим, що при розрахунках не приймалося в увагу те, що вхід натрію й вихід калію частково перекриваються в часі. Таким чином, кількість вхідного натрію перевищує значення, необхідне для деполяризації мембрани до пікового рівня потенціалу дії, оскільки вихід калію починається до моменту досягнення піка.
Вплив входу натрію на його внутрішньоклітинну концентрацію
Сегмент аксона кальмара довжиною 1 см і діаметром 1 мм має площу поверхні близько 0,31 см2, тому при швидкості входу натрію 3,5 • 10-12 приведе до нагромадження приблизно 10-12 M натрію. Обсяг цього відрізка аксона становить 7,85 • 10-12 л, у ньому втримується (при щільності 50 ммоль/л) 4 • 10-7 M натрію, тому зміна концентрації натрію в результаті потенціалу дії дуже мало й становить 0,0000025. Вихід калію робить порівнянний ефект і змінює внутрішньоклітинну концентрацію калію в 0,000003 разів.
Потенціал дії в більш тонких відростках нейрона може привести до більш значних змін внутрішньоклітинних концентрацій натрію й калію, чим у гігантському аксоні кальмара. Так, нервове закінчення діаметром 1 мкм і довжиною 100 мкм має площу поверхні 3 • 10-6 см2 й обсяг 8 • 10-14 л. Під час потенціалу дії вхід натрію 3 • 10-6 моль/см2 робить збільшення кількості натрію усередині термінали на 10-17 М. При внутрішньоклітинній концентрації 20 мм у термінали втримується приблизно 1,5 • 10-15 М, так що збільшення концентрації натрію в результаті одного потенціалу дії становить 0,7 %. Пачка з 50 імпульсів теоретично приведе до збільшення рівня натрію на 35 % і подібному зниженню рівня калію. Вхід натрію збільшує активність натрій-калієвого обмінника так, що концентрації швидко повертаються до початкового рівня.
Кальцієві потенціали дії
Слід зазначити, що натрієво-калієвий механізм генерування потенціалу дії не є єдиним. У генерації потенціалу дії можуть брати участь й інші катіонні канали. У клітинах водоростей Хаара реверсія мембранного потенціалу обумовлена дифузією в клітину іонів хлору. У деяких клітинах фаза росту потенціалу дії визначається активацією кальцієвих каналів, а реполяризация відбувається завдяки активації різних типів калієвих каналів.
Виявлені так само клітини, у яких реверсія мембранного потенціалу обумовлена дифузією іонів кальцію. До таких, зокрема, відносяться волокна гладеньких м'язів кишечнику, матки, судин й інших порожнистих органів.
У мембрані нервів і м'язових волокон утримується велика кількість потенціалзалежних кальцієвих каналів. Кальцій, що входить у клітину через ці канали під час потенціалу дії, впливає на самі різні процеси. Приміром, короткочасне збільшення рівня кальцію в ході потенціалу дії викликає як секрецію хімічних медіаторів у нервовому закінченні, так і скорочення м'язового волокна.
У деяких м'язових волокнах і нейронах кальцієві струми досягають такої величини, що або вносять значний вклад, або повністю формують фазу росту потенціалу дії. Процес цей носить регенеративний характер завдяки зростанню дСа при деполяризації, точно такому, як у натрієвих каналів. Участь кальцієвих каналів у потенціалі дії було вперше вивчене Фаттом і Гинзборгом, а згодом Хагиварой. Кальцієві потенціали дії описані в серцевому м'язі, у цілому ряді нейронів безхребетних, а також у нейронах вегетативної й центральної нервової системи хребетних. Наявність кальцієвих потенціалів дії також показано в ненейрональних типах клітин, таких як ряд ендокринних клітин і деяких яйцеклітин безхребетних. Потенціалзалежні кальцієві струми блокуються в мілімолярних концентраціях кобальтом, магнієм або кадмієм, доданим у позаклітинний розчин. Барій може замінити кальцій у проходженні через пору каналу; магній на це не здатен. Разючим прикладом співіснування натрієвих і кальцієвих потенціалів дії в одному типі клітин є клітина Пуркіньє в мозочку ссавців. Натрієві потенціали дії генеруються в тілі клітини Пуркіньє, у той час як кальцієві - у дендритах.
Іони кальцію впливають також на збудливість мембрани: зниження позаклітинної концентрації кальцію приводить до збільшення збудливості; підвищення позаклітинного рівня кальцію, навпроти, спричиняє зниження збудливості.
Поширення потенціалу дії
Нервові та м’язові волокна являють собою циліндричні провідники. Їхній внутрішній вміст, що має відносно низький питимий опір, ізольовано мембраною від зовнішнього добре провідного середовища. Поширення ПД відбувається завдяки виникненню локальних струмів між збудженою та не збудженою ділянками нервового або м’язового волокна.
Що більший внутрішній опір кабельної структури, її ємність і опір зовнішнього середовища, то менша швидкість проведення. Швидкість поширення ПД пропорційна кореню квадратному від діаметра волокна.
У мієлінізованих нервових волокнах потенціал дії виникає ("генерується") у перехопленнях Ренвьє, а потім передається від одного перехоплення до іншого чисто електричним шляхом. У безмієлінових нервових волокнах кожна ділянка волокна, сприймаючи електричний сигнал від сусідніх ділянок нерва, генерує потенціал дії, що потім поширюється далі.
Безперервне поширення нервового імпульсу
У стані спокою вся внутрішня поверхня мембрани нервового волокна несе негативний заряд, а зовнішня сторона мембрани - позитивний. Електричний струм між внутрішньою й зовнішньою стороною мембрани не протікає, тому що ліпідна мембрана має високий електричний опір. Під час розвитку ПД, тобто при відкритті натрієвих каналів, у збудженій ділянці мембрани відбувається реверсія заряду. На межі збудженої й не збудженої ділянки починає протікати електричний струм (рух іонів Na+).
Усередині нервового волокна виникає струм від позитивного полюса до негативного полюса, тобто струм спрямований від збудженої ділянки до не збудженої. Електричний струм подразнює найближчу ділянку мембрани, деполяризує його й приводить його в стан збудження, у той час як раніше збуджені ділянки повертаються в стан спокою через стадію рефрактерності. У зв'язку з наявністю рефрактерності, зворотний хід хвилі виявляється неможливим. Таким чином, хвиля збудження електротонічно (пасивно) охоплює все нові ділянки мембрани нервового волокна. При безперервному проведенні поверхня має електрогенні властивості по всієї довжині. Тому малі кругові струми виникають на відстані в кілька мікрометрів. Збудження має вигляд хвилі, що постійно рухається. Розмір ділянки мембрани, що перебуває під впливом ПД, залежить від його тривалості й швидкості проведення. Наприклад, якщо тривалість ПД дорівнює 2 мс, а швидкість його проведення 10 м/с, то потенціал пошириться на ділянці мембрани 2 см.
Сальтаторне поширення нервового імпульсу
У мієлінізованому нервовому волокні ділянки мембрани, покриті мієліновою оболонкою, є незбудливими; збудження може виникати тільки в ділянках мембрани, розташованих в області перехоплень Ренвьє, де перебуває максимальна кількість керованих потенціалчуттєвих натрієвих каналів - 12000 на 1мкм. Час проведення порушення по мієліновому волокну назад пропорційно довжині між перехопленнями.
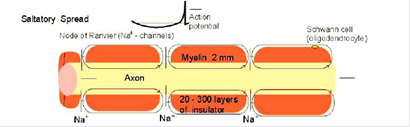
Поширення потенціалу по немієлізованому й мієлінізованому волокнах.
При розвитку ПД в одному з перехоплень Ренвьє відбувається реверсія заряду мембрани. Між електронегативними й електропозитивними ділянками мембрани виникає електричний струм, що подразнює сусідні ділянки мембрани. Однак стан порушення може перейти тільки на ділянку мембрани в області наступного перехоплення Ренвьєе. Таким чином, порушення поширюється по мембрані стрибкоподібно (сальтаторно) від одного перехоплення Ренвьє до іншого. "Перестрибування" ПД через ділянку між перехопленнями можливо тому, що амплітуда ПД в 5-6 разів перевищує граничну величину, необхідну для збудження середнього перехоплення. ПД може "перестрибувати" навіть через два міжперехватних проміжки. Сальтаторне проведення порушення є високошвидкісним, высокоекономічним.
Швидкість поширення ПД у фазних м’язових волокнах набагато менша (1-5 м/с), ніж у нервових волокнах такого ж діаметру. Це пов’язане в першу чергу з більшою вхідною ємністю мембрани, зумовленою тубуляторною системою м’язового волокна.
Швидкість проведення збудження в гладеньких м’язах невелика та складає декілька см/с.
Фіксація потенціалу
Кількісний опис механізмів, що беруть участь у генерації потенціалу дії, стало можливим завдяки методу виміру мембранних струмів в умові фіксації потенціалу. Цей метод дозволяє визначити, який внесок вносять іони того або іншого типу в мембранний струм, а також обчислити величину й часовий хід змін відповідних іонних провідність.
Винахід методу фіксації потенціалу
Той факт, що фіксація трансмембранного потенціалу дозволить вимірювати мембранну провідність по змінах струму при постійній напрузі, був уперше усвідомлений ще в 30-х рр. XX століття, і тоді ж англійські дослідники Алан Ходжкін й Ендрю Хакслі (Alan Hodgkіn and Andrew Huxley) почали експерименти із двухелектродною фіксацією потенціалу (2-electrode voltage clamp).
Суть методу полягає в наступному. У клітину вводяться два електроди, ще один - електрод порівняння - залишається поза клітиною. Перший внутрішньоклітинний електрод служить для виміру трансмембранної різниці потенціалів (тобто різниці потенціалів між ним й електродом порівняння), другий може подавати струм. Спеціальний пристрій - генератор сигналу - задає командний потенціал, якому повинен бути рівен трансмембранний потенціал. Обмірюваний трансмембранний потенціал подається на вхід пристрою порівняння, що віднімає обмірюваний потенціал з командного й, залежно від величини різниці, подає струм на струмовий електрод так, щоб компенсувати цю різницю. Монітор струму, у свою чергу, постійно вимірює величину струму, що для цього необхідна. В 1930-х й 40-х роках, коли працювали Ходжкін і Хакслі, не існувало мікроелектродів, тому як внутрішньоклітинні електроди використалися тонкі дроти. Це визначило вибір об'єкта - гігантський аксон кальмара діаметром 1 мм, усередину якого й вводилися ці дроти. На цьому об'єкті методом двохелектродної фіксації потенціалу дослідники виконали експерименти, у яких була встановлена іонна природа потенціалу дії й уперше постульоване існування іонних каналів (Нобелівська премія 1963 р., поділена із Дж. Екклзом, що получили її за дослідження в області синапатичної передачі). Двохелектродна фіксація потенціалу застосовується й у цей час, з використанням гострих скляних мікроелектродів, однак навіть із ними ця методика має істотні обмеження: по-перше, два електроди можуть бути уведені тільки в досить велику клітину (наприклад, овоцит жаби).
Дослідження провідності одиночних калієвих і натрієвих каналів під час потенціалу дії проводилися в умовах фіксації потенціалу ділянки мембрани. Спостережувані при цьому принципи роботи окремих каналів відповідають результатам, отриманим раніше в експериментах з фіксацією потенціалу цілої клітки: при деполяризації ймовірність відкриття натрієвих і калієвих каналів зростає. Зростання ймовірності відбувається з тим же тимчасовим ходом, що й відповідні струми в умовах фіксації потенціалу. Так, натрієві канали найбільше часто відкриваються на початку деполяризуючого імпульсу й імовірність таких відкриттів падає в міру розвитку натрієвої інактивації.
Первісний короткочасний викид струму являє собою ємнісний струм, обумовлений зміною заряду на мембрані в результаті зміни мембранного потенціалу. Якщо підсилювач зворотного зв'язку здатний проводити більші струми, то ємнісний струм триває дуже недовго. На практиці викид ємнісного струму триває близько 20 мс, і за ним треба невеликий, але стійкий вихідний струм.
Цей вихідний струм, що протікає через провідності, активні при потенціалі спокою, називається струмом витоку. Здебільшого це струм іонів калію й натрію, що має лінійну залежність від величини зсуву потенціалу фіксації від потенціалу спокою й з на всьому протязі стрибка потенціалу. Більшу частину часу, однак, цей струм замаскований іншими, набагато більшими по величині іонними струмами.
Ходжкін і Хакслі показали, що друга й третя стадії струму обумовлені спочатку входом іонів натрію, а потім виходом іонів із клітини. Їм також удалося виділити індивідуальні компоненти струму й розрахувати їхню величину й часовий хід. Одним із зручних способів домогтися цього послужило видалення з розчину більшої частини іонів натрію й заміна їх на іони холіну (які не проходять через мембрану). Знизивши зміст позаклітинного натрію, удалося домогтися того, що натрієвий рівноважний потенціал зрівнявся з деполяризованим мембранним потенціалом. Таким чином, сумарний струм зрівнявся нулю.
Метод петч-кламп (patch - clamp)
Наприкінці сімдесятих років XX в. Эрвін Неєр (E.Neher) і Берт Сакман (B.Sakmann) виявили, що конусоподібні скляні піпетки з діаметром кінчика 1-2 мікрона можуть утворювати контакти із клітинною мембраною (границі "мембрана - скло") з опором у декілька гігаом - це так званий гігаомний контакт. Він дозволяє ізолювати від зовнішнього середовища й від іншої частини мембрани той її фрагмент, що перебуває усередині піпетки. Відмежований піпеткою фрагмент мембрани й називається patch - "фрагмент", слово clamp (фіксація) у назві методу можна інтерпретувати і як захоплення й ізоляцію цього фрагмента, так й як фіксацію трансмембранного потенціалу в ізольованому фрагменті, або, як буде описано пізніше, потенціалу або струму на цілій клітині. У піпетку, заповнену розчином електроліту, міститься хлор-срібний електрод, другий електрод розміщується позаклітинно, в оточуючій рідини. Відмінність електричної схеми від раніше описаної для двохелектродної фіксації полягає в тім що той самий електрод використовується як для виміру різниці потенціалів, так і для подачі струму.
Величезна сфера, частина якої видна на моніторі в центрі фотографії - це клітина (овцит жаби Xenopus laevіs), що перебуває в цей момент на предметному столику мікроскопа, до нього підведена patch-піпетка, її діаметр у носика - близько 3 мікронів. Після встановлення гігаомного контакту вона ізолює фрагмент мембрани площею приблизно 7 мкм2, струм від іонних каналів у цьому фрагменті можна буде записувати.
Конфігурація "Cell-attached"
Тільки що описана конфігурація називається cell-attached patch-clamp (patch-clamp "із прикріпленою кліткою"). Вона ілюструється схемою 1.
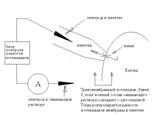
Схема 1. Принципова схема patch-clamp у конфігурації Cell-attached.
Ця конфігурація, однак, має дві незручності. По-перше, вона не дозволяє з достатньою надійністю вимірювати, а, отже, і задавати трансмембранну різницю потенціалів - оскільки обидва електроди - як піпетковий, так і зовнішній, перебувають по одну сторону мембрани. Загалом кажучи, можна, використовуючи оточуючий розчин з іонною композицією, що повторює склад цитоплазми, деполяризувати мембрану поза піпеткою, так що різниця потенціалів між зовнішнім електродом і цитоплазмою зникне, а тоді різниця потенціалів між електродами буде дорівнювати трансмембранному потенціалу - але все це досить приблизно, тому що точний склад цитоплазми нам невідомий.
По-друге, ця конфігурація не дозволяє контролювати склад середовища поза піпеткою - там залишається цитоплазма, склад якої не цілком визначений. У силу цих причин cell-attached mode застосовується досить обмежено.
Конфігурація Іnsіde-out
Однак, якщо піпетку швидким рухом відвести від клітини, то "внутрішній" шматочок мембрани відірветься від клітини й вийде конфігурація іnsіde-out (зовнішня сторона усередині), названа так тому, що внутрішня, звичайно звернена до цитоплазми, сторона мембрани виявиться зовні - в оточуючому розчині, а зовнішня - усередині піпетки. (Схема 2) Альтернативний метод переходу в конфігурацію іnsіde-out полягає в наступному: з конфігурації cell-attached піпетку відводять плавно, формуючи із двох сторін закриту везикулу; потім піднімають піпетку в повітря й опускають у другу ванночку, із внутрішньоклітинним розчином. При переносі зовнішня мембрана везикули руйнується, у результаті формується конфігурація іnsіde-out.
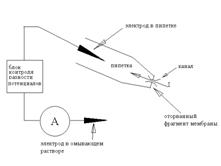
Схема 2. Принципова схема patch-clamp у конфігурації іnsіde-out.
Тепер різниця потенціалів на фрагменті мембрани строго дорівнює різниці потенціалів між електродами. Очевидно, що при використанні такої модифікації піпетку заповнюють розчином, що імітує позаклітинне середовище, тоді як оточуючий розчин роблять близьким по складу до цитоплазми. При цьому, міняючи склад оточуючого розчину, можна вивчати, як такі зміни в цитоплазмі впливають на струм каналів, що цікавлять нас, - адже оточуючий розчин контактує із цитоплазматичною стороною мембрани.
Конфігурація Whole-cell
Якщо за умовами експерименту необхідно міняти склад позаклітинного середовища, можна використати конфігурацію whole cell ("англ. ціла клітина"). У цьому випадку піпетку не відводять від клітини, а подають у неї негативний тиск й у такий спосіб руйнують ізольований фрагмент мембрани.
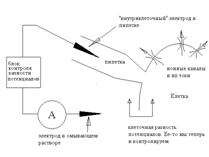
Схема 3. Принципова схема patch-clamp у конфігурації whole-cell.
Після цього піпетка з'єднана із внутрішньоклітинним середовищем; оскільки клітина звичайно маленька, то, завдяки дифузії, склад цитоплазми незабаром виявляється ідентичним складу піпеткового розчину, тому ми, як й у попередньому випадку, знаємо як склад рідин, так і різницю потенціалів. Відзначимо, що якщо в конфігурації іnsіde-out піпетковий електрод був "позаклітинним", а внутрішній - "внутрішньоклітинним", те тепер вони помінялися ролями. З погляду вивчення струму через канали, перевага цього методу полягає в тому, що тут залишаються цілими клітинні структури й регуляторні механізми. Але є й недолік - ми вимірюємо сумарний струм всіх каналів у клітині, а не струми через одиночні канали.
Конфігурація Outsіde-out
Вирішити цю проблему дозволяє конфігурація outsіde-out (зовнішня сторона зовні). Якщо після переходу в whole-cell mode повільно відводити піпетку від клітини, мембрана не відривається відразу, а починає витягуватися в трубку (це видно на фото).

Початкова фаза переходу з Whole-cell в Outsіde-out. Перехід в outsіde-out. Заключна фаза.
Мембранна трубка стала зовсім тонкою й майже невидимою ("протуберанець" у поверхні клітини - це невелика частина цитоплазми, що залишилася в мембранній трубці). У наступну мить трубка порветься, а мембрана зімкнеться на піпетці в "вивернутому" виді - ми опинемось в конфігурації "outsіde-out"
Perforated patch
Perforated patch ("перфорований patch") - це специфічний варіант patch-clamp в whole-cell mode. У цьому випадку, у піпетковому розчині утримується невелика кількість спеціального антибіотика. наприклад, граміцидина. Антибіотики цього класу формують іонні канали в клітинній мембрані на ділянці, приєднаній до мікропіпетки.
Такий підхід дозволяє уникнути заміщення внутрішнього середовища клітини розчином з піпетки-електрода, тобто клітина залишається живою з мінімальними, наскільки це можливо, ушкодженнями. Таким чином, відповіді клітки на подразники є максимально наближеними до природного. У той же час, даному методу властиве ряд недоліків. По-перше, у порівнянні із класичним whole-cell mode, електричний опір доступу (яке складається з опору піпетки й опору в місці з'єднання піпетки з мембраною) є значно більш високим. Це знижує якість фіксації потенціалу, підвищує рівень шуму при записі, і збільшує значення всіх помилок, пов'язаних зі змінами опору повного ланцюга (від електрода в зовнішньому розчині до електрода в піпетці). По-друге, для того, щоб антибіотик подіяв, потрібно досить багато часу (до 30 хвилин), що істотно зменшує корисний період експерименту. І, по-третє, антибіотик ушкоджує мембрану також й у місці з'єднання з кінчиком піпетки, що приводить до прискореного руйнування гігаомного контакту й додатково зменшує ефективний експериментальний час. Таким чином, даний варіант методу може бути з успіхом використаний тільки в експериментах, які не вимагають тривалого часу для виявлення досліджуваних явищ.
Nucleated patch
Цікавий різновид patch-clamp - nucleated patch. (Фіксація потенціалу із клітинним ядром).
Метод полягає в наступному. Піпетка підводить до клітини, а потім ривком пробиває мембрану. Після цього кінчик піпетки підводить до клітинного ядра, на піпетку подається невеликий негативний тиск. У результаті піпетка присмоктує до ядра. Потім піпетка з ядром на кінці плавно приділяється назад і виймається із клітини. Піпетку необхідно вивести із клітини таким чином, щоб ділянка мембрани в місці виходу "наділася" на ядро, що присмокталося, і відірвалася, обернувшись довкола нього. У результаті виходить специфічний варіант outsіde-out patch, при якому набагато більший, ніж у звичайному варіанті методу, ділянка мембрани приєднана до кінця піпетки, будучи обгорнутою навколо клітинного ядра.
Фіксація потенціалу й фіксація струму
Коли вивчаються зміни провідності однотипних каналів у відповідь на якісь хімічні впливи або залежність їхньої провідності від різниці потенціалів на мембрані, зручно фіксувати потенціал і вимірювати струм каналу - як ми дотепер й описували. Цей, найчастіший варіант patch-clamp, називається voltage clamp (фіксація потенціалу). Однак, іноді дослідника цікавлять процеси трансмембранного переносу іонів, пов'язані зі зміною мембранного потенціалу - наприклад, проведення нервового імпульсу. У таких випадках можна надійти протилежним образом: зафіксувати на постійному рівні струм, і вивчати зміни різниці потенціалів при цьому. Такий варіант називається current clamp (фіксація струму).
Приклади записів, одержуваних методом patch-clamp

Запис струму одиночного каналу (гліцинового рецептора)
Запис одиночного каналу рецептора гліцину. Чітко видно два стани: закрите (йому відповідає нульовий струм) і відкрите (йому відповідає струм приблизно в 7 пикоампер). Канал час від часу спонтанно переходить із одного стану в інше, проміжних станів немає. Запис дозволяє одержати дві найважливіші характеристики каналу: провідність (виходячи з величин трансмембранного потенціалу й струму) і ймовірність знаходження у відкритому стані (обумовлене як відношення часу, коли канал відкритий до часу, коли він закритий). До слова, найчастіше активність каналу фізіологічно регулюється шляхом зміни цієї ймовірності.
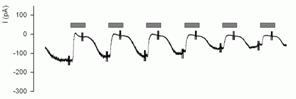
Запис амилорид-чуттєвого струму в головній клітині збірної трубки бруньки в конфігурації whole-cell.
Запис у конфігурації whole-cell. Клітина епітелію збірної трубки. Трансмембранний потенціал -60 мВ. З інтервалами в 1 хвилину на 30 секунд в оточуючий розчин уводиться амилорид - інгібітор епітеліального натрієвого каналу (час введення показаний темними прямокутниками). До речі, чутливість до інгібіторів - це ще одна з найважливіших характеристик каналу. Введення амилорида щораз призводить до падіння струму до нуля, із чого видно, що практично вся провідність при -60 мВ у даній клітині - це провідність епітеліального натрієвого каналу. На противагу попереднього запису, зміни струму виглядають безперервними - це пов'язане з тим, що одночасно записується багато каналів (близько 2000), відповідно, є приблизно 2000 рівнів струму - від "усі відкриті" до "усі закриті".
Висновки
· Потенціал дії в більшості клітин виникає за рахунок короткочасного зростання натрієвої провідності, що прагне привести мембранний потенціал до рівня натрієвого рівноважного потенціалу й за яким треба збільшення калієвої провідності, що повертає мембрану в стан спокою.
· Зростання провідностей виникає завдяки потенціалзалежності натрієвих і калієвих каналів: імовірність їхнього відкриття зростає при деполяризації.
· Експерименти на аксоні кальмара з фіксацією потенціалу надали детальну інформацію про потенціалзалежності й тимчасовий хід змін провідностей. При деполяризації мембрани натрієва провідність спочатку швидко активується, а потім інактивується. Калієва провідність активується із затримкою й залишається на високому рівні доти, поки не скінчиться деполяризація.
· Часовий хід і потенціалзалежність змін натрієвих і калієвих провідностей у точності визначають амплітуду й часовий хід потенціалу дії, а також такі характеристики мембрани, як поріг активації й рефрактерний період.
· Теоретично активація натрієвої й калієвої провідностей при деполяризації повинна бути зв'язана зі зсувом заряду усередині клітини. Ці зсуви, названі воротними струмами, удалося виміряти експериментально.
· Експерименти з використанням методу пэтч-кламп доповнили відомості, отримані в більше ранніх досвідах з фіксацією потенціалу, новими деталями про процес збудження. Так, наприклад, натрієві канали відкриваються на досить короткий час, і ймовірність їхнього відкриття в ході деполяризації спочатку зростає, а потім знижується, відповідно до активації й інактивацією натрієвої провідності в цілій клітині. Різні кінетичні моделі були запропоновані з метою опису процесів активації й інактивації каналів.
· Кальцій відіграє важливу роль у збудженні. У деяких клітинах саме вхід кальцію, а не натрію, відповідає за фазу росту потенціалу дії. Крім того, позаклітинний рівень кальцію визначає збудливість мембрани. Зниження позаклітинної концентрації кальцію призводить до збільшення збудливості.
Література
1. Биофизика: Учеб. для биол. спец. унив. Под ред. Б.Н. Торусова и О.Р. Кольс. — М.:Высш.шк., 1968. — 467 стр.
2. Валькенштейн М.В. Биофизика: Учеб. пособие для биолог. и физиол. факультетов университетов. — М.: Наука, 1981. — 575 стр.
3. П.Г. Костюк, В.Л. Зима, І.С. Магура, М.С. Мірошниченко, М.Ф. Шуба Біофізика / За редакцією П.Г. Костюка. — Київ: «Обереги», 2001. — 544 стр.
4. П.Г. Костюк. Микроэлектродная техника. — К.: Изд-во АН УССР, 1960. — 244 стр.
5. Регистрация одиночных каналов / Под ред. Б.Сакмана, Э.Неера. — М.: «Мир», 1987. — 275 стр.
6. Рубин А.Б. Биофизика: Учеб. пособие для биолог. и физиол. факультетов университетов: В 2-х книгах. Кн.1. Теоретическая биофизика. — М.: Высшая школа, 1987. — 319 стр.
7. Рубин А.Б. Биофизика: Учеб. пособие для биолог. и физиол. факультетов университетов: В 2-х книгах. Кн.2. Биофизика клеточных процессов. — М.: Высшая школа, 1987. — 303 стр.
8. B. Sakmann, E. Neher Single-channel recording. — 1995.
9. Numberger M., Draguhn A. Patch-Clamp Technik. — 1996.
10. Jan C. Behrends & Niels Fertig: Planar Patch Clamping. In: Neuromethods. Bd. 38, S. 411—433.
Похожие работы
... ість і індивідуальність, що є мірою виявлення людини - її творчого потенціалу. Для отримання об'єктивних і точних даних про закономірності, механізми психіки і психологічні факти використовуються методи дослідження. Метод - шлях пізнання і застосування системи прийомів для досягнення певної мети. У психології розроблені методи вивчення психічних фактів і феноменів. Методи психології доцільно ...

... хронічної гіперімунокомплексемії, проаналізувала дані експериментальних досліджень та підготувала тези до друку). АНОТАЦІЯ Садляк О. В. Лімфоцитопосередковані механізми за умов хронічної гіперімунокомплексемії та вплив на них корвітину в експерименті. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю 14.03.04 – патологічна фізіологія. – ...
... і відношення в класі виникають на основі другорядних, часто педагогічне небезпечних форм спілкування. Отже оптимальна організація педагогічного спілкування в навчальному процесі - актуальна проблема сучасної школи. За визначенням О.О. Леонтьєва "... оптимальне педагогічне спілкування - це таке спілкування вчителя із школярами в процесі навчання, яке створює найкращі умови для розвитку мотивації ...
... країнами, які цей ліміт використовують. Уряди країн реалізують керівні принципи через свої національні стратегії сталого розвитку. 2. КЛАСИФІКАЦІЯ СИСТЕМ І МЕХАНІЗМІВ ЕКОЛОГІЧНОГО УПРАВЛІННЯ Екологічне управління, як і сама екологія, є досить розгалуженою і багатофункціональною сферою діяльності різних суспільних, державних, корпоративних та інших інституцій, у тому числі міжнародних. Воно ...
0 комментариев