Как в клетке возникает разность потенциалов
Возможно, в результате нашего рассказа создается впечатление, что Дюбуа-Реймон был «чистым» экспериментатором. Но это совершенно неверно. Он имел, как уже говорилось, научную позицию: «Нельзя приписать частицам материи в организме каких-либо новых сил, которые бы не действовали и вне организма». Исходя из этой позиции, он и предложил первое теоретическое объяснение потенциала повреждения: электрические явления в живых организмах возникают потому, что вдоль мышц и нервов якобы тянутся цепочки особых «электромоторных» молекул. Каждая такая молекула представляет собой как бы два гальванических элемента, соединенных положительными полюсами, так что наружу выходят только отрицательные полюса. Где бы ни разрезать мышцу, на разрезе обнажатся отрицательные полюса, чем и объясняется потенциал повреждения.
Здесь четко видно, как биологическая теория строится на основании аналогии с современной ей физической теорией: последним словом о магнетизме была тогда теория Ампера о том, что свойства постоянных магнитов объясняются тем, что каждая молекула является маленьким магнитиком.
Дюбуа-Реймон придумал, как теперь сказали бы, модельный эксперимент для проверки своей гипотезы. Он напаял много маленьких элементиков «медь — цинк», соединил их попарно положительными полюсами, укрепил на деревянной доске и, погрузив всю систему в раствор соли, стал проводить на этой «искусственной мышце» такие же эксперименты, которые он проводил на мышце живой. Обнаружилось, что распределение токов в такой модели действительно было сходно с распределением токов у реальной мышцы.
Благодаря такой оригинальной разработке, а отчасти и авторитету Дюбуа-Реймона теория электромоторных молекул, несмотря на ее фантастичность, была общепризнанной почти четверть века с момента ее выдвижения молодым Дюбуа в 1846 г.
Но вот подросли ученики Дюбуа-Реймона — они-то и оказались «подрывателями устоев». Все началось с Людвига Германа, в котором Дюбуа-Реймон воспитал уважение к техническим деталям, тщательность и точность в экспериментах. Дело в том, что по модели Дюбуа-Реймона в нормальной целой мышце тоже должны идти токи, так как в области сухожилий торчат ничем не скомпенсированные «самые последние в ряду» отрицательные концы электромоторных молекул. Дюбуа-Реймон обнаруживал такие токи, не делая специального разреза. Но когда аккуратный Герман научился так препаровать мышцу, чтобы действительно не повредить ее, оказалось, что в таких условиях никакие токи не возникают.
Значит, рассуждал Герман, в нормальной целой мышце и нерве вообще нет никаких токов и потенциалов. Токи возникают только тогда, когда мышцу или возбуждают, или повреждают. И причиной этого, предположил Герман, являются химические реакции, которые начинают идти на границе нетронутой и поврежденной протоплазмы.
Возражения Германа не смутили Дюбуа-Реймона: ну, и что же, значит, в конце мышцы есть другие молекулы — положительные, которые экранируют отрицательные полюса. Но все равно внутри мышцы токи есть всегда, а повреждение нужно только, чтобы их обнаружить.
На этом и сосредоточился второй «великий спор» в электробиологии: возникает ли электричество в мышце только в момент повреждения или существует до повреждения.
Одним из самых весомых аргументов в научных спорах является эксперимент, однако бывает и так, что одна группа ученых придумывает красивый опыт, вполне убедительно его истолковывает, и все считают, что она победила, но потом выясняется, что опыт может иметь и совсем другое истолкование.
Спор двух гипотез о возникновении электричества в живых тканях в этом отношении очень интересен: если подумать, можно прийти к заключению, что разрешение этого спора прямым экспериментом невозможно. Ведь какой бы опыт для измерения разности потенциалов между наружной и внутренней частями мышцы ни поставить, неминуемо придется проникать в эту внутреннюю область, а тогда сторонники гипотезы альтерации скажут, что нанесено повреждение. По той же причине сторонники гипотезы альтерации не могут доказать, что в нетронутой мышце не существует разности потенциалов между ее внутренними и наружными частями.
Однако это соображение, по-видимому, не пришло в голову участникам дискуссии, и для подтверждения той или иной гипотезы ставились многочисленные новые опыты, которые хотя и не могли переубедить противников, но существенно способствовали развитию электрофизиологии.
Например, Герман поставил такой опыт. Он очень быстро разрезал мышцу и показал, что потенциал регистрируется не сразу после разреза, а спустя примерно 0,02 с. Это обстоятельство он объяснил тем, что требуется время для развития необходимых химических реакций. Но даже если бы опыт был верен, он не опроверг бы гипотезу предсуществования: Дюбуа-Реймон считал, что при возбуждении молекулярные батарейки выключаются и потом через некоторое время снова включаются; то же самое может происходить и при повреждении.
Однако в 1904 г, Бернштейн, который к этому времени стал научным противником Германа, и его сотрудник Чермак, повторив опыт Германа, показали, что уже через 0,3 мс после разреза можно зарегистрировать разность потенциалов. Но это не возымело никаких последствий. Герман просто решил, что реакции в результате повреждения развиваются быстрее, чем он первоначально думал.
Так можно ли вообще решить, кто прав в этом споре? Конечно, можно. Ведь уверен, например, каждый из нас, что семечки внутри арбуза существуют там до того, как мы его разрезали, а не появляются в момент разрезания и в результате разрезания,
А, собственно, почему? Почему мы так уверены в существовании семечек внутри арбуза, хотя никак не можем это проверить, не разрезая его? Такую уверенность нам дает понимание того, как возникают эти семечки. Точно так же и в случае с биопотенциалами: для того чтобы решить спор Дюбуа-Реймона и Германа, надо было объяснить механизм возникновения этого потенциала или до повреждения, или в результате повреждения. При этом следовало строить это объяснение так, чтобы оно не противоречило другим научным данным и вписывалось в общую картину. То есть основное содержание спора было не в том, когда возникает потенциал, а как он возникает.
Творцом теории биопотенциалов стал ученик Дюбуа-Реймона Юлиус Бернштейн.
А различные идеи, факты и открытия, на которых эта теория основывается, накапливались в науке очень долго.
Отступление о детективах. Научное исследование часто похоже на детектив. Вернее, не на литературный детектив, а на реальное настоящее следствие. Чтобы выяснить истину: как все произошло и кто виновник, а главное — собрать доказательства, приходится проделывать огромную работу. Здесь все важно: и сбор свидетельских показаний, и уточнение отдельных фактов вплоть до мельчайших деталей. При этом подавляющее большинство данных оказываются не имеющими отношения к делу, а тем не менее их нужно собирать — ведь заранее это не известно. И самое трудное, вероятно, выделить в этом наборе сведений ту систему, которая приведет к истине и позволит потом выстроить стройную цепочку доказательств.
А вот когда следствие описывается в книге, то автору уже известна разгадка. Он тогда знает, какие детали существенны, а какие нет; но разные авторы используют эти детали по-разному. Одни авторы включают в ткань повествования все данные, необходимые для получения разгадки. Но читатель, как правило, их не замечает или не придает им значения. И только потом, когда Шерлок Холмс рассказывает доктору Ватсону, как он нашел разгадку, читатель понимает, почему было важно, что на ужин был чесночный соус, а собака не лаяла. Читателю иногда даже становится досадно, что он сам не догадался обо всем.
Очень редко детектив начинается с рассказа о разгадке.
Историю же научных открытий, как правило, пишут именно так, сразу сообщая ответ. Мы же старались включить в свое изложение, по крайней мере, те факты и идеи, которые были важны для научного следствия, не имея возможности рассмотреть многочисленные ложные версии, возникавшие по ходу дела.
Сейчас наступило время выстроить из, казалось бы, иногда совсем не относящихся к делу сведений ту цепочку, которая покажет, кто виновник «преступления», т. е. «животного электричества», и каким оружием «преступник» пользовался».
От осмоса к электричеству
В самом начале нашего рассказа мы обмолвились вскользь, что аббат Нолле знаменит открытием осмоса. В то время никому и в голову не могли прийти, что именно исследование этого явления будет первым шагом к разгадке тайны «животного электричества». Это случилось так.
В 1826 г. парижский врач и физиолог А. Дютроше заинтересовался таким замечательным свойством растения, как корневое давление, которое гонит сок из срезанного ствола. Если заключить ствол растения в трубку, то сок поднимается по ней на заметную высоту.
И сейчас нам кажется удивительным, как это шампиньон своей мягкой шляпкой пробивает асфальт, а набухший в трюме горох может разорвать пароход. А тогда это казалось чудом: откуда у слабых корней растений такая сила, что они ломают камни? Недаром А. Н. Толстой в своем фантастическом романе «Аэлита» придумал, что атланты, переселившиеся на Марс, использовали для двигателей своих космических кораблей «растительную силу семян».
Дютроше подумал: а нельзя ли все эти чудеса объяснить осмосом — явлением, открытым еще в 1748 г. аббатом Нолле?
Напомним, что осмос — это явление самопроизвольного перехода растворителя в раствор, отделенный от него перепонкой, через которую проходит растворитель, но не проходит растворенное вещество. Такие пленки называют полупроницаемыми. Полупроницаемостью обладают разные пленки биологического происхождения: кожа лягушки, стенка мочевого пузыря и т. д. Если взять стеклянную трубку и затянуть один ее конец полупроницаемым материалом, затем налить в трубку раствор сахара и опустить ее затянутым концом в воду, то вода будет проникать из наружного сосуда в трубку и уровень раствора в трубке поднимется выше, чем в наружном сосуде, создав так называемое осмотическое давление.
Дютроше предположил, что это-то давление и является причиной замечательных свойств растений. Однако знаменательно, что само явление осмоса Дютроше считал проявлением «жизненной силы». Он попробовал заменить бычий пузырь или другие ткани биологического происхождения сосудом из пористой глины, полагая, что такой опыт безнадежен, ведь «жизненной силы» в глине нет. Но глиняный сосуд работал ничуть не хуже! Заслуживает глубокого уважения научное мужество Дютроше, который смог полностью пересмотреть свои научные воззрения. С той поры он, по его словам, «...навсегда соединил физику с физиологией». Дютроше много сделал для объяснения поглощения воды из почвы, подъема воды вверх по стволу, движений листьев мимозы и др. Все эти явления он пытался объяснить осмосом.
Но самое главное, Дютроше показал, что осмос — это чисто физическое явление. С этого момента сторонники физико-химического направления в биологии начали многочисленные исследования роли осмоса в организме, в частности в организме животных. Осмосом начали объяснять всасывание пищи в кишечнике, а К. Людвиг пытался объяснить на основе осмоса работу почек. До сих пор корневое давление объясняется на основе осмоса в принципе так же, как это было сделано в 60-х годах прошлого столетия немецким физиологом растений Ю. Саксом.
Основную роль в исследовании осмоса сыграли в это время работы ботаников. Именно ботаники первыми начали изучать живое, как теперь говорят, на клеточном уровне. И это естественно, потому что впервые клетки были обнаружены именно у растений: они часто более крупные, чем животные клетки, и, главное, отделены друг от друга четко видимой под микроскопом перегородкой.
Главная задача состояла в том, чтобы выяснить, какие вещества и как быстро могут проникать в клетку. Представим себе, что растительную клетку поместили в концентрированный раствор какого-то вещества. Если это вещество не проникает в клетку, то за счет осмоса вода начнет выходить из клетки наружу и клетка должна уменьшить объем, сжаться. Но изучение осмоса показало, что видимая в микроскоп клеточная оболочка вовсе не сжимается, она ведет себя как прочный жесткий каркас, а вот объем внутреннего содержимого клетки меняется по законам осмоса. Отсюда немецким ботаником В. Пфеффером был сделан вывод, имеющий чрезвычайно важное значение для развития биологии. Он предположил, что на поверхности растительной клетки под «панцирем» имеется еще одна, невидимая в микроскоп оболочка — клеточная мембрана, которая и играет на самом деле роль полупроницаемой оболочки.
Пфеффер сделал также следующий принципиальный шаг в изучении осмоса — он измерил осмотическое давление. Для этого Пфеффер воспользовался искусственными полупроницаемыми мембранами. Эти непрочные пленки он наносил на пористый глиняный сосуд, который не давал им лопаться от избыточного осмотического давления. Присоединив к такому сосуду ртутный манометр, Пфеффер получил прибор для количественного измерения осмотического давления — осмометр. Измеряя это давление для разных растворов, он обнаружил, что для каждого раствора оно прямо пропорционально концентрации растворенного вещества, не проходящего через мембрану. Но почему для разных растворов при одной и той же концентрации получаются разные давления, ботаник Пфеффер догадаться не сумел.
Мы уже говорили с вами о тесной взаимосвязи разных наук. В большинстве случаев, о которых речь шла выше, физика помогала биологии, подготавливала ее развитие. Однако, как говорится, долг платежом красен. Без работ Гальвани не был бы так быстро открыт электрический ток, без открытия ботаника Броуна медленнее развивалась бы молекулярно-кинетическая теория, а без работ ботаника Пфеффера... Но об этом мы сейчас и расскажем.
В 70-х годах прошлого века молодой голландский ботаник X. Де Фриз занимается влиянием осмоса на изменение объема клеток растений в растворах разной концентрации.
В 1894 г. Де Фриз рассказал о работах Пфеффера по осмотическому давлению молодому химику Я. Вант Гоффу. Экспериментальные данные Пфеффера оказались для Вант-Гоффа тем же, чем данные Тихо де Браге для Кеплера. Внимательно рассмотрев эти данные, Вант-Гофф увидел, что осмотическое давление в разных растворах получается одинаковым, если измерять концентрацию не в граммах на литр, а в молях, т. е. существенной является не масса, а число молекул растворенного вещества. Естественно считать, что молекулы растворенного вещества ведут себя как молекулы идеального газа. Таким образом, для выражения осмотического давления можно использовать уравнение Менделеева—Клапейрона
![]()
где m — масса растворенного вещества, i — масса моля, Л — газовая постоянная, Т — температура, V — объем. За теорию растворов Вант-Гофф через 15 лет получит Нобелевскую премию по химии. Вот какой важный вклад в науку внес Де Фриз, поговорив с Вант-Гоффом.
Теория Вант-Гоффа прекрасно выполнялась для растворов многих веществ, например для сахарозы, для водного раствора С02. Но для некоторых веществ осмотическое давление оказывалось больше расчетного, и не на какие-нибудь 10—15%, а вдвое! Погрешность составляла 100%. Вряд ли такую погрешность можно объяснить неточностью измерений.
Обдумывание возможных причин этого расхождения привело к важнейшему открытию. Единомышленник Вант-Гоффа шведский ученый С. Аррениус догадался, что если, например, для поваренной соли давление оказывается вдвое больше расчетного, то значит, в растворе вдвое больше частиц, чем молекул NaCl, т. е. молекула NaCl в воде распадается на две частицы: Na и Cl. Из сопоставления этого факта с другими Аррениуе пришел к идее электролитической диссоциации, к идее, что частицы, на которые распадаются многие вещества, и есть те самые ионы — носители электрических зарядов, с помощью которых еще Фарадей объяснял законы электролиза.
До Аррениуса считалось, что ионы возникают под действием электрического тока, но их участие в явлении осмоса показало, что это не так: ионы предсуществуют в растворе! Уже в самом растворе без действия тока имеются и движутся заряженные атомы и молекулы.
Так изучение осмоса привело к открытию двух главных «виновников» возникновения «животного электричества» — клеточных мембран и ионов; но об их роли пока еще никто не догадывался.
Горячо! Совсем горячо!
В 1887 г. вышли статьи Вант-Гоффа и Аррениуса в первом номере «Журнала физической химии», основанного В. Оствальдом. Все вокруг говорили о новом подходе к химии, о доказательстве существования ионов.
А в Берлине в этом году окончил университет 23-летний В. Нернст. Он станет знаменитым физиком и химиком, откроет третье начало термодинамики, получит Нобелевскую премию по химии и сменит М. Планка на посту директора Института физики в Берлине. А пока он скромный ассистент Оствальда, хотя и не новичок в науке. Работая как экспериментатор, он уже успел открыть новое явление: возникновение электрического поля в телах, в которых создан градиент температуры, если их поместить в магнитное поле. Теперь он занялся диссертацией и взялся за теоретическую работу о гальванических элементах. Конечно, до него такой теорией занимались великие умы: У. Томсон, Гиббс, Гельмгольц. Но ведь тогда не знали о ионах! Нернст решил рассмотреть связь э.д.с. гальванических элементов с диффузией ионов.
В 1889 г. его диссертация была готова и опубликована. В ней, в частности, была развита идея Вольта о том, что электрические явления могут возникать при соприкосновении двух разных жидкостей.
Что такое Нернстовский потенциал
Пусть в какой-то части сосуда находится электролит, а в другой — чистый растворитель. Как будет идти диффузия соли? Положительно и отрицательно заряженные ионы имеют разную массу, по-разному взаимодействуют с водой, следовательно, движутся в жидкости с разными скоростями. Пусть отрицательные ионы хлора движутся быстрее и при диффузии «убегают» вперед от катионов — возникает «расслаивание» отрицательных и положительных ионов. Это расслаивание вызывает появление в растворе электрического поля, которое будет тормозить убежавшие вперед анионы и подгонять отставшие катионы, выравнивая скорости их диффузии.
Естественно, Нернст в своей диссертации не ограничился таким качественным описанием, а как настоящий физик вывел формулу для диффузионного потенциала Уд, возникающего на границе контакта двух растворов с концентрациями электролита С1 и С2:
![]()
где и — скорость более быстрого иона, и — скорость более медленного иона, F — газовая постоянная, Р — число Фарадея, Т — температура.
Эта формула давала возможность подвергнуть прямой количественной проверке гипотезу Германа, согласно которой биопотенциалы возникают на границе двух участков протоплазмы — нормального и поврежденного, и через семь лет такую проверку предпримет замечательный русский ученый Василий Юрьевич Чаговец. Это будет первая работа по применению точных количественных методов к исследованию биопотенциалов.
Когда была опубликована диссертация Нернста, Чаговец еще учился в киевской гимназии. В 1892 г. он поступил в Петербургскую военно-медицинскую академию. Лабораторией физиологии там руководил И. Р. Тарханов — ученик Сеченова. Тарханов предложил студенту третьего курса Чаговцу заняться изучением электрических явлений в нерве лягушки, точно так же, как за 50 лет до того И. Мюллер предложил сходную тему студенту третьего курса Дюбуа-Реймону.
Чаговец, конечно, предпочитал гипотезу Германа фантастическим электромоторным молекулам Дюбуа. Но и гипотеза Германа была слишком абстрактной. Она говорила о каких-то реакциях в протоплазме, о каких-то молекулах, границах... И Чаговец развивает конкретный вариант теории повреждения, впервые объясняя возникновение биопотенциалов диффузией ионов.
Но какое же конкретное вещество создает потенциал повреждения?
Как видно из формулы, желательно, чтобы анионы и катионы вещества имели разную подвижность, а также, чтобы его концентрация в месте разреза сильно отличалась от концентрации в неповрежденном участке. Чаговец решил, что это вещество — угольная кислота. Он рассуждал так: разрез, как всякое раздражение, усиливает обмен веществ, а значит и выделение С02. На основании данных, полученных еще Л. Германом, Чаговец принял, что концентрация угольной кислоты в месте разреза увеличивается в 6,5 раз. Разность скоростей ионов Н+ и СОз_ давала для первого сомножителя формулы значение 0,8. Чаговец произвел расчет потенциала повреждения мышцы при комнатной температуре, получилось примерно 35 мВ. На самом деле потенциал повреждения мышцы составлял 50—60 мВ. Разница была не слишком велика, но все же вполне ощутима.
Работа Чаговца под названием «О применении теории диссоциации Аррениуса к электрическим явлениям на живых тканях» вышла в 1896 г., а на следующий год ее краткое изложение было опубликовано в «Журнале физической химии» Оствальда. В своей работе Чаговец пытался объяснить расхождение результатов расчета с экспериментом тем, что концентрация угольной кислоты при разрезе может быть выше, чем при утомлении мышцы. Но это предположение было опровергнуто прямыми экспериментами: например, нейтрализация угольной кислоты щелочью практически не меняет потенциала повреждения.
Чаговец остановил свое внимание на угольной кислоте не только потому, что знал о возникновении разности ее концентрации при работе мышцы, а еще и потому, что при ее диссоциации возникает ион водорода — самый быстрый из всех ионов. Скорости наиболее распространенных ионов примерно одинаковы, так что первый сомножитель в формуле для них вместо 0,8 будет 0,01—0,2, и в результате получатся значения разности потенциалов,, еще более далекие от реальных. Придумывать же какие-то фантастические ионы было совсем не в духе Чаговца. На первый взгляд, идея диффузии ионов как основы биопотенциалов зашла в тупик. Однако, хотя конкретный вариант теории биопотенциалов, предложенный Чаговцем, оказался ошибочным, основная идея Чаговца оказалась совершенно верной; Чаговец был очень близок к решению загадки.
Загадка решена
Еще в 1890 г. Вильгельм Оствальд, который продолжал заниматься полупроницаемыми искусственными пленками, предположил, что полупроницаемость может вызывать не только осмос, но и электрические явления. Осмос возникает тогда, когда пленка пропускает маленькие молекулы воды1 но не пропускает большие молекулы сахара. Но ведь и ионы могут иметь разные размеры! Тогда мембрана будет пропускать ионы только одного знака, например, положительные. Оствальд прямо показал, что на искусственных мембранах могут возникать заметно большие разности потенциалов, чем при свободной диффузии ионов на границе растворов с разной концентрацией. И не требуется быстрый ион водорода: годится любой ион, который умеет проходить через мембрану и имеет разные концентрации по разные ее стороны.
Действительно, если посмотреть на формулу и предположить, что мембрана для анионов непроницаема, т. е. V = 0, то можно видеть, что должны получаться большие значения для диффузионного потенциала:
![]()
Таким образом, Оствальд объединил формулу Нернста и знания о полупроницаемых мембранах. Он предположил, что свойствами такой мембраны объясняются потенциалы мышц и нервов и удивительное действие электрических органов рыб.
Эта идея Оствальда, как ни странно, прошла мимо биологов, хотя члены самой передовой тогда школы физиологов — школы Дюбуа-Реймона — были заняты спором как раз о той проблеме, путь к решению которой дал Оствальд. А может быть, именно спор помешал воспринять эту идею: Герман1 как автор гипотезы повреждения, не думал о мембране, а Бернштейн, возглавивший сторонников гипотезы предсуществования, не думал о ионах. К тому же и Герман,; и Бернштейн были уже не молоды: и тому и другому перевалило за 50,; возраст9 когда нелегко воспринимать новые идеи.
И все же для Бернштейна это оказалось возможным. Он оценил идею Оствальда, и, может быть, немалую роль в этом сыграло то, что в самом главном мембранная гипотеза была очень в духе школы Дюбуа, никакой таинственной «жизненной силы», никаких сложных неизвестных свойств клетки или протоплазмы! все, что она требует,— это мембраны и электролиты. Решающий шаг, который пришлось сделать Бернштейну, состоял в том, чтобы объяснить электрические свойства мышц и нервов не устройством этих органов в целом, а свойствами клеток, из которых состоят все ткани я органы. Наконец-то был прямо указан «виновник», создающий «животное электричество»,— клеточная мембрана, а «оружие» — перенос ионов. Таким образом, в гипотезе Бернштейна объединяются электрохимия и клеточная теория.
Ю. Бернштейн считается основателем, так называемой мембранной теории биопотенциалов. Его первая статья по мембранной теории вышла в 1902 г. Начинался новый век в электробиологии.
Мембранная теория
Давайте же посмотрим, сначала чисто качественно, как объясняется этой теорией возникновение биопотенциалов.
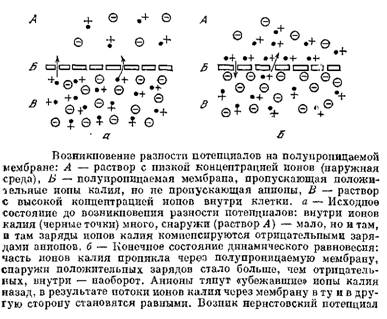
Что было важно для Бернштейна в строении органов и клеток? Мышца или нерв состоят из клеток, окруженных межклеточной жидкостью. Каждая клетка представляет собой мешочек или пузырек, покрытый оболочкой и содержащий жидкость другого состава.
Оболочка клетки — это и есть мембрана. Она отделяет клетки не друг от друга, а внутреннюю среду клетки от внешней межклеточной среды.
Пусть теперь внутри клетки имеется много свободных ионов какого-то элемента, например калия, а снаружи таких ионов нет или их гораздо меньше. Пусть клеточная мембрана пропускает только ионы К+ и не пропускает никаких других ионов. Тогда ионы К+ начнут выходить из клетки, где их много, наружу. Вместе с ними будет выноситься наружу их положительный заряд. Внутрь через мембрану будет проходить мало ионов, так как снаружи мало калия. В результате на клеточной мембране будет возникать разность потенциалов: снаружи клетки — «плюс», а внутри — «минус». Эта разность потенциалов будет тормозить движение новых положительных заряженных ионов калия наружу, и увеличивать поток этих ионов внутрь. Когда потоки ионов наружу и внутрь сравняются, установится динамическое равновесие и на мембране будет поддерживаться постоянная разность потенциалов. Это и есть потенциал покоя. Его величина описывается формулой Нернста.
Чтобы вывести эту формулу, воспользуемся тем, что согласно теории растворов Вант-Гоффа переход ионов из раствора с большей концентрацией С2 в раствор с меньшей концентрацией Сх представляет собой как бы расширение «ионного газа»: если в растворе с концентрацией С2 некоторое количество ионов занимает объем V2, то в растворе с концентрацией Сг они займут объем Vlt причем C1V1= СгУг. При этом меняется внутренняя анергия ионного газа, которая и переходит в потенциальную энергию зарядов, равную qE, где q — заряд, а Е — возникшая разность потенциалов.
Так как изменение внутренней энергии газа определяется только его начальным и конечным состояниями и не зависит от того, каким способом произошел переход из одного состояния в другое, попробуем найти изменение энергии в самом простом случае — когда при расширении газа совершается механическая работа.
Пусть у нас есть цилиндр с газом, закрытый поршнем, и пусть газ, расширяясь, движет поршень, совершая работу, равную произведению силы на путь: А = FI. Сила равна произведению давления газа на площадь поршня, т. е. А = PSI. Но произведение площади на перемещение есть изменение объема газа. Следовательно, изменение энергии газа при расширении определяется формулой AW=PаV.
Если бы давление Р при расширении, т. е. при изменении объема, не менялось, то работа была бы равна произведению Р, т. е. площади прямоугольника ABCD. Но из уравнения газового состояния RT = PV следует, что с увеличением объема давление падает по закону Р = ЯТ/У. Значит, при постоянной температуре Т работа расширения одного моля газа от Уг до У2 равна площади криволинейной трапеции под гиперболой и может быть вычислена по формуле
![]()
А так как концентрация газа обратно пропорциональна объему, то У2 = сУС2 и А = ИТ 1п.
Итак, если при расширении ионного газа его концентрация изменилась от Сг до С2, то внутренняя энергия этого газа меняется
Работа расширения газа при постоянном давлении Рх равна площади прямоугольника с вершинами I, II, III, IV, т. е А=Р1{У2—У1). Работа при переменном давлении Р = ВТ'IV равна площади криволинейной трапеции. т. е.
![]()
на величину А И7 = ЯТ на каждый моль. Но, с другой стороны, каждый моль одновалентного иона переносит заряд, равный — числу Фарадея. Отсюда и получаем, что ВТ 1п = ЕР и окончательно![]() . Таким образом, мы получили формулу Нернста, и вы теперь понимаете, откуда в этой формуле появилась газовая постоянная.
. Таким образом, мы получили формулу Нернста, и вы теперь понимаете, откуда в этой формуле появилась газовая постоянная.
Нам очень хотелось, чтобы, взглянув на формулу Нернста, вы не просто поняли ее смысл, но и увидели, какой труд за ней стоит. Это труд ботаников, измеривших осмотическое давление; физиков, выяснивших законы электричества, пополнивших науку понятиями заряда и разности потенциалов, открывших законы электролиза и газовые законы; химиков, создавших теорию растворов и электролитической диссоциации; математиков, труды которых позволили Ньютону и Лейбницу создать дифференциальное и интегральное исчисление. О некоторых из этих работ мы вам кратко рассказали, а о многих не можем рассказать, так как они лежат далеко в стороне от нашей основной темы. Но на этом примере нам хотелось показать, как в одной формуле собрались воедино труды и идеи тысяч ученых разных времен и стран.
Снова о Бернштейне
Но не следует думать, что достаточно было только высказать общую идею, чтобы все сразу с ней согласились; и самому Бернштейну, а затем и его последователям потребовались годы и десятилетия напряженного труда, споров, сомнений и разочарований, пока, наконец, удалось доказать свою правоту. Сама же по себе гипотеза мембранного потенциала выглядела в то время нисколько не лучше, чем гипотеза повреждения или даже гипотеза электромоторных молекул. Поэтому первую статью с изложением своих взглядов Бернштейн выпустил в свет только тогда, когда он сумел получить экспериментальные данные, которые могли послужить аргументом, хотя и косвенным, в их пользу.
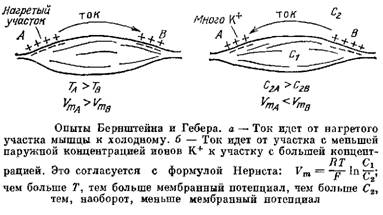
Найти один из таких аргументов Бернштейну помогла все та же формула Нернста. В эту формулу кроме неизвестных концентраций неизвестно какого иона входит еще Т — температура, а уж ее-то можно было не только измерить, но и менять по своему усмотрению.
Проведя серию опытов на мышце лягушки, Бернштейн показал, что если нагревают продольную неповрежденную поверхность мышцы, то в определенном диапазоне регистрируемый потенциал повреждения, действительно, прямо пропорционален Т, как и следует из формулы Нернста. Нагревание самого разреза, напротив, не влияло на значение потенциала. Это и был аргумент против теории повреждения.
Другая серия опытов была еще более эффектной. Бернштейн показал, что если нагреть один конец целой неповрежденной мышцы, то от нагретого к холодному месту поверхности мышцы потечет ток, Этот результат также прямо следует из теории: в более теплом месте поверхности возникает больший положительный потенциал, чем в холодном.
Как мы уже упоминали, в 1902 г, вышла первая статья Бернштейна по мембранной теории. Этот год и считается годом ее рождения.
Очень слабым местом мембранной гипотезы было полное отсутствие данных о том, какой именно ион вызывает потенциал. Но вот в 1905 г. в Берлине молодой сотрудник Нернста Гебер обнаруживает, что все соли, содержащие калий, оказывают сходное действие на мышцу: участок мышцы, на который действует раствор такой соли, приобретает отрицательный потенциал по отношению к другим участкам мышцы.
Бернштейн сразу оценивает значение работы Гебера — ведь мембранная теория объясняет эти результаты очень просто: стоит только предположить, что К+ и является тем ионом, который создает потенциал. Все соли, содержащие калий, диссоциируя в растворе, увеличивают наружную концентрацию ионов калия, при этом отношение концентраций СуСг падает, и область, на которую действуют солью, приобретает меньший, чем другие участки, потенциал.
Факты сами по себе мало о чем говорят. Герман, почти за 40 лет до Берпштейна, наблюдая влияние температуры на мышцу, видел, что при нагревании участка, удаленного от разреза, потенциал растет. Эти факты не имели тогда истолкования и поэтому были полузабыты. Влияние солей калия на потенциал, оказывается, тоже было описано за 10 лет до Гебера в книге Бидермана, посвященной электробиологии, и на это тоже не обратили внимания. Только теория придает экспериментальным фактам смысл, позволяет отделить существенные от второстепенных, освещает их значение внутренним светом,
В 1912 г. вышла в свет большая книга Бернштейна «Электробиология». В этой книге дано объяснение не только потенциала покоя, но и многих других явлений, о чем мы расскажем дальше. Но очень важно, что Бернштейн предсказывает гораздо более общее значение мембранной теории, чем просто объяснение работы мышцы и нерва. Ведь по мембранной теории для возникновения ПП никаких специальных свойств от клетки не требовалось, и вполне естественно было предположить, что ПП присущ не только этим тканям, что все клетки могут вырабатывать электричество. Бернштейн объясняет работу электрического органа рыб, работу желез, движения насекомоядных растений и даже пытается объяснить движение внутриклеточных частиц — хромосом — при делении клеток клеточными потенциалами.
Эта книга заставляет вспомнить о другой книге, книге В.Ю. Чаговца, вышедшей в 1903 г. Невольно возникает впечатление, что Вернштейн идет прямо по стопам Чаговца, у которого в книге мы видим те же главы: о работе желез и кожных потенциалах, о работе электрических органов, о потенциалах в растительных клетках. Пришло время обобщений. Накопленный за столетие материал оказалось возможным увидеть с единой точки зрения.
Однако и после выхода в свет книги Бернштейна мембранная теория не вызвала особого восторга. И это закономерно, так как несмотря на достаточно глубокую разработку теории, проведенную Бернштейном, все экспериментальные ее подтверждения были косвенными: они доказывали следствия теории, а как хорошо понимают математики, если из А следует Б, то это не означает, что из Б следует А. Поэтому все основные утверждения мембранной теории были по существу гипотезами, и для доказательства ее истинности не было другого пути, как доказать правильность гипотез, положенных в ее основу, а именно доказать, что:
а) клетки имеют мембрану, проницаемую для какого-то иона,
б) концентрация этого иона внутри клетки сильно отличается от его концентрации в наружной среде;
в) потенциал возникает на мембране только за счет ее проницаемости для этого иона и потому равен нернстовскому потенциалу.
Пока данных, прямо подтверждающих истинность этих фундаментальных положений мембранной теории, не было, следствие из нее могло получить и получало в продолжавшемся споре другое толкование. Так, влияние температуры могло объясняться ускорением химических реакций пусть не в месте повреждения, а на поверхности мышцы; влияние калия на величину ПП также могло быть истолковано как его химическое взаимодействие с веществом клетки и т. п.
Наибольшим нападкам подвергалась основа основ теории — само существование мембраны, которую никак не удавалось увидеть в самый сильный микроскоп. Возражения относились не только к качественной картине явлений: были созданы теории, которые объясняли ПП свойствами протоплазмы и предсказывали ту же линейную зависимость ПП от температуры.
Таким образом, установить, какая теория верна, с помощью косвенных экспериментов оказалось невозможным. А прямые экспериментальные доказательства мембранной теории были получены очень нескоро: они требовали исследований на клеточном уровне, а в то время клетка как таковая еще не стала объектом электрофизиологического эксперимента.
Доказательства мембранной теории. Что снаружи? Что внутри?
Из трех основных «действующих лиц» мембранной теории — мембрана, наружная среда, внутренняя среда — довольно хорошо была исследована лишь наружная среда, и не только потому, что она была наиболее доступной. Химическим составом среды, окружающей клетки организма, биологам уже давно пришлось заниматься вне всякой связи с мембранной теорией. При любых экспериментах на изолированных органах их нужно держать в специально подобранном растворе. Например,, сердце лягушки нельзя оставлять просто на воздухе — оно высохнет и перестанет работать, но нельзя поместить его и в воду — под действием осмоса клетки органа погибнут.
Естественно, подбор таких растворов был связан с исследованием состава крови, гемолимфы и т. д., поэтому этот состав и был довольно хорошо известен.
Поэтому важный для подтверждения мембранной теории солевой — а значит и ионный — состав таких внеклеточных жидкостей был хорошо известен. Оказалось, что «главный поставщик» ионов всех таких растворов — обыкновенная поваренная соль. Предполагают, что в этом отражается происхождение животных, главные ионы морской воды — тоже натрий и хлор.
Особенно интересно и очень существенно с точки зрения мембранной теории то, что хотя абсолютная концентрация солей в тканевой жидкости разных организмов в зависимости от условий их обитания сильно варьирует, но соотношение концентраций ионов натрия и калия в среде, окружающей клетки организма, примерно одинаково для всех животных — от медузы до человека. Концентрация калия примерно в 50 раз меньше, чем натрия. Таким образом, у всех животных эта среда по существу представляет собой в большей или в меньшей мере разбавленную морскую воду.
Гораздо труднее было определить ионный состав внутриклеточного содержимого. Ученые пытались использовать для определения этого состава самые разные методы, о которых мы не будем вам подробно рассказывать, по все данные, полученные этими методами, были лишь приблизительными: клетки слишком малы, а между ними всегда имеется межклеточное вещество и жидкость. Кроме того, сторонникам мембранной теории надо было не просто показать наличие калия внутри клеток, а наличие именно свободных, несвязанных ионов калия. Эта проблема была, как уже не раз случалось, решена с помощью двух подходов: и разработкой новых методов измерения, и подбором подходящего объекта.
В 1936 г. английский специалист по головоногим моллюскам Дж. Юнг обнаружил у кальмаров нервное волокно, диаметр которого доходил до миллиметра, т. е. по клеточным масштабам гигантское, хотя сам моллюск вовсе не был гигантским. Такое волокно, выделенное из организма и помещенное в морскую воду, не погибало. Наконец-то появилась живая клетка, в которую можно было проникнуть, с которой можно было работать.
В 1939 г. английские ученые А. Ходжкин и его ученик А. Хаксли впервые измерили разность потенциалов на мембране животной клетки. Удалось также доказать, что внутри этого волокна действительно много ионов калия, что эти ионы представляют собой «ионный газ», т. е. могут участвовать в создании мембранного потенциала. При этом расчетное значение ПП неплохо совпадало с непосредственно измеренным.
Распространить эти данные с уникальной клетки — гигантского аксона — на обычные клетки стало возможным, когда в 1946 г. американские ученые Джерард и Линг разработали новую методику — методику микроэлектродов. Микроэлектрод — это вытянутая иэ нагретой стеклянной трубочки тоненькая пипетка с диаметром кончика менее 1 мкм, заполненная раствором электролита.
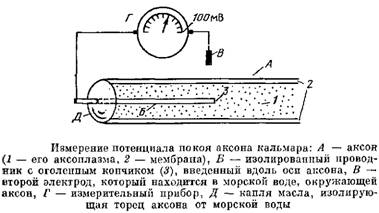
Стекло играет роль изолятора, а электролит — проводника. Такой электрод можно ввести в любую клетку, практически ее не повреждая.
Новая техника эксперимента быстро получила широкое распространение в самых разных исследованиях и за несколько лет буквально завоевала мир. Сам Джерард в США зарегистрировал ПП мышцы лягушки, сокращение которой наблюдал Гальвани и токи повреждения, в которой изучал Дюбуа-Реймон. Дж. Экклс в Новой Зеландии зарегистрировал ПП клетки мозга, Б. Катц в Англии начал изучать с помощью новых электродов действие нерва на мышцу; в Швейцарии С. Вайдман первым сумел ввести хрупкий микроэлектрод в сокращающееся сердце. В нашей стране пионером микроэлектродных исследований стал сотрудник Киевского государственного университета Платон Григорьевич Костюк, который начал применять эту технику в середине 50-х годов.
Вскоре были получены достаточно полные данные о величине ПП для разных клеток, ПП был обнаружен не только у мышечных и нервных клеток, но и у эритроцитов, клеток кожи, печени и др. Если считать, что причиной ПП служит разность концентраций ионов калия во внутренней и наружной средах клеток, разделенных мембраной, а все клетки в этом отношении устроены в общем сходно, то наличие ПП совершенно закономерно, хотя и не ясно, зачем понадобился ПП, например, клеткам слюнной железы или печени.
О пользе бракованных микроэлектродов
Теперь, когда достаточно точное измерение ПП в отдельной клетке не представляло проблемы для проверки мембранной теории по формуле Нернста, оставалось сделать последний шаг — научиться так же точно определять ионный состав в клетке. И замечательно, что развитие микроэлектродной техники помогло решить и эту задачу.
Дело в том, что изготовление микроэлектродов — это целое искусство: и стекло подобрать, чтобы хорошо тянулось, и режим нагрева и скорость вытягивания, чтобы копчик не обламывался и внутренний канал не закупоривал, и т. д. и т. п. И вдруг оказалось, что даже если в опыте использовался плохой, «бракованный» электрод, у которого на кончике вообще не было отверстия, потенциал все равно регистрировался, как будто стекло было не изолятором, а проводником. Когда стали в этом разбираться, оказалось, что, действительно, тоненькие стенки микроэлектродов из некоторых сортов стекла представляют собой — что бы вы думали? — полупроницаемую мембрану, т. е. избирательно пропускают ионы какого-либо сорта.
Если такой электрод, заполненный раствором с известной концентрацией изучаемого иона, опустить в раствор, содержащий тот же ион, то, зная значение возникшего нернстовского потенциала, можно использовать формулу Нернста для решения обратной задачи — определить неизвестную концентрацию иона в исследуемом растворе.
Использование таких электродов в сочетании с методом меченых атомов и другими методами позволило определить ионный состав внутриклеточного содержимого. Оказалось, что, как и предсказывала мембранная теория, внутри клеток концентрация ионов калия в 30—40 раз выше, чем в наружной среде. Оказалось также, что соотношение калия и натрия внутри клеток животных совершенно иное, чем вне их: в то время как вне клетки много натрия, в клетках, наоборот, калия почти в 10 раз больше, чем натрия.
Высокое содержание ионов калия было обнаружено не только в нервных, но и в разных других клетках организма. Таким образом, внутреннее содержимое всех клеток животных оказалось непохожим на морскую воду. В ходе эволюции эти клетки создали свою особую внутреннюю среду.
Клеточная мембрана
Итак, вопросы об ионном составе внутренней и внешней среды, а также о величине ПП были решены; это косвенно доказывало и существование мембраны.
Увидеть мембрану удалось лишь в середине 50-х годов нашего века. Это было сделано с помощью электронного микроскопа, так как толщина мембраны составляет всего 7 —15 нм и в световой микроскоп ее увидеть нельзя. Однако еще до того, как ее непосредственно увидели, сомнения в ее существовании практически исчезли; многочисленные исследования, опирающиеся на факты из разных областей, свидетельствовали, что мембрана существует; более того, к этому времени было известно достаточно много об ее устройстве и свойствах: толщине, электрическом сопротивлении и т. д.
Обычно, говоря о научных предсказанниях, рассказывают об открытии «на кончике пера» планеты Нептун, или о предсказании существования атомов и молекул, или о предсказании Менделеевым химических элементов, которые заполнили оставшиеся пустыми соответствующие клетки его таблицы. Но и в биологии можно найти немало примеров таких научных предсказаний. Например, Гарвей, создавая теорию кровообращения, предсказал существование капилляров, но увидеть их смог только М. Мальпиги после изобретения микроскопа. Точно так же клеточная мембрана была предсказана задолго до появления электронного микроскопа.
Очень много сведений о свойствах мембраны дало изучение проникновения разных веществ в клетку. Это особый, весьма увлекательный и весьма запутанный рассказ, который мы не можем тут привести. Но общий вывод из него весьма поучителен. Дело в том, что, как сейчас выяснено, разные вещества попадают в клетку разными способами: одни, растворяясь в жирах мембраны, проникают в клетку прямо через них, другие вещества, которые не могут проходить через жиры, проникают через особые «поры», образованные мембранными белками, третьи — совсем иначе, например, «заглатываясь» клеткой, в которой образуется отшнуровывающийся и уходящий внутрь мембранный пузырек; и это еще не все способы. Между тем ученые стараются объяснить некоторое явление с единой точки зрения. Для науки идеалом является, например, теория Максвелла, которая позволила связать воедино электрические, магнитные и оптические явления, описав их основные свойства несколькими уравнениями. Такую же единую теорию искали и ученые, изучавшие клеточную проницаемость. Однако, как мы теперь понимаем, в случае клеточной проницаемости такой единой теории просто не существовало. При наличии многих принципиально различных способов проникновения веществ в клетку для каждой теории, претендующей на полное объяснение фактов с единой точки зрения, находился опровергающий ее эксперимент. Мы все сталкивались с аналогичной ситуацией; вспомните, как Вольта пытался объяснить с единой точки зрения и контактную разность потенциалов, и работу химических элементов. Так, естественное стремление ученого к созданию единой теории иногда играет роль тормоза в развитии науки. Но вернемся к мембране.
Поверхность всякого организма играет особую роль, так как это — место соприкосновения организма и среды. Именно на поверхности находится защитный слой; на поверхности находятся органы чувств; через поверхность организм получает воду, пищу и кислород. Все эти соображения применимы и к отдельной клетке. Через поверхностную мембрану клетка получает разные вещества, через нее же выводятся наружу продукты обмена. В мембрану встроены особые белки-рецепторы — «органы чувств» клетки. Но мембрана покрывает не только поверхность клетки, внутри клетки тоже имеется множество мембран, которые делят клетку на отсеки, окружают ее органеллы. Особой двухслойной ядерной мембраной окружено ядро клетки, свою мембрану имеют «энергетические станции клетки» — митохондрии. В пузырьках из мембран — лизосомах — хранятся некоторые клеточные ферменты и т. д.
Про мембраны теперь известно так много, что возникла особая наука — мембранология; в нашей стране выходит специальный журнал «Биологические мембраны», а за рубежом журнал «Биология мембран». Для подробного рассказа о мембранах нужно было бы написать отдельную книгу. Такие книги уже написаны, поэтому мы расскажем здесь о мембранах лишь то, что будет нам совершенно необходимо для дальнейшего.
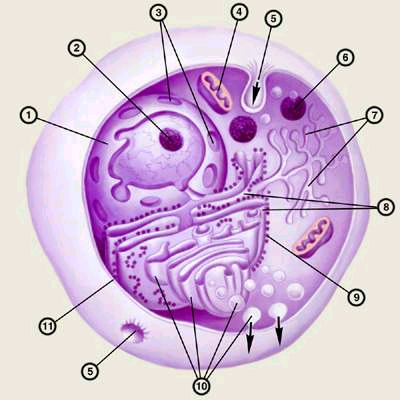
Клеточная мембрана представляет собой жидкую пленку, образованную жироподобными веществами — липидами. Она состоит из двух слоев липидных молекул, в которые встроены молекулы белка.
Нас, прежде всего, будут интересовать, конечно, электрические характеристики мембраны, которые были определены с помощью самых разных методов. Эта работа была начата еще в 1910 г. в лаборатории Нернста и в ней принимал участие тот самый Гебер, который открыл влияние солей калия на потенциал мышц. Измерения проводились на суспензии клеток. Основной метод измерения состоял в том, что через суспензию пропускали ток разной частоты и определяли ее удельное сопротивление. Была развита теория, которая позволяла отдельно определить сопротивления мембраны клетки и ее протоплазмы, так как их отношение зависело от частоты пропускаемого тока.
Развивая это направление, Г. Фрикке в 1925 г. показал, что мембрана ведет себя в опытах как параллельно соединенные сопротивление и емкость, т. е., как говорят, выяснил эквивалентную схему мембраны. Схема эта первоначально была установлена Для мембраны эритроцитов; Фрикке использовал при измерениях частоты до 4,5 МГц, так что успеху его работы способствовало развитие физических приборов — появление генераторов высокой частоты. В дальнейшем на самых разных клетках удалось измерить сопротивление мембраны, ее емкость, а также удельное сопротивление протоплазмы. Г. Фрикке и Г. Кертис работали с эритроцитами, лейкоцитами, одноклеточной водорослью — хлореллой, дрожжами; К. Кол работал с яйцеклетками иглокожих, мышечными и нервными клетками.
Оказалось, что удельная емкость мембраны у всех изученных клеток составляет примерно 1 мкФ/см2. Не слишком сильно отличается у разных клеток и удельное сопротивление протоплазмы, оно обычно близко к 100 Ом-см, т. е. примерно такое же, как у морской воды. Напротив, удельное сопротивление мембраны у разных клеток оказалось весьма разным! Так, например, у яйцеклеток морского ежа удельное сопротивление мембраны составляет всего 100 Ом-см2, а у водоросли нигеллы — 100 000 Ом-см2 *).
Почему же у разных клеток мембрана имеет одинаковую удельную емкость и столь разное удельное сопротивление? Дело в том, что емкость мембраны определяется ее липидным слоем, который можно рассматривать как диэлектрик, находящийся между двумя пластинами конденсатора.
А вот сопротивление мембраны определяется не липидами, а теми молекулами белка, которые встроены в жидкий липидный слой и плавают в нем, как айсберги, «прошивая» мембрану насквозь. Эти молекулы в разных клетках отличаются и по числу, и по свойствам — отсюда и разное удельное сопротивление. Если убрать из липидного бислоя белок, то удельное сопротивление такой «чистой» мембраны оказывается весьма высоким — порядка 108 Ом-см2.
С помощью изотопных и других методов было показано, что клеточная мембрана действительно является полупроницаемой: она хорошо пропускает положительные ионы калия, но не пропускает клеточные анионы. Но вернемся к мембранной теории.
Опыты на «голой» мембране — торжество мембранной теории
В 1961 г. А. Ходжкин и его сотрудники Бекер и Шоу поставили очень красивый опыт. Еще когда Юнг открыл гигантский аксон у кальмара, он заметил, что при надавливании на такой перерезанный аксон его содержимое выдавливается из него, как паста из тюбика, и остается пустая «шкурка». Вот с этой оболочкой, т. е. мембраной аксона, и начали работать Ходжкин и его сотрудники. Они брали аксон, клали его на резиновую подложку, чтобы не повредить, и с помощью резинового ролика выдавливали из него протоплазму. Получалась пустая трубка из одной мембраны. Эту трубку через пипетку, вставленную в торец, можно было заполнять разными растворами. Таким образом, появилась возможность работать на мембране нервного волокна, меняя произвольно концентрацию веществ как снаружи, так и внутри аксона.
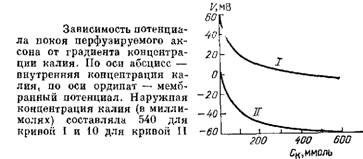
Оказалось, что при заполнении аксона растворами, содержащими такую же концентрацию К+, как и протоплазма, в аксоне возникал нормальный потенциал покоя порядка 50—60 мВ. При этом выяснилось, что природа отрицательных ионов, которые вводились в волокно вместе с калием, не слишком существенна. Если концентрация калия внутри волокна и в наружной среде была одинаковой, то потенциал покоя в полном соответствии с формулой Нернста не возникал. Если же внутрь волокна накачивали морскую воду, а в наружную среду — раствор с высокой концентрацией калия, то знак потенциала покоя менялся: внутри волокна возникал знак «плюс», а снаружи — «минус»; при этом величина ПП соответствовала формуле Нернста. Таким образом, введение внутрь волокна солевых растворов разного состава подтвердило результаты, полученные ранее при изменении наружной концентрации ионов калия.
Эти опыты показали, во-первых, решающую роль мембраны в возникновении ПП — ведь протоплазма со всеми органеллами и белковыми молекулами попросту отсутствовала, а во-вторых, — решающую роль ионов калия в этом процессе. Мембрана нервного волокна работала так, как это предсказывалось мембранной теорией Бернштейна.
Мембранная теория требует уточнений
Часто, когда теория достигает зрелости, получает признание, живет, начинает расти точность измерений и обнаруживаются отступления от теории, требующие объяснений. Законы Кеплера утверждали, что планеты движутся по эллипсам, но при более точных наблюдениях обнаружилось, что орбиты не вполне эллипсы, что существуют возмущения, которые были объяснены как результат действия на планету силы притяжения других планет. Именно учет возмущений позволил открыть планету Нептун.
То же самое произошло и с мембранной теорией. Когда методы измерения ПП и ионных концентраций достигли достаточно высокой точности, обнаружилось, что на самом деле измеряемый ПП всегда ниже, чем вычисленный теоретически по формуле Нернста. Ясно, что существовал какой-то «возмущающий» фактор, не учтенный теорией Бернштейна.
Найти причину расхождения эксперимента и теории помог опять-таки кальмар. Работая с гигантским аксоном, А. Ходжкин и Б. Катц установили, что ПП, измеряемый в опыте, оказывается ближе всего к теоретическому в одном особом случае: если из наружной среды убирается натрий.
Но причем здесь натрий? Если, как считал Берн-штейн, мембрана проницаема только для калия, то натрий вообще не должен влиять на ПП! А что если допущение Бернштейна ошибочно? Что будет, если, посягнув на «основу основ» мембранной теории, предположить, что натрий тоже проходит через мембрану? Легко понять, что тогда ионы натрия, которых снаружи клетки гораздо больше, чем внутри, будут идти по градиенту концентрации внутрь, создавая свой нернстовский потенциал, противоположный по знаку тому, который создается калием, и значит, будут снижать ПП. Это равносильно тому, что к мембране подключен не один, а два соединенных навстречу друг другу источника э.д.с. Э.д.с. этих источников определяются концентрационными градиентами калия и натрия. Ходжкин и Катц проверяют свое предположение экспериментально, меняя концентрацию натрия во внешней среде и сравнивая значения ПП с теоретическими. Оказалось, что для совпадения экспериментальных данных с расчетными приходится принять, что сопротивление, через которое идут ионы натрия в 25 раз больше, чем для калия. Это означает, что проницаемость мембраны для ионов натрия, хотя и в 25 раз меньше, чем для калия, но все же не равна нулю.
Вскоре предположение, что ионы натрия могут проходить через мембрану гигантского аксона, было прямо доказано в опытах с радиоактивным изотопом натрия. В ходе этих работ ученые открыли особое вещество, тетродотоксин, блокирующее движение натрия через мембрану. Теперь удалось показать, что если добавить в морскую воду, окружающую гигантский аксон, тетродотоксин и натрий перестает идти через мембрану, то ГШ начинает нарастать и достигает значения, в точности равного тому, которое предсказывается формулой Нернста. Тем самым роль натрия была полностью доказана.
На первый взгляд кажется, что уточнения, внесенные Ходжкином и Катцем в мембранную теорию Бернштейна, несущественны. Всего-то и оказалось, что мембрана пропускает, кроме калия, немного натрия: калий создает потенциал одного знака, натрий — другого, в результате получается ПП чуть ниже, чем только от калия.
Однако на самом деле ситуация меняется в корне.
На мембране устанавливается некоторый промежуточный потенциал, который, естественно, не является равновесным ни для одного из этих ионов. Поэтому и тот и другой ион будут двигаться по градиенту концентрации: ионы натрия — внутрь, а ионы калия — наружу, пока не уравняются концентрации, по крайней мере, одного из них.
Похожие работы
... ). Но некоторые однофункциональные белки могут иметь сходное строение у далеких групп организмов (к примеру, инсулин собаки и человека). В процессе жизнедеятельности белковые молекулы постепенно разрушаются, теряют свою структуру — денатурируют. Их активность падает, и клетки заменяют их новыми. В организмах постоянно происходит синтез необходимых белков. иосинтез белковых молекул — сложный ...
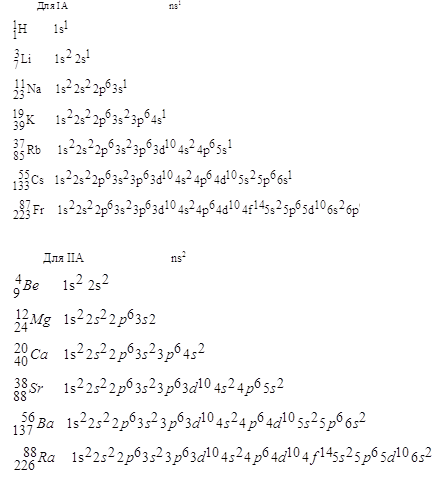
... в виде соединений. По содержанию в организме человека натрий (0,08%) и калий (0,23%) относятся к макроэлементам, литий, рубидий и цезий – к микроэлементам. Натрий и калий относятся к жизненно необходимым элементам, постоянно содержатся в организме, участвуют в обмене веществ. Натрий Содержание натрия в организме человека массой 70 кг – около 60 г: 44% - во внеклеточной жидкости, 9% - во ...
... составляют тела клеток, дендриты и немиелинизированные аксоны, организованные в комплексы, которые включают бесчисленное множество синапсов и служат центрами обработки информации, обеспечивая многие функции нервной системы. Белое вещество состоит из миелинизированных и немиелинизированных аксонов, выполняющих роль проводников, передающих импульсы из одного центра в другой. В состав серого и белого ...

... в репликации и распределении наследственной информации между дочерними клетками, а следовательно, и в регуляции клеточного деления и процессов развития организма. Обмен веществ и превращение энергии в клетке. Все живые организмы на Земле представляют собой открытые системы, способные активно организовывать поступление энергии и вещества извне. Энергия необходима для ...
0 комментариев