Корково-подкорковые отношения в процессах высшей нервной деятельности
Современные исследования подтверждают высказывание И.П. Павлова, что условно-рефлекторная деятельность осуществляется совместной работой коры и подкорковых структур. Из рассмотрения эволюции мозга как органа высшей нервной деятельности следует, что способность к образованию временных связей, обеспечивающих приспособительное поведение, проявляли структуры промежуточного мозга у рыб и стриарных (полосатых) тел у птиц, представляющие собой филогенетически наиболее молодые его отделы. Когда же у млекопитающих над этими отделами мозга возникла филогенетически самая молодая новая кора, осуществляющая наиболее тонкий анализ сигналов, то к ней перешла ведущая роль в образовании временных связей, организующих приспособительное поведение.
Структуры мозга, оказавшиеся подкорковыми, сохраняют в какой-то мере свои способности к замыканию временных связей, которые обеспечивают приспособительное поведение, свойственное тому уровню эволюции, когда эти структуры были ведущими.
Взаимодействие коры и подкорковых отделов мозга осуществляется и путем тонических влияний, регулирующих функциональное состояние нервных центров. Нейрофизиологические исследования механизмов подкорковых влияний на кору показали, что ретикулярная формация среднего мозга оказывает на нее восходящее активирующее действие. Получая коллатерали от всех афферентных путей, ретикулярная формация участвует во всех реакциях поведения, обусловливая деятельное состояние коры. Однако ее активирующее влияние при условном рефлексе организуется сигналами из проекционных зон коры. Раздражение ретикулярной формации вызывает изменение электроэнцефалограммы в виде ее десинхронизации, характерной для состояния активного бодрствования.
Иное действие на функциональное состояние коры оказывают неспецифические ядра таламуса. Их низкочастотное раздражение приводит к развитию в коре процессов торможения, которое может привести к засыпанию животного, и пр. Раздражение этих ядер вызывает в электроэнцефалограмме появление своеобразных волн - «веретен», которые переходят в медленные дельта-волны, характерные для сна. Ритм веретен может определяться тормозными постсинаптическими потенциалами (ТПСП) в нейронах гипоталамуса. Наряду с регуляторным влиянием неспецифических подкорковых структур на кору наблюдается и обратный процесс. Такие двусторонние корково-подкорковые взаимные влияния являются обязательными в осуществлении механизмов формирования временных связей.
Некоторые исследователи рассматривают факты об участии подкорковых структур в процессах высшей нервной деятельности как основание для того, чтобы считать их местом замыкания временных связей. Так возникло представление о «центрэнцефалической системе» как ведущей в поведении человека (У. Пенфилд, Г. Джаспер, 1958).
Как показали многочисленные исследования, участие подкорковых структур в процессах высшей нервной деятельности не ограничивается регуляторной ролью ретикулярной формации среднего мозга и лимбических структур. Ведь уже на подкорковом уровне происходят анализ и синтез действующих раздражителей и оценка их биологического значения, в значительной мере определяющая характер образующихся связей с сигналом. Использование показателей формирования наиболее коротких путей, по которым сигнал достигает разных подкорковых структур мозга, выявило наиболее выраженное участие в процессах обучения задних отделов таламуса и гиппокампа. Роль гиппокампа в явлениях памяти подтверждается многими фактами. Наконец, нет оснований предполагать, что способность к примитивной замыкательной деятельности структур мозга, которая приобреталась в эволюции, когда они были ведущими, теперь у них полностью исчезла, когда эта функция перешла к новой коре.
Таким образом, корково-подкорковые отношения определяются регулированием функционального состояния коры активирующей системой - ретикулярной формацией среднего мозга и тормозной системой неспецифических ядер таламуса, а также возможным участием в образовании примитивных временных связей на нижнем уровне сложных иерархических механизмов высшей нервной деятельности.
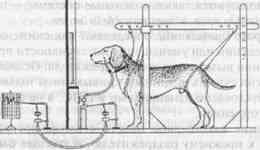
Межполушарные отношения. Каким образом полушария головного мозга, являющиеся парным органом, участвуют в процессах образования условных связей? Ответ на этот вопрос был получен в опытах на животных, которым была сделана операция «расщепления» мозга путем перерезки мозолистого тела и передней комиссуры, а также продольного разделения перекреста зрительных нервов. Если у оперированной таким образом обезьяны выработать условный рефлекс на световой стимул, подаваемый на один глаз, а затем подать его на другой глаз, то никакой реакции не последует. «Обучение» одного полушария оставляло другое «необученным». Однако при сохранении мозолистого тела и другое полушарие оказывается «обученным». Мозолистое тело осуществляет межполушарный перенос навыка.
Предположения о месте замыкания временных связей в больших полушариях головного мозга. Изучение двигательных условных рефлексов показало сложную структуру образующихся при этом временных связей. Каждое совершающееся по сигналу движение само становится сигнальным для складывающихся при этом двигательных координации. Образуются две системы временных связей: на сигнал и на движение.
Исходя из того, что условные рефлексы сохраняются при хирургическом разделении сенсорных и моторных корковых областей и даже после множественных разрезов коры, а также учитывая, что кора обильно снабжена как входящими, так и выходящими путями, было высказано предположение, что замыкание временных связей может происходить в каждом ее микроучастке между его афферентными и эфферентными элементами, которые активируют центры соответствующих безусловных рефлексов, служащих подкреплением. Такое предположение соответствует представлению о возникновении временной связи в пределах анализатора условного раздражителя, мнению о возможности «местных» условных рефлексов, замыкающихся внутри проекционных зон, и заключению, что в замыкании временной связи ключевую роль играет всегда афферентное звено.
Нейронная структура временной связи в коре больших полушарий. Современные сведения о микроскопическом строении коры больших полушарий в сочетании с результатами электрофизиологических исследований позволяют с известной степенью вероятности судить о возможном участии тех или иных корковых нейронов в образовании временных связей.
Высокоразвитая кора мозга млекопитающих, как известно, делится на шесть слоев разного клеточного состава. Приходящие сюда нервные волокна заканчиваются большей частью у клеток двух видов. Один из них - это вставочные нейроны, расположенные во II, III и отчасти IV слоях. Их аксоны идут в V и VI слои к крупным пирамидным клеткам ассоциативного и центробежного типа. Это наиболее короткие пути, которые, возможно, и представляют собой врожденные связи корковых рефлексов.
Другой вид клеток, с которыми приходящие волокна образуют наибольшее число контактов, составляют кустовидно ветвящиеся округлые и угловатые короткоотростчатые клетки, часто имеющие звездчатую форму. Они располагаются главным образом в IV слое. Их количество увеличивается с развитием мозга млекопитающих. Звездчатые клетки являются основными воспринимающими корковыми клетками анализаторов и увеличение их численности в эволюции представляет собой морфологическую основу достижения высокой тонкости и точности отражения окружающего мира.
Система вставочных и звездчатых нейронов может вступать в бесчисленное множество контактов с ассоциативными и проекционными крупными нейронами пирамидной формы, расположенными в V и VI слоях. Ассоциативные нейроны своими аксонами, проходящими через белое вещество, соединяют между собой разные корковые поля, а проекционные нейроны дают начало путям, соединяющим кору с низшими отделами мозга.
Согласно такому представлению о функциях корковых клеток замыкание временной связи осуществляется в системе звездчатых, вставочных и ассоциативных нейронов. Возможны два типа замыкания. Один из них - это замыкание с участием преимущественно зрительных, слуховых и других экстероцептивных раздражителей, которое происходит с помощью всех указанных нейронов, начиная со звездчатых. Другой - замыкание с участием преимущественно проприо- и интероцептивных раздражителей, происходящее прямо через вставочные нейроны верхних слоев без участия звездчатых нейронов. Предполагают, что именно поэтому некоторые условные рефлексы на проприо- и особенно интероцептивные раздражители, осуществляясь по коротким путям, не вовлекают в деятельность механизмы высших функций мозга человека и, не получая четкого «зеркального» отображения в сознании, проходят помимо него.
В качестве основы для формирования временных связей, как переходную форму от суммационного к настоящему условному рефлексу рассматривают доминанту. В доминантном очаге возникают сложные взаимоотношения между нервными и глиальными клетками. Формирование доминанты имеет динамичный характер. Она проходит ряд стадий среди которых стадия специализации имеет особое значение для организации целенаправленного поведения.
Деятельность неспецифических структур мозга, в частности восходящей активирующей системы ретикулярной формации среднего мозга, как показали исследования, действительно играет важную роль в образовании временных связей, обеспечивая высокую степень работоспособности высших отделов мозга. Замыкание временных связей является многоуровневым процессом, в который вовлекаются как кора, так и подкорковые образования, причем большую роль в этих процессах играет гиппокамп.
Широкое участие различных структур мозга в образовании временных связей проявляется и в синхронизации ритмов электрической активности корковых областей и многих подкорковых структур. Однако функция большинства из них, особенно неспецифических областей, связана с созданием условий для образования временных связей. Это достигается активацией высших отделов мозга и распространением этого активного состояния на обширные области коры. По мере укрепления связей эти функции отпадают и участие неспецифических областей становится все более ограниченным.
Способность нейронов включаться в условные реакции в значительной мере зависит от уровня возбудимости. Именно нейроны высокой возбудимости, на которые сходятся сигналы от многих полей рецепции, играют важную роль в механизме образования временных связей. Вероятно поэтому нейронов, проявляющих условные реакции, в первичных проекционных полях оказалось меньше, чем во вторичных, где сопоставляется информация, приходящая по многим путям.
Активность «обучающихся» нейронов изменяется в соответствии со степенью выраженности внешних проявлений условного рефлекса и зависит от факторов, влияющих на эти проявления.
Например, насыщение животного пищей ведет к уменьшению проявлений его пищевых условных рефлексов и резкому снижению условных реакций корковых нейронов.
При формировании временных связей происходит наряду с усложнением путей передачи сигналов образование нейронных цепочек облегченного их проведения.
Исходя из свойств рефлексов было выделено шесть уровней эволюционного совершенствования их временных связей. Первый уровень - суммационный рефлекс, когда в результате повторных, до того не вызывавших реакции раздражений возрастает возбудимость структур синапса и он становится проходимым. Второй - реализует явления так называемого привыкания, когда повторные раздражения, ранее вызывавшие реакцию, приводят к гиперполяризации синапсов и снижают реакцию до полного прекращения. Третий - несамовосстанавливающиеся условные рефлексы с нестойкой временной связью. Четвертый - истинные условные рефлексы со стойкой временной связью. Пятый - комбинационные условные рефлексы, где создаются сложные множественные взаимообусловленные межнейронные связи. Шестой - абстрактно-логические связи, характерные для процессов человеческого мышления (имеет сложную организацию и включает конструкции второй сигнальной системы).
Сложные формы высшей нервной деятельности животных
Приспособительное поведение животных, особенно в сложных ситуациях, нередко принимает такие формы, которые трудно «уложить» в рамки рассмотренных выше актов элементарной условно-рефлекторной деятельности. Наблюдая, как шимпанзе ставит ящик на ящик и, взобравшись на них, достает приманку, подвешенную к потолку клетки, И.П. Павлов говорил: «Когда обезьяна строит свою вышку, чтобы достать плод, то это «условным рефлексом» назвать нельзя. Это есть случай образования знания, уловления нормальной связи вещей». Действующая при этом сложная система условных связей на основе пищевой мотивации обеспечивает целесообразную последовательность действий животного, посредством которых оно достигает удовлетворения своей потребности.
Одной из сложных форм высшей нервной деятельности является так называемое образное или психонервное поведение (И.С. Бериташвили, 1963).
Считают, что в отличие от простых условных рефлексов, требующих повторения сочетаний сигнала и подкрепления, условные связи формирования образов возникают сразу.
Поиски нервного субстрата формирования образов привели к заключению о том, что оно осуществляется с участием подкорковых структур совместной деятельностью проекционных и ассоциативных областей коры мозга, в которой особую роль играют звездчатые нейроны (И.С. Беритов, 1969). Их морфофизиологические особенности позволяют предположить, что звездчатые нейроны с околоклеточной сетью аксонов, образующие своеобразную матрицу в IV слое проекционных полей коры, являются сенсорными, порождающими ощущения; а звездчатые нейроны с аксонами, идущими к вставочным и ассоциативным пирамидным клеткам, связывают через них отдельные компоненты восприятия разных модальностей в целостный образ внешнего мира.
Другая сложная форма высшей нервной деятельности, демонстрирующая «улавливание нормальной связи вещей», о котором говорил И.П. Павлов, была исследована на примере экстраполяционного, или рассудочного, поведения (Л.В. Крушинский, 1977). Как показывает само название, речь идет о способности животного, наблюдавшего течение существенного для него события, «уловить» закономерности его протекания, а когда наблюдение прерывается, то экстраполировать, т.е. «мысленно» продолжить, ход событий и соответственно строить свое поведение без специального обучения. Примером такого поведения в природных условиях может служить случай, описанный известным натуралистом Э. Сетон-Томпсоном, когда ворона, пролетая над ручьем, уходящим в трубу, уронила в него кусок хлеба, который она несла в клюве. Заглянув внутрь, она быстро облетела трубу, дождалась выплывания куска, выловила его и полетела дальше.
Исследования позволили дать сравнительную оценку способности к экстраполяции у исследованных животных разных видов. Так, среди млекопитающих на 1-м месте оказались обезьяны, дельфины и бурые медведи, 2-е место заняли лисицы, волки, собаки, 3-е - песцы и лисицы звероферм, 4-е - кошки, 5-е - мышевидные грызуны, зайцы, 6-е - пасюки, кролики. Среди птиц на 1-м месте - вороновые, на 2-м - хищные птицы, утки, куры, на 3-м - голуби. Способность к экстраполяции у черепах оказалась на уровне между 1-ми 2-м местами у птиц. Лягушки и жабы были совершенно неспособны к обучению, рыб можно было обучить экстраполяции.
Поиски нейрофизиологических механизмов, лежащих в основе экстраполяционного поведения, привели к заключению, что у млекопитающих особую роль в его осуществлении играют лобные отделы мозга, удаление которых делает экстраполяцию невозможной. У птиц, по-видимому, эту роль выполняет особый вырост стриарных тел - вульст, развитие которого коррелирует с показателями экстраполяционной деятельности. У рептилий такая деятельность нарушается при повреждении передних отделов примитивной дорсальной коры мозга. В отношении нейронной организации экстраполяционных процессов предполагают, что они осуществляются при участии нейронов, детектирующих характер изменения действующих раздражителей, свойства которых формировались генетически в онтогенезе.
Нейрофизиологические механизмы аналитико-синтетической деятельности. Процессы синтеза начинаются уже на первичном нервном уровне. На более высоких уровнях обработки сенсорной информации происходит дальнейший синтез афферентных сигналов о признаках объекта.
Аналитико-синтетические процессы обеспечивают поэтапную обработку информации, направленную на восприятие целостного образа объекта.
Изучения межнейронных отношений привели к заключению, что гибкая приспособительная изменчивость интегративных механизмов имеет в своей основе не жестко фиксированные связи между нейронами, а их вероятностное участие в формировании пространственно-временных мозаик возбуждающихся и тормозящихся нейронов. Мощное влияние на организацию аналитико-синтетической деятельности зрительной системы оказывают экологические различия образа жизни. Так, в зрительной коре у кошки преобладает ориентационная избирательность, у приматов -цветоразличение и стереоскопическое зрение, у белки - избирательность к быстрым движениям. Такая оценка происходит на сравнительно высоком уровне интеграции поступившей сенсорной информации, ее сигнальной роли и текущего состояния организма. При этом мономодальные образы синтезируются в полимодальные и с учетом состояния организма, его потребностей и жизненного опыта формируется динамичный целостный образ воспринимаемого явления. В зависимости от того, является ли этот образ показателем благоприятных или опасных событий, возникает и соответствующая эмоциональная окраска восприятия.
Сложными процессами аналитико-синтетической деятельности мозга обусловлено возникновение новых видов приспособительного поведения животных. При этом формируются нервные механизмы объединения афферентных и эффекторных структур в целостную организацию достижения полезного результата.
На основании обширного материала проведенных экспериментов выделены основные принципы организации интегративной деятельности мозга, осуществляемой на базе анализа и синтеза. Это, прежде всего, упорядоченность его функциональной организации, понимаемая как диалектическое сочетание «жесткого каркаса» и динамичных «кружевных переплетений». Другой принцип - мультифункциональность - проявляется в многочисленных примерах участия структур мозга, в частности ассоциативных отделов коры гиппокампа, таламуса и других, в осуществлении разных функций. Принцип иерархической организации обеспечивает оптимальный порядок использования поступающей информации (которая передается на последующие уровни в обобщенном виде) и рациональное управление (когда на высоком уровне задается цель, достигающаяся рядом действий, которые организуются на низших уровнях центральной нервной системы).
Похожие работы


... методов водного и радиального лабиринтов, см. 4.2). Существует даже тенденция считать эти феномены основным проявлением когнитивной деятельности животных (в действительности же в эту категорию входит значительно более широкий круг индивидуально-приспособительных реакций животных, в том числе довербальные понятия, способность к усвоению и использованию символов, которая была обнаружена при ...
... и левой руки у праворуких при тактильном узнавании предметов дигаптическим методом. Обнаружен половой диморфизм по соотношению длин левой и правой височных плоскостей.[18] 3. Заключение. Рассмотрение исследований межполушарной асимметрии показывает, что она проявляется практически у всех животных. Более интересно это выявлено у человека. Исследование показали, что полушария различны не только ...
... эмиссионная томография (ОЭТ); позитронная эмиссионная томография (ПЭТ). Весь этот комплекс методов позволяет проводить неинвазивное изучение структуры и функций мозга. Психофизиологическое изучение психических процессов и состояний Принципы кодирования информации в нервной системе Сегодня можно говорить о нескольких принципах кодирования в нейронных сетях. Одни из них достаточно просты и ...




... -третьим). Полученные результаты, в целом, позволяют сделать вывод о том, что личностное развитие подростков связано с развитием интеллекта и когнитивных структур. ЗАКЛЮЧЕНИЕ Настоящее диссертационное исследование посвящено экспериментальному изучению старших подростков музыкальных школ и проведено в контексте направления когнитивной психологии, разрабатываемого Н.И. Чуприковой и ее коллегами ...
0 комментариев