КЛАСС ХРЯЩЕВЫЕ РЫБЫ – CHONDRICHTHYES
Характеристика класса и системы
Морские рыбы (лишь несколько видов живут в пресных водоемах). Есть плакоидные чешуи, или кожа голая. У современных форм скелет хрящевой, кожные окостенения отсутствуют. У пластиножаберных развиваются амфицельные хрящевые позвонки, сдавливающие хорду; у цельиоголовых образуются верхние и нижние дуги, но тела позвонков не развиваются, а в оболочке хорды откладываются известковые соли. Череп амфистиличный или гиостиличный, у цельноголовых – аутостиличный. Хвост гетероцеркальный. Есть 5–7 пар наружных жаберных щелей или с каждой стороны они прикрыты общей кожистой жаберной крышкой. Плавательного пузыря нет. В кишечнике хорошо развит спиральный клапан. В сердце есть артериальный конус. Видимо, для всех видов характерно внутреннее оплодотворение; у самцов обособляются участки брюшных плавников – птеригоподий – выполняющие функцию копулятивных органов; яйца крупные в плотной рогоподобной капсуле. У многих видов развивается яйцеживорождение и даже живорождение. Быстрые и ловкие пловцы, питающиеся крупной подвижной добычей, или относительно малоподвижные формы, поедающие преимущественно донных беспозвоночных. Размеры варьируют: от мелких скатов длиной около 15 см до акул длиной в 15–20 м.
Ныне живущие хрящевые рыбы разделяются на два подкласса: пластиножаберные – Elasmobranchii и цельноголовые – Holocep-hali. Пластиножаберные распадаются на два надотряда: акул Selacho-morpha и скатов – Batomorpha. В подклассе цельноголовых один отряд химерообразные – Chimaeriformes.
Класс Chondrichthyes – Хрящевые рыбы
+ Подкласс Cladoselachii – Примитивные акулы + Подкласс Xenacanthida – Ксенакантиды
Подкласс Elasmobranchii – Пластиножаберные, или акуловые рыбы Надотряд Selachomorpha – Селяхоидные, или Акулы Отряд Chlamydoselachiformes – Плащеносцеобразные Отряд Hexanchiformes – Многожаберникообразные Отряд Heterodontiformes – Разнозубообразные Отряд Lamniformes – Ламнообразные Отряд Carcharhiniformes – Кархаринообразные, или Пилозубообразные Отряд Squaliformes – Катранообразные Отряд Pristiophoriformes – Пилоносообразные Отряд Squatiniformes – Скватинообразные Надотряд Batomorpha – Батоидные, или Скаты Отряд Pristiformes – Пилорылообразные Отряд Rhinobatiformes – Рохлеобразные Отряд Raiiformes – Ромбообразные Отряд Myiiobatiformes – Орлякообразные Отряд Torpediniformes – Гнюсообразные Подкласс Holocephali – Цельноголовые – f – Отряд Bradyodonti – Брадиодонты Отряд Chimaeriformes – Химерообразные
Краткая характеристика современных хрящевых рыб
ПОДКЛАСС ПЛАСТИНОЖАБЕРНЫЕ – ELASMOBRANCHII
Передний конец морды вытянут в более или менее выраженное рыло – рострум. С каждой стороны открывается по 5–7 жаберных щелей. Обычно есть брызгальце – остаток щели между челюстной и подъязычной дугами. Череп амфистиличный, или гиостиличный.
Жаберные лепестки имеют вид пластин, расположенных на кожистых межжаберных перегородках между щелями.
НАДОТРЯД АКУЛЫ – SELACHOMORPHA
Тело удлиненное, более или менее торпедообразное. Мощный хвостовой стебель заканчивается большим гетероцеркальным хвостовым плавником. Жаберные щели расположены по бокам головы. На челюстях обычно расположено большое число мощных острых зубов хва-тательно-режущего типа.
Отряд Плащеносцеобразные акулы – Chlamydoselachiformes
Включает один вид – плащеносную акулу – Chlamydoselachus ап-guineus – длиной 1,2–2 м. По бокам головы 6 пар жаберных щелей. Перепонки первой щели соединяются на горле, образуя широкую кожистую лопасть – «плащ». Живут около дна на глубинах 400–1200 м в умеренных и теплых морях. Всюду малочисленны. Питаются рыбами и головоногими моллюсками. Яйцеживородящи.
Отряд Многожаберниковообразные акулы – Hexanchiformes
Крупные акулы длиной до 4–8 м, с 6–7 парами жаберных щелей, но без кожистой складки на горле. Держатся на больших глубинах в тропиках и субтропиках; с теплыми водами проникают и в умеренные широты. Питаются главным образом рыбой. Яйцеживородящи; у одной самки находили 50–108 эмбрионов. Местами – объекты промысла. Всего 3 рода и 5 видов. Ископаемые представители отряда встречаются с юры.
Отряд Разнозубообразные акулы – Heterodontiformes
Акулы длиной до 1,5 м, с массивной головой, с 5 парами, как и у всех следующих отрядов, жаберных щелей. 4 вида населяют теплые прибрежные воды Тихого и Индийского океанов. Зубы в передней части челюстей мелкие и острые, а на остальных участках – мощные и притуплённые, дробящие. Питаются крабами, моллюсками, морскими ежами, легко дробя плотные покровы добычи мощными зубами. Скоплений не образуют. Яйцекладущи. Самка выметывает 1–2 яйца размером 10x5 см с двумя спиральными складками на поверхности яйцевой оболочки. Через 7 месяцев после откладки яйца из него вылупляется акулка длиной около 20 см.
Отряд Ламнообразные акулы – Lamniformes
Включает 6 семейств с примерно двумя десятками довольно крупных видов. Своеобразна морская лисица – Alopias vulpes, достигающая 6 м длины и массой до 450 кг. Ударами длинного хвоста она иногда оглушает добычу или, кружась вокруг стаи, заставляет рыб сбиться в кучу, а затем бросается На них и заглатывает. Сельдевые акулы длиной 2,5–3,5 м населяют умеренные воды Мирового океана. Прекрасные пловцы, держатся стаями по 5–30 особей, преследуя стайных пелагических рыб – сельдей, сардин, лососей. Серо-голубые акулы, или мако, достигают 4–5 м длины и населяют тропические воды. Острые треугольные зазубренные зубы у крупных особей достигают 7–10 см длины. Питаются крупными рыбами. Опасны для человека. Акула-людоед, или «белая смерть», – Carcharodon carcharias достигает длины 6–7 м и массы 2–3 т, становится половозрелой при длине 4 м. Очень агрессивны. Питаются крупными рыбами, в том числе и акулами. Рыб длиной до 2–2,5 м заглатывают целиком, а более крупных разрывают на куски мощными 5–7-сантиметровыми зазубренными зубами. Один из наиболее опасных видов: отмечены многочисленные случаи нападения на человека. На глубинах 1–1,5 км встречается достигающая длины 4 м акула-домовой – Scaphanorhynchus owstoni, питающаяся разнообразными донными животными. Ее длинное уплощенное рыло имеет форму клина и, вероятно, используется для раскапывания грунта. Ламнообразные яйцеживородящи. Самка морской лисицы рождает 2–4 акулят длиной 1–1,5 м; сельдевая акула – 3–5 детенышей длиной 50–70 см. Часть видов имеет некоторое промысловое значение.
Отряд Кархаринообразные, или Пилозубые, акулы – Carcharhiniformes
Объединяет до 150 видов мелких, средних и очень крупных акул, группируемых в 7 семейств. Сюда относятся два самых крупных вида ныне живущих рыб: китовая акула Rhincodon typus, достигающая длины до 20 м, и гигантская акула – Cetorhinus maximus длиной до 12–15 м. Первая встречается поодиночке в тропических, а вторая – группами до 20–30 особей преимущественно в умеренных водах. Оба гиганта питаются планктонными ракообразными, моллюсками и мелкими стайными рыбами. На жаберных дугах густо сидят длинные тычинки, преграждающие доступ к жаберным щелям и образующие цедильный аппарат. Напавшая на скопление планктонных организмов акула медленно плывет с открытой пастью, пропуская через ротовую полость воду и выбрасывая ее через жаберные щели; планктон задерживается цедильным аппаратом и заглатывается. Зубы у этих акул мелкие, служат не для хватания, а для «запирания» добычи во рту. За час акула профильтровывает до 1,5–2 тыс. кубометров воды. Зимой гигантские акулы погружаются на большие глубины и держатся у дна, видимо, находясь в состоянии, подобном спячке. Китовая акула откладывает яйца; гигантская акула, видимо, яйцеживородяща.
Семейство серых акул, объединяющее до 60 видов, включает серых акул длиной до 3,6 м, распространенных в прибрежных водах тропической области; проникают в крупные реки Африки, Азии и Южной Америки, поднимаясь на 200–300 км от устья; один вид постоянно живет и размножается в пресном озере Никарагуа. Всеядны, отмечены нападения на людей. Более крупная и подвижная тигровая акула – один из наиболее опасных для человека видов; встречается в прибрежных районах и в открытом море. К серым акулам близки своеобразные акулы-молоты. Их голова сильно уплощена и имеет два больших боковых выроста, на наружных краях которых расположены глаза, а несколько отступя от них – большие ноздри. 1 видов этого семейства имеют длину до 3–6 м. Это быстрые, неутомимые и маневренные пловцы, встречающиеся как в открытых, так и в прибрежных водах тропиков. Питаются донными и пелагическими беспозвоночными, различными рыбами, в том числе и крупными; известны случаи нападения иа людей. Мелкие кошачьи и куньи акулы тоже относятся к пилозубым акулам. Несколько десятков видов их встречается в прибрежных водах, немногие живут на глубинах до 600–1500 м.
Среди отряда пилозубообразных встречаются яйцекладущие, яйце-живородящие и живородящие формы. Так, самки кошачьих акул откладывают от 2 до 20 яиц в роговых капсулах с длинными роговыми нитями. Самка тигровой акулы рождает до 30–50 детенышей длиной в 45–50 см, а самки акулы-молот – до 30–40 детенышей такой же длины. Некоторые виды служат объектами промысла или спортивного рыболовства.
Отряд Катраиообразиые – Squaliformes
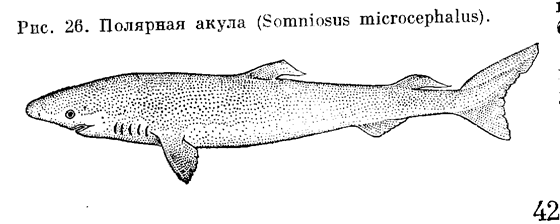
Около двух десятков видов колючих акул имеют длину до 1, редко до 2 м. Часть видов характерна для прибрежных вод глубиной до 200 м, другие встречаются на глубине 1 км и глубже. Питаются рыбами, ракообразными и различными донными беспозвоночными. Некоторые ведут стайный образ жизни. Служат объектом промысла, иногда причиняют существенный ущерб рыболовству: объедают рыбу в сетях, разрывают снасти. Катраны становятся половозрелыми, достигнув 1 м длины в возрасте около 19 лет. Яйцеживородящи. Самка через 6–7 месяцев после оплодотворения выметывает от 12 до 30 детенышей длиной 20–26 см. Живут свыше 30 лет. Сюда же относят акул, не имеющих шипов. В Северной Атлантике живет достигающая 6,5 м длины и массой около тонны полярная акула Somniosus tnicrocephalus. Летом она держится на больших глубинах, а зимой поднимается в поверхностные слои. Прожорливый хищник, поедающий различных рыб и беспозвоночных, трупы китов и тюленей. Весной на большой глубине самка выметывает до 500 эллипсовидных яиц в мягкой оболочке, диаметром до 8 см. В теплых водах встречаются карликовые акулы, не превышающие в длину 20–25 см. Они живут в открытом океане, ночью поднимаясь к поверхности, а днем уходя в глубину. Всю нижнюю поверхность тела этих акулок густо усеивают мелкие люминесцентные органы – фотофоры, излучающие при возбуждении рыбы бледно-зеленый свет. Питаются, видимо, мелкими головоногими моллюсками.
Отряд Пилоносообразные – Pristiophoriformes
Акулы этого отряда, длиной до 1,5 м, живут в теплых, преимущественно прибрежных водах Тихого и Индийского океанов. Их внешний облик очень своеобразен и похож на пилу-рыбу. Форма тела акулья, 5 или 6 пар жаберных щелей расположены по бокам головы, рыло превратилось в длинный и плоский мечевидный отросток, несущий по бокам крупные зубы; примерно посередине отростка расположена пара длинных подвижных осязательных усиков. Это медлительные рыбы, живущие у дна и раскапывающие «пилой» грунт в поисках различных беспозвоночных; ловят и мелких рыб. Самка рождает до 12 акулок, у которых уже хорошо сформированные зубы пилы покрыты мягкой, спадающей после рождения оболочкой.
Отряд Скватинообразные, или морские ангелы, – Squatiniformes
Донные акулы, приобретшие некоторое сходство со скатами. Тело уплощено в спинно-брюшном направлении, рыло тупое, округлое. Грудные и, в меньшей степени, брюшные плавники увеличены. Как и у типичных акул, жаберные щели расположены по бокам головы. Достигают длины 2,5 м и массы 100 кг. Распространены в тропиках и умеренной зоне. Живут на дне, питаясь мелкими рыбами и беспозвоночными, Обычно держатся на мелководьях. Встречаются яйцеживородящие и яйцекладущие виды.
НАДОТРЯД СКАТЫ – BATOMORPHA
Тело уплощено в спинно-боковом направлении. У многих форм хвостовой стебель развит слабо, а лопасть хвостового плавника может редуцироваться. Грудные плавники сильно увеличены, с широкими основаниями. 5 пар жаберных щелей расположены на брюшной стороне. Зубы обычно имеют вид невысоких призм, плотно прилегающих друг к другу и образующих мощную «терку».
Отряд Пилорылообразные скаты, или Пилы-рыбы, – Pristiformes
Отряд состоит из одного рода и 7 видов пил-рыб. Достигают 5–6 м длины; тело слабо уплощено, грудные плавники относительно невелики. Жаберные щели расположены на нижней поверхности головы.
Удлиненное мечеобразное рыло по краям усажено крупными зубами, но в отличие от пилоносых акул не несет осязательных усиков. Живут на мелководьях тропических и субтропических морей, иногда заходят в нижние участки рек. Хорошие пловцы. Питаются мелкими стайными рыбами и различными донными беспозвоночными. Яйцеживородящи. Зубы на «пиле» детенышей показываются из кожи только после рождения.
Отряд Рохлеобразные – Rhinobatiformes
В отряде около полусотни видов; длина тела от 70 см до 3 м. Тело уплощено, но мощный хвостовой стебель заметно не обособлен от туловища и обычно несет хорошо развитую хвостовую лопасть, широкие плавники имеют небольшую длину, рыло удлиненное. Живут у дна в прибрежных районах тропиков и субтропиков, иногда проникают в опресненные эстуарии. Плавают медленно, используя в качестве движителя хвост, а не грудные плавники, как скаты описанных ниже отрядов. Питаются донными беспозвоночными, хватают мелких рыб. Местами причиняют ущерб устричным банкам. Яйцеживородящи.
Отряд Ромбообразные, или Ромботелые, скаты, – Rajiformes
Сюда относят более ста видов трех семейств, характеризующихся сильно уплощенным, более или менее ромбовидным телом, заостренным рылом и четко отграниченным от туловища тонким хвостовым стеблем, заканчивающимся маленьким хвостовым плавником. У большинства видов в коже разбросаны крупные плакоидные чешуи, имеющие вид погруженной в кожу округлой пуговицы с выдающимся над кожей мощным, слегка изогнутым назад зубом. Держатся на дне, населяют холодные и умеренные воды, в тропиках встречаются лишь на больших глубинах. Длина от 30 см до 2 м. Днем обычно лежат на грунте или слегка закапываются в него, ночью плавают у дна. Основной способ плавания, как и у нижеописываемых отрядов, – волнообразные движения грудных плавников. Питаются мелкой рыбой, донными ракообразными, моллюсками, иглокожими. На рыб наплывают сверху, прижимают их телом к грунту и потом поедают. Самки откладывают одновременно только 1–2 яйца, а за растянутый период яйцекладки – несколько десятков яиц в роговых капсулах с усиками. Развитие длится до 4–6 месяцев. У метровых скатов длина яйца достигает 6–10 см, а выходящий из яйца молодой скат имеет длину 10–15 см.
Отряд Орлякообразные, или Хвостоколообразные, – Myliobatiformes
В отряд входит около ста видов скатов различной величины. Тело ромбовидное или, реже, овальное. Резко отграниченный от тела хвостовой стебель имеет вид постепенно утончающегося упругого стержня без следов спинных и хвостового плавников. У многих видов примерно посередине хвостового стебля находится одна длинная уплощенная роговая игла с острой вершиной и зазубренными краями. На нижней поверхности иглы проходит бороздка, выстланная железистыми клетками, выделяющими ядовитый слизистый секрет. При нападении врага хвостокол бьет его хвостом, нанося иглой глубокие рваные раны; попадающий в рану секрет вызывает резкую боль. Хвостоколы обычно лежат на песчаном или илистом дне, полузакопавшись в грунт. Если человек случайно наступит на ската, то он сразу же начинает отбиваться хвостом. При попадании в раны даже небольшого количества яда помимо резкой боли падает кровяное давление, начинается рвота, мышечные параличи; известны и смертельные исходы. Обычны в тропических и субтропических морях, редки в умеренных широтах; у черноморского побережья Советского Союза встречается морской кот длиной до 1, редко до 2–2,5 м. Живут на мелководьях, но есть и глубоководные формы. Заходят в устья рек. Несколько видов речных скатов живут в бассейне Амазонки и в других реках Бразилии и Парагвая.
Близкие к хвостоколам орляки, достигающие длины 4,5 м и ширины тела 2,5 м, прекрасно плавают и питаются не только донными животными, но и пелагическими. Хвостовой иглы не имеют. К орлякам близко семейство рогатых скатов или мант, насчитывающее около десятка видов. Самый мелкий вид едва превышает в ширину 1 м, а гигантская манта, или морской дьявол, – Manta birostris, особенно многочисленный у тропических берегов Америки, достигает в размахе 7 м и массы 2 т. Передние части грудных плавников образуют выступающие перед глазами выросты. Ведут пелагическую жизнь, плавая в верхних слоях воды. Движутся быстро, «взмахивая» заостренными крылообразными грудными плавниками. Могут выпрыгивать из воды на высоту до 1,5–2 м; звук падения громадного плоского тела слышен за несколько километров. Полагают, что таким способом манты избавляются от кожных паразитов. Охотятся за стайными пелагическими рыбами. Предполагают, что движения головных выростов создают ток воды, облегчающий засасывание мелкой рыбы в широкий, находящийся почти на конце головы рот.
Скаты этого отряда яйцеживородящи или живородящи. Одновременно рождают 4–12 детенышей; гигантская манта – только одного массой до 15–20 кг и в размахе плавников – более 1,2 м.
Отряд Гнюсообразные, или Электрические скаты, – Torpediniformes
Три семейства примерно с 40 видами, распространенными в прибрежных районах тропических и субтропических морей всего земного шара. Размеры от мелких – длиной в 12–15 см – до крупных – длиной до 2 м и массой до 100 кг. Тело округлое, короткий хвостовой стебель широк у основания, заканчивается небольшой хвостовой лопастью и несет один-два маленьких спинных плавничка. Плакоидных чешуи и шипов нет. В отличие от других скатов часто ярко окрашены. По бокам головы расположены парные электрические органы, образованные видоизмененной мышечной тканью. Их масса может составлять до Ус массы тела. В электрических органах накапливается животное электричество. Разряд осуществляется произвольно, под влиянием импульсов головного мозга. Одиночный разряд длится 0,003–0,05 с, но обычно скат производит серию быстро следующих друг за другом разрядов: до 20–30. Напряжение при разряде может достигать 60–300 вольт при силе тока до 5 ампер.
Электрические скаты малоподвижны, плавают плохо; обычно лежат на дне, полузарывшись в песок или ил. Когда добыча подплывает к скату, следует короткая серия сильных разрядов, оглушающих жертву. Добычей крупных скатов становятся рыбы массой до 2–3 кг; питаются и донными беспозвоночными. Скаты используют электрические разряды и при обороне; наносят сильные электрические удары, если человек случайно наступит на ската или попытается его схватить. Кроме сильных разрядов, производимых при нападении и защите, скаты создают вокруг себя электрическое поле, генерируя слабые разряды напряжением 0,2–2, редко до 10 и даже 50 В и силе тока около 1 А, такие разряды следуют с частотой 35–300 раз в 1 с. Вторжение любого предмета искажает однородность электрического поля. Восприятие чувствительными рецепторами нарушений электрического поля дает возможность судить о приближении добычи или врага. Видимо, преобладание подобного способа рецепции обусловило появление нескольких видов слепых скатов. Яйцеживородящи; вынашивание зародышей продолжается до года. Одновременно рождается 4–10 детенышей.
ПОДКЛАСС ЦЕЛЬНОГОЛОВЫЕ – HOLOCEPHALI
Конец морды обычно более или менее закруглен. Развиваются кожистые жаберные крышки, и поэтому с каждой стороны головы видно только по одному жаберному отверстию. Брызгальце отсутствует. Череп аутостиличный. Зубы сливаются в мощные зубные пластинки, несколько напоминающие мощные резцы грызунов. В связи с образованием кожистых жаберных крышек кожистые межжаберные перегородки заметно редуцируются.
Отряд Химерообразные – Chimaeriformes
Вальковатое тело несколько сжато с боков и заметно утончается к хвосту. У высокого первого спинного плавника имеется острый шип; у некоторых видов у его основания лежит ядовитая железа. Хвост гетероцеркальный или продолжается в виде длинного утончающегося бича. Морские, преимущественно глубоководные формы. Распространены в умеренных и теплых водах Мирового океана. Лишь немногие виды образуют промысловые скопления. Держатся у дна; мощными зубами легко разгрызают панцири ракообразных и иглокожих, крепкие раковины моллюсков. Плавают за счет волнообразных движений грудных плавников и боковых изгибов хвоста. Длина от 60 см до 1,5–2 м. Размножение растянутое. Одновременно самка откладывает лишь одно-два крупных яйца, каждое из которых заключено в удлиненную овальную роговую капсулу с извитым нитевидным придатком на конце. Яйца падают на каменистое дно или повисают на водорослях. Развитие продолжается 9–12 месяцев. По бокам головы развивающегося зародыша образуются нитевидные кожные выросты – наружные «жабры», которые, вероятно, облегчают всасывание желтка и, возможно, получение кислорода. Перед вылуплением эти «жабры» исчезают, и покидающая оболочку молодая химера отличается от взрослых лишь размерами.
Экологическая дифференцировка позволила хрящевым рыбам занять основные экологические ниши в морях и успешно конкурировать с более разнообразным классом костных рыб. Распространение по всем морям и значительное число живущих ныне видов позволяют считать хрящевых рыб, несмотря на эволюционную древность многих групп и относительную примитивность морфологических особенностей, вполне конкурентноспособными морскими позвоночными животными. Очень малое число видов, постоянно живущих в пресных водоемах, и их ограниченные ареалы свидетельствуют о том, что в пресных водоемах хрящевые рыбы не смогли противостоять давлению костных рыб. Нужно выяснить причину такой ситуации. Есть основания предполагать, что это определяется своеобразными биохимическими особенностями хрящевых рыб.
Особенности организации хрящевых рыб
Покровы. Кожа и ее производные. Кожа хрящевых рыб образована многослойным эпидермисом и подстилающим его соединительнотканным слоем – кориумом. В эпидермисе расположены многочисленные одноклеточные железы, слизистый секрет которых тонким слоем покрывает все тело. Клетки нижних слоев эпидермиса содержат пигменты. Специальные пигментные клетки расположены и в кориуме. В совокупности они создают свойственный каждому виду тип окраски. Некоторые скаты способны менять окраску в зависимости от цвета грунта, на который они легли. Это достигается перемещением пигмента из отростков в тело клетки и наоборот либо сжатием и расширением всей пигментной клетки.
В коже хрящевых рыб образуются плакоидные чешуи. Чешуя такого типа состоит из лежащей в кориуме округлой или ромбической пластинки и сидящего на ней шипа, вершина которого через эпидермис выдается наружу. Внутри чешуи имеется полость, заполненная насыщенной кровеносными сосудами мякотью. Вся чешуя состоит из близкого к кости, но более плотного вещества – дентина, образованного клетками кориума; вершину шипа покрывает чехлик из еще более плотного костеподобногс вещества – эмали, образуемой клетками базального слоя эпидермиса. У акул чешуи более или менее равномерно покрывают все тело, у скатов в коже разбросаны крупные чешуи, между которыми в беспорядке рассеяны мелкие чешуйки. У части скатов чешуи редуцируются. У цельноголовых кожа голая, лишь у некоторых видов на ограниченных участках встречаются сильно измененные плакоидные чешуи.
Плакоидные чешуи крупной и усложненной формы располагаются на челюстях, превращаясь в зубы. Шип при этом увеличивается, возрастает и толщина эмали. При изнашивании или поломке зуб отпадает, и ему на смену вырастает новый; замена зубов может проходить в течение всей жизни рыбы. У цельноголовых отдельные зубы сливаются б зубные пластинки. Зубы всех позвоночных животных построены по сходному типу; в сущности это измененные плакоидные чешуи предков.
Кожа хрящевых рыб обеспечивает им достаточно надежную механическую защиту. Но она проницаема для воды, частично и для растворенных в ней веществ.
Скелет и мышечная система. У ныне живущих хрящевых рыб в скелете отсутствуют кости. Внутренний скелет полностью хрящевой; его отдельные участки могут пропитываться солями извести и приобретать значительную твердость. Скелет подразделяется на осевой, скелет черепа, скелет парных и непарных плавников и скелет поясов парных плавников.
Осевой скелет пластиножаберных рыб состоит из позвоночного столба, разделяющегося на туловищный и хвостовой отделы. Он образован многочисленными позвонками. Тело позвонка спереди и сзади вогнуто. Такие позвонки называют двояковогнутыми, или амфицельными. В центре тела позвонка есть канал, в котором проходит хорда. Она пронизывает весь позвоночный столб и имеет четкообразный вид: расширения в полостях между телами соседних позвонков и сужения – в телах позвонков; реальной опорной функции хорда уже не несет. Над телами позвонков расположены верхние дуги, имеющие широкое основание и суженную вершину. В месте слияния правой и левой верхних дуг образуются короткие остистые отростки. Между верхними дугами расположены вставочные пластинки, вместе с верхними дугами образующие стенки хрящевого канала, в котором лежит спинной мозг.
В туловищном отделе слабо развитые нижние дуги образуют короткие поперечные отростки, к которым причленяются очень короткие хрящевые ребра. В хвостовом отделе разрастающиеся нижние дуги правой и левой стороны смыкаются друг с другом; между ними располагаются небольшие нижние вставочные пластинки; так образуется гемальный канал, в котором проходят хвостовая артерия и вена.
В ходе онтогенеза в соединительнотканной оболочке хорды вначале в каждом сегменте тела образуются хрящевые зачатки: два парных верхних и два парных зачатка нижних дуг. Далее вокруг хорды начинают развиваться сжимающие хорду кольцевидные хрящи – тела позвонков. Основания задних верхних и нижних дуг разрастаются вокруг тела позвонка, увеличивая его толщину. Вершины верхних дуг срастаются; нижние дуги в туловищном отделе образуют поперечные отростки, а в хвостовом отделе сливаются, замыкая гемальный канал. Передние пары зачатков верхних дуг превращаются в верхние вставочные пластинки, а передние пары зачатков нижних дуг в хвостовом отделе образуют нижние вставочные пластинки.
У цельноголовых рыб тела позвонков не образуются. Соединительнотканная оболочка хорды уплотняется и в ней возникают кольцевые обызвествления, внешне напоминающие тела позвонков. Передняя пара хрящевых зачатков развивается в верхние дуги, опирающиеся на оболочку хорды; задняя пара превращается во вставочную пластинку. В хвостовом отделе развиваются нижние дуги и нижние вставочные пластинки. Ребра отсутствуют.
Череп хрящевых рыб подразделен на два отдела: мозговой и висцеральный.
Мозговой череп пластиножаберных рыб представляет сплошную хрящевую коробку, со всех сторон окружающую головной мозг; в крышке черепа остается затянутая соединительной тканью небольшая фонтанель. Передний конец черепа вытянут в рострум, поддерживающий рыло. У основания рострума с черепом срастаются парные обонятельные капсулы, а в заднебоковые стенки врастают слуховые капсулы. Затылочный отдел прободен затылочным отверстием, через которое проходит начальная часть спинного мозга. На боковых поверхностях черепа находятся глубокие углубления – глазницы, защищающие глаз сверху и с боков. Череп плати-базальный: основание широкое, стенки глазниц разобщены и между ними лежит головной мозг.
Висцеральный скелет возник из преобразованных жаберных дуг, поддерживавших у предков глоточную область. Его обычно подразделяют на челюстную дугу, подъязычную дугу и жаберные дуги.
Челюстная дуга образована двумя парными хрящами. Левый и правый элементы каждой пары спереди соединяются друг с другом. Функцию верхней челюсти выполняет небно-квадрат-ный хрящ, образующий челюстной сустав с нижней челюстью – меккелевым хрящом. На верхней и нижней челюстях сидят зубы. У многих акул спереди челюстной дуги лежат 1–2 пары маленьких губных хрящей, рассматриваемые как остатки двух редуцированных жаберных дуг; они свидетельствуют о том, что челюстная дуга образовалась из третьей жаберной дуги предков.
Подъязычная дуга лежит сразу за челюстной. Она состоит из двух парных и одного непарного хряща. Крупный верхний парный хрящ – подвесок или гиомандибуляре – у пластиножаберных рыб верхним концом при помощи сустава и связок подвижно прикрепляется к слуховому отделу мозгового черепа, а его нижний конец подвижно сочленяется как с обоими элементами челюстной дуги в области челюстного сустава, так и с нижним парным элементом подъязычной дуги – гиоидом. Гноиды правой и левой сторон соединяются друг с другом через небольшой непарный хрящ – копулу. Такой тип соединения челюстной дуги с мозговым черепом – через верхний элемент подъязычной дуги – называется гиостилией. Он характерен для подавляющего большинства ныне живущих пластиножаберных. У наиболее древних и примитивных акул отмечена амфистилия: челюстная дуга прикрепляется к мозговому черепу через подвесок и, кроме этого, отросток небно-квадратного хряща непосредственно сочленяется с основанием мозгового черепа. Для скатов характерна типичная гиостилия, но нижний элемент подъязычной дуги – гиоид – редуцируется. Гиостилия, обеспечивающая подвижность всей челюстной дуги и широкий раствор рта, выгодна при схватывании и заглатывании крупной добычи.
Позади подъязычной дуги расположены пять пар жаберных дуг, отделенных друг от друга жаберными щелями. Первая жаберная щель расположена позади подъязычной и перед первой жаберной дугой, а пятая – перед пятой жаберной дугой. Каждая жаберная дуга состоит из подвижно сочленяющихся друг с другом четырех парных элементов и замыкающим дугу снизу непарным пятым элементом – копулой. У большинства акул и скатов копулы сливаются в единую пластинку, что укрепляет нижнюю часть жаберного аппарата. Верхние элементы каждой жаберной дуги эластичными связками соединяются с позвоночным столбом. От переднего края всех жаберных дуг отходят палочковидные хрящи – жаберные тычинки, перегораживающие жаберную щель и удерживающие пищу в полости глотки, когда вода проходит через жаберные щели наружу. Тычинки особенно длинны, тонки и густо расположены у планктоноядных видов, образуя настоящий цедильный аппарат. По заднему краю двух центральных элементов каждой жаберной дуги, а также по заднему краю подвеска и гиоида укрепляются тонкие стержневидные хрящи – жаберные лучи; они лежат в толще кожистых межжаберных перегородок, поддерживая и укрепляя их.
В отличие от пластиножаберных в черепе цельноголовых рострума нет или он развит слабо и обонятельные капсулы прирастают к мозговой коробке спереди. Небно-квадратный хрящ срастается с основанием мозгового черепа; его задний край образует выступ, к которому суставом прикрепляется нижняя челюсть – меккелев хрящ. Не несущая функции прикрепления челюстей и с ними не связанная подъязычная дуга по строению почти неотличима от жаберных дуг. Жаберные лучи подъязычной дуги сильно развиты и поддерживают кожистую жаберную крышку, прикрывающую жаберные щели. Весь жаберный аппарат располагается под черепной коробкой, а не позади нее, как у пластиножаберных. Зубы сливаются в зубные пластинки – мощные орудия грызения. Череп тропи-базального типа. Аутостилия и образование зубных пластинок – приспособления, укрепившие челюстной аппарат и позволившие разгрызать прочные раковины моллюсков, панцири ракообразных и иглокожих.
Скелет непарных плавников образован варьирующими по величине рядами палочковидных хрящей – радиалий, погруженных в мускулатуру и проникающих в основание плавника. Иногда часть радиалий сливается в крупные пластинки. Лопасть самого плавника поддерживают эластотрихии – тонкие лучи из рогоподобного вещества, продуцируемого клетками кожи. Хвостовой плавник у большинства хрящевых рыб гетероцеркальный: верхняя лопасть значительно больше нижней и в нее заходит конец позвоночного столба. Скелетную основу хвостового плавника образуют удлиненные верхние и нижние дуги позвонков и ряддорзо-спинальных и вентро-спинальных нерасчлененных хрящей. Кожные лопасти плавника поддерживаются эластотрихиями. У части скатов и химер лопасти хвостового плавника значительно уменьшаются или вовсе исчезают.
Пояс передних конечностей имеет вид хрящевой дуги, лежащей в толще мускульной стенки позади жаберного отдела. У акул и химер он никак не связан с осевым скелетом, у скатов – в связи с увеличением размеров грудных плавников – верхние части хрящевой дуги пояса сочленяются с позвоночным столбом. На боковой поверхности пояса с каждой стороны находится вырост с сочленованной поверхностью – место причленения скелета плавника. Часть пояса, лежащая выше сочленованного выроста, называется лопаточным отделом, а нижележащая – коракоидным. В основании скелета грудного плавника лежат три массивных уплощенных базальных хряща, суженными вершинами причленяющиеся к сочленованному выросту пояса. К дистальньш концам базальных хрящей прикрепляются радиальные хрящи, каждый. из которых состоит из 2–3 члеников. Дистальная часть плавника поддерживается эластотрихиями.
Тазовый пояс хрящевых рыб образован хрящевой пластинкой, лежащей в мускулатуре брюшной стенки непосредственно перед клоакой. К его боковым поверхностям причленяется скелет брюшного плавника. Он состоит из одного удлиненного базального элемента, к наружной поверхности которого прикрепляется ряд радиальных хрящей; первый из них более крупный. Остальная часть плавника поддерживается многочисленными эластотрихиями. У самцов хрящевых рыб удлиненный базальный элемент продолжается за пределы лопасти плавника и служит скелетной основой копулятивного выроста – птеригоподия.
Соматическая мускулатура отчетливо сегментарна и состоит из мышечных сегментов – миомеров. Миомеры разделяются друг от друга соединительнотканными перегородками – миосептами, образующими сложную ломаную линию в виде лежащей на боку латинской буквы W, средний угол которой вершиной направлен вперед и лежит на горизонтальной соединительнотканной перегородке, делящей все миомеры на спинной и брюшной отделы. На отдельных участках тела метамерия нарушается: перестройка миомеров или их участков приводит к дифференцировке отдельных мышц. Так обособляются глазные, наджаберные и поджаберные мышцы и мышцы парных плавников.
Висцеральная мускулатура – это слои гладких мышц, окружающих пищеварительную трубку. У хрящевых рыб в области челюстных и жаберных дуг гладкие мышечные волокна висцеральной мускулатуры замещаются поперечнополосатыми и формируют довольно сложно дифференцированные группы мышц, управляющих движениями челюстей и жаберных дуг. Участки миомеров соматической мускулатуры в этой области еще сохраняются, но развиты относительно слабо.
Характерной особенностью мускулатуры хрящевых рыб служит ее относительная автономность – сохранение способности сокращаться при нарушении связи с центральной нервной системой. С этой особенностью, физиологические механизмы которой еще достаточно не выяснены, связана хорошо известная необычайная «живучесть» хрящевых рыб. Так, обезглавленная акула довольно долго сохраняет способность к плаванию; акула с вырезанными внутренностями не только нормально плавает, но бросается на добычу и проглатывает ее. Своеобразен и химический состав мышц хрящевых рыб. В них необычайно велико содержание мочевины: до 1,5–2,8% у морских и около 0,7% у пресноводных; у костистых рыб количество мочевины не превышает 0,02–0,03%. Такая высокая уремия, губительная для прочих позвоночных, связана со своеобразным осморегу-ляторным механизмом акуловых рыб, о чем подробнее сказано ниже. Мышцы хрящевых рыб содержат относительно много солей тяжелых металлов: содержание Ti в 10 раз, Мп в 20 раз, Со, № в 40 раз выше, чем в мышцах костистых рыб. Физиологическое значение этого феномена не выяснено.
Особенности мускулатуры и всего двигательного аппарата, видимо, обусловливают повышенные резервы мощности хрящевых рыб. Так, в экспериментах акулы заметно не снижали скорость и маневренность плавания при прикреплении к ним дополнительного груза, достигавшего у4 массы их тела, а подвижность костистых рыб значительно снижается уже при небольшой добавочной нагрузке. Акулы-рейдеры, постоянно находящиеся в движении, имеют температуру тела на 7–8 °С, а акулы Isurus на 10% С выше окружающей среды, что связано с высокими скоростями их передвижений.
Органы пищеварения и питания. Захват и частичная механическая обработка пищи проводится челюстями, снабженными дифференцированной и мощной жевательной мускулатурой и зубами. У хищников, питающихся преимущественно подвижной и крупной добычей, зубы крупные, многовершинные, с режущими краями. Такое строение зубов позволяет удержать сильную добычу или мгновенно откусить от жертвы крупный кусок. У видов, питающихся преимущественно моллюсками и ракообразными, уплощенные, бугорчатые, сидящие плотными рядами зубы образуют своеобразную «терку», дробящую или перетирающую панцирь или раковину. Цельноголовые раскусывают и дробят раковины моллюсков и панцири ракообразных с помощью зубных пластинок.
На дне ротовой полости находится небольшая складка слизистой оболочки – язык; его поддерживает непарный элемент подъязычной дуги. У всех рыб, в том числе и у хрящевых, язык не имеет собственной мускулатуры, его движения обусловлены движениями подъязычной дуги. Ротовая полость незаметно переходит в глотку, стенки которой пронизаны жаберными щелями. Потере пищи через них препятствуют хрящевые жаберные тычинки. Слизь, выделяемая клетками стенок ротовой полости, не содержит пищеварительных ферментов и лишь облегчает проглатывание пищи. Короткий мускулистый пищевод без заметной границы переходит в объемистый желудок, имеющий V-образную форму.
В первой, более объемистой кардиальной части желудка начинается химическое переваривание пищи под воздействием пепсина. Переваривание идет относительно медленно: крупной добычи до 5 и более суток. В значительной степени измельченная и переваренная пищевая кашица переходит в направленную вперед более узкую пилорическую часть желудка, где обрабатывается трипсином, забрасываемым из начальной части кишечника, куда выделяется секрет поджелудочной железы. Кислотность содержимого пилорической части желудка резко снижается. За желудком начинается кишечник, отчетливо разделяющийся на три отдела. Очень короткая тонкая кишка отделена от пилорической части желудка сфинктором – кольцевым утолщением мускульной оболочки. В тонкую кишку открываются протоки поджелудочной железы и желчный проток. Внутри широкой и длинной толстой кишки расположен вырост ее стенки – спиральный клапан, образующий 12–13 витков. Через сфинктер в кишечник протекает только разжиженная пищевая кашица. В тонкой кишке она смешивается с желчью и пищеварительными соками поджелудочной железы. В толстой кишке в щелочной среде завершается переваривание пищи и ее всасывание. Этот процесс облегчается развитием спирального клапана, многократно увеличивающего поверхность кишечника. По короткой прямой кишке непереваренные остатки пищи попадают в клоаку и через клоакальную щель выбрасываются наружу.
От спинной поверхности прямой кишки отходит ректальная железа – полый пальцеобразный вырост – орган солевого обмена. Он Удаляет избыток солей, попадающих в организм с пищей и морской
водой: его железистые клетки выделяют в просвет железы секрет, в котором содержание NaCl примерно вдвое больше, чем в плазме крови. Ректальная железа акул, пойманных в пресных водах, обычно сильно регрессирует: у морских обитателей она весит 3,8–15,9 г, а в пресной воде – только 0,35–1,5 г. По требующим подтверждения данным, у половозрелых особей в период размножения ректальная железа выделяет пахучую слизь, в виде тонких нитей остающуюся в воде позади плывущей рыбы, что помогает встрече особей разного пола.
Трехлопастная печень хрящевых рыб отличается очень крупными размерами: у некоторых акул от 14–20 до 25% массы тела. Печень акуловых рыб накапливает запасы жира, что определяет ее роль не только как хранилища энергических резервов, но и как гидростатического органа, повышающего плавучесть тела. В печени резервируются и некоторые витамины.
Потребность хрящевых рыб в кормах, видимо, относительно невелика. Содержавшаяся в океанариуме австралийская песчаная акула – Carcharias avenarius длиной около 3 м и массой в 150 кг за год съедала всего 80–90 кг рыбы, причем ее «аппетит» резко изменялся по сезонам: наивысший в феврале – апреле и очень низкий в остальное время года. Хрящевые рыбы, особенно акулы, способны за короткое время поглотить большие количества пищи, а затем длительное время голодать, медленно расходуя накопленные ресурсы. Описанные случаи необычайной прожорливости акул, возможно, относятся к таким длительное время голодавшим особям.
Органы дыхания и газообмен. Стенки глотки пронизаны открывающимися наружу жаберными щелями. К подъязычной дуге и хрящевым жаберным дугам прикрепляются межжаберные перегородки, края которых образуют кожистую крышечку над следующей жаберной щелью. На боковых сторонах межжаберных перегородок расположены продольные складки слизистой оболочки – жаберные лепестки, имеющие эктодермальное происхождение. Совокупность жаберных лепестков на одной стороне перегородки образует полужабру. Две полужабры, расположенные по обеим сторонам одной перегородки, составляют жабру. У подавляющего большинства пластиножаберных рыб пять жаберных щелей и 4,5 жабры: одна полужабра на задней стороне перегородки, отходящей от подъязычной дуги, и четыре полных жабры на I–IV жаберных дугах, V жаберная дуга жабр не несет. У пластиножаберных сохраняется остаток жаберной щели между челюстной и подъязычной дугами в виде округлого отверстия, лежащего позади глаза – брызгальца; оно ведет в начальную часть глотки. На его передней стенке есть кожистая складка – клапан, прикрывающий наружное отверстие. По нижнему краю этой складки прикреплена небольшая жабра. В основании каждой межжаберной перегородки проходит приносящая жаберная артерия, распадающаяся на капилляры в жаберных лепестках обеих полужабр; капилляры соединяются в парные выносящие жаберные артерии, над жаберной щелью сливающиеся в общую выносящую артерию, по ним течет уже артериальная кровь.
У акул при вдохе глоточная область расширяется и в нее через ротовое отверстие и через брызгальце насасывается вода, омывающая, жаберные лепестки и проходящая в наружные жаберные полости; при этом давление воды снаружи прижимает клапаны – подвижные края межжаберных перепонок – и они закрывают наружные жаберные щели. При выдохе жаберные дуги правой и левой сторон сближаются, уменьшая объем глотки; жаберные лепестки соседних полужабр почти смыкаются, затрудняя отток воды назад в глотку. Возросшее давление в наружных жаберных полостях отгибает клапаны, и вода вытекает наружу. У лежащих на дне скатов наружные жаберные щели переместились на брюшную сторону. Акт дыхания у них осуществляется сходно с акулами, но вода поступает через брызгальца. Ложная жабра выполняет роль добавочной жабры при поглощении кислорода, но, видимо, более важна ее роль при выделении углекислоты. В отличие от костных рыб функция жабр хрящевых рыб ограничена газообменом; их жаберные лепестки не могут выделять продукты азотистого обмена и соли.
У цельноголовых в отличие от пластиножаберных межжаберные перегородки укорачиваются, а разросшаяся перегородка, прикрепленная к подъязычной дуге, образует кожистую общую жаберную крышку, прикрывающую наружные щели; брызгальца исчезают. Механизм дыхания сходен с дыханием костных рыб.
Кровеносная система и кровообращение. Общая схема кровеносной системы хрящевых рыб схожа с кровеносной системой круглоро-тых, но отличается рядом особенностей, связанных с большей подвижностью хрящевых рыб и с их более высоким уровнем обмена. Но так же, как и у круглоротых, v у хрящевых и костных рыб один круг кровообращения и в сердце только венозная кровь.
Сердце двухкамерное, но состоит из четырех отделов: тонкостенного венозного синуса или венозной пазухи, предсердия, желудочка и артериального конуса. Внешне артериальный конус кажется расширенным началом брюшной аорты, но, как и в остальных отделах сердца, его стенки образованы поперечнополосатой мускулатурой; в стенках кровеносных сосудов, включая и брюшную аорту, – гладкие мышечные волокна. На внутренней поверхности артериального конуса есть клапаны, препятствующие обратному току крови. Последовательное сокращение мускулистых желудочка и артериального конуса повышает кровяное давление, ускоряя кровоток. Такое «поэтапное» увеличение давления, видимо, не особенно эффективно. Кровяное давление в брюшной аорте у хрящевых рыб колеблется в пределах 7–45 мм рт. ст., а у костных рыб 18–120 мм рт. ст.;
при этом у костистых рыб возросла мощность стенок желудочка, тогда как артериальный конус редуцировался.
От артериального конуса отходит брюшная аорта, распадающаяся на пять пар приносящих жаберных артерий. Первая снабжает кровью полужабры подъязычной дуги, вторая – пятая – жабры I–IV жаберных дуг. Окислившаяся в капиллярах жаберных лепестков артериальная кровь собирается в выносящие жаберные артерии. От первой выносящей жаберной артерии ответвляется общая сонная артерия, снабжающая кровью голову. Выносящие жаберные артерии обеих сторон сливаются, образуя лежащую под позвоночником спинную аорту. На уровне плечевого пояса от спинной аорты отходят парные подключичные артерии, несущие кровь к жаберному аппарату и грудным плавникам. В туловищной области от спинной аорты отходит ряд непарных и парных артерий, снабжающих кровью внутренние органы, стенки тела и брюшные плавники. Далее спинная аорта уходит в гемальный канал хвостовых позвонков, получая название хвостовой артерии.
Венозная кровь возвращается в сердце по венам – обычно более широким и тонкостенным, чем артерии. Хвостовая вена, собравшая кровь из хвоста, входит в полость тела и разделяется на правую и левую воротные вены почек; они подходят к почкам и распадаются в них на капилляры, т.е. образуют воротную систему почек. Почечные капилляры вновь сливаются в вены, образующие правую и левую задние кардинальные вены – выносящие сосуды воротной системы почек. По боковым стенкам полости тела идут парные боковые вены, собирающие кровь от брюшных плавников и стенок тела; уже в области сердца каждая из них сливается с подключичной веной, несущей кровь от грудных плавников. Из головы венозная кровь собирается в парные передние кардинальные вены и парные нижние яремные вены. Задние и передние кардинальные и нижние яремные вены каждой стороны сливаются в кювьеров проток. Кювьеровы протоки правой и левой сторон впадают в венозную пазуху сердца.
От пищеварительного тракта и селезенки венозная кровь собирается в несколько вен, перед печенью обычно сливающимися в воротную вену печени. Войдя в печень, она распадается на капилляры; кровь как бы «фильтруется» через ткани печени и вновь собирается в печеночную вену, впадающую в венозный синус.
У хрящевых рыб появляется селезенка – крупный компактный орган, лежащий около желудка. Она выполняет функцию депо крови и служит кроветворным органом: в ней, как и в почках, идет образование форменных элементов крови: эритроцитов, лейкоцитов, тромбоцитов.
Таблица 5. Некоторые показатели крови в разных группах рыб
| Группы рыб | Количество кровн | Содержание гемоглобина | Кислородная емкость крови |
| Скаты Акулы Костные рыбы | 1,5–7,2 3,7–6,8 1,1–7,3 | 0,8–4,5 3,4–6,5 1,1–17,4 | 1,1–6,0 4,5–8,7 1,5–23,0 |
Количество крови в организме, ее кислородная емкость и содержание гемоглобина у разных видов рыб варьирует в широких пределах, в зависимости от их подвижности. У менее подвижных скатов эти показатели несколько ниже, чем у акул. У наиболее активных и подвижных костных рыб содержание гемоглобина и кислородная емкость крови в несколько раз выше максимальных показателей хрящевых рыб. Это свидетельствует в среднем о более высокой энергетике организма костных рыб.
Органы выделения и водно-солевой обмен. Основные органы выделения хрящевых рыб – туловищные, или мезонефрические, почки, в виде удлиненных тел лежащие по бокам позвоночника, вдоль почти всей полости тела. Передние их части сильно сужены; задние, собственно выполняющие функцию почки, расширены. Основную массу почки заполняют неф-роны, состоящие из гломерул; от последних отходят почечные канальцы. У некоторых хрящевых рыб наряду с гломерулами сохраняются и нефростомы – мерцательные воронки, открывающиеся в полость тела на поверхности почки. Стенки почечных канальцев, особенно их извитых частей, оплетены кровеносными капиллярами сосуда, вышедшего из боуме-новой капсулы, и капиллярами воротных вен почек. В стенках некоторых отделов почечных канальцев имеются железистые клетки, секретирующие в просвет канальцев продукты азотистого распада. Почечные канальцы впадают либо в вольфов канал, выполняющий функцию мочеточника, либо в обособляющийся от нижней части вольфова канала мочеточник. Почки получают артериальную кровь по почечным артериям; большие порции венозной крови приходят по воротным венам почек.
Продукты распада лишь в малой доле попадают из целомической жидкости в почечные канальцы через сохранившиеся у некоторых видов реснитчатые нефростомы. Основной путь – это фильтрация кровяной плазмы через стенки капилляров мальпигиевых клубочков. Такая первичная моча, поступившая из гломерул в почечные канальцы, содержит не только большое количество воды, но и такие ценные для организма низмолекулярные вещества, как сахара, витамины, аминокислоты и др. Они вновь всасываются через стенки канальцев. Подобный тип фильтрующей почки возник еще у живших в пресных водах Древних позвоночных: он позволял удалять избыток проникавшей в тело через покровы воды, но угрожал потерей и необходимых организму веществ. Образование сложной структуры почечных канальцев с железистым аппаратом в их стенках разрешило и эту проблему. Однако при переходе в море обладавшие такой пресноводной почкой примитивные хрящевые рыбы столкнулись с другой опасностью – обезвоживанием тела вследствие отдачи воды через покровы во внешнюю среду, соленость которой в 2–3 раза превышала содержание солей в крови и тканевых жидкостях. Эволюция хрящевых рыб шла своеобразным биологически удачным путем, сохранив фильтрационный аппарат почек и биохимически обеспечив почти полную изотоничность крови и тканевых жидкостей внешней среде. Это было достигнуто удержанием в крови и тканевых жидкостях мочевины и солей.
Таблица 6. Величины депрессии внешней среды, крови и мочи у различных групп рыб
| Экологические группы | Д внешней среды | Д крови | Содержание мочевины в крови, % | Д мочи |
| Хрящевые рыбы: морские пресноводные Костистые рыбы: морские пресноводные | 1,80–2,30 0,02–0,03 1,80–2,30 0,02–0,03 | 1,90–2,30 0,90–1,20 0,67–0,91 0,42–0,70 | 1,25–2,60 0,10 0,01–0,03 0,005–0,03 | 1,87–2,3 0,49–0,78 0,09–0,90 0,07–0,08 |
За счет повышения содержания солей у морских хрящевых рыб обеспечивается примерно 50% осмотического давления крови и тканей большее увеличение содержания солей для позвоночных, видимо, физиологически невозможно или невыгодно. Дальнейшее повышение осмотического давления крови хрящевыми рыбами было достигнуто удержанием в кровяном русле и в тканях большого количества мочевины – довольно токсичного вещества – и физиологически менее вредного триметиламиноксида. Это позволило повысить общее осмотическое давление крови и тканей и практически уравнять его с осмотическим давлением окружающей среды.
Условием удержания в организме мочевины и других близких соединений была выработка непроницаемости для них жаберных лепестков; у костистых рыб последние легко отдают мочевину во внешнюю среду. Постоянно высокая уремия хрящевых рыб должна была сопровождаться приобретением особых буферных приспособлений. Хотя эти биохимические и физиологические механизмы пока достаточно не изучены, можно предполагать, что они имеют отношение к тому своеобразному химизму тела хрящевых рыб, в частности высокому содержанию солей тяжелых металлов, о котором говорилось выше.
Накопление и удержание мочевины обеспечивается рядом особенностей. В почках через мальпигиевы клубочки идет энергичная фильтрация. Из первичной мочи, перемещающейся по просвету канальца, через его стенки абсорбируются сахара, аминокислоты, некоторые соли и мочевина. Это облегчается не только большей относительной длиной почечных канальцев хрящевых рыб по сравнению с другими рыбами, но и обособлением особого сегмента почечного канальца, отсутствующего у других рыб, в стенках которого, видимо, и происходит в основном обратная абсорбция мочевины. Лишь излишки мочевины, солей и воды выделяются с мочой наружу. Нарушению ионного равновесия препятствует деятельность ректальной железы, способной, активно концентрируя соли, выводить их избыток во внешнюю среду.
Морские хрящевые рыбы, достигнув этим путем изотонии со средой, выделяют небольшое количество мочи: всего от 2 до 50 мл на 1 кг массы тела в сутки. Хрящевые рыбы, живущие в опресненных или пресных водоемах, снизили осмотическое давление в крови и тканях и содержание мочевины, но остаются гипертоничными к среде. Через их покровы проникает вода, грозя организму излишним обводнением, что усиливает водо-выделительную функцию их почек: количество выделяемой мочи у пилы-рыбы достигает 250 мл на 1 кг массы тела за сутки; одновременно соли активно адсорбируются в почечных канальцах. Такой тип водно-солевого обмена и ионного равновесия со средой требует минимальных затрат энергии.
Половая система и особенности размножения. Парные семенники у самцов подвешены на брыжейках по бокам пищевода над печенью. Протоки семенных канальцев семенников – тонкие семя-выносящие канальцы – лежат в брыжейке н впадают в почечные канальцы передней узкой части почки. Эта часть почки не функционирует как орган выделения и превращается в придаток семенника; его канальцы открываются в вольфов канал. Он функционирует как семяпровод. В самой задней части семяпровода у половозрелых самцов образуется расширение – семенной пузырек. Семяпроводы правой и левой стороны открываются в полость мочеполового сосочка. Рядом с ними туда же открываются отверстия тонкостенных полых выростов – семенных мешков; это остатки мюллеровых каналов. В полость мочеполового сосочка впадают и мочеточники. Мочеполовой сосочек отверстием на своей вершине открывается в полость клоаки. Формирование мужских половых клеток происходит в канальцах семенника. Еще не дозревшие сперматозоиды по семевыносящим канальцам попадают в придаток семенника–переднюю часть почки – ив его канальцах дозревают. Зрелые сперматозоиды проходят по семяпроводу и скапливаются в семенных пузырьках и семенных мешках. При оплодотворении сокращением стенок семенных пузырьков и семенных мешков сперматозоиды выбрасываются в клоаку самца, а затем с помощью копулятивных органов вводятся в клоаку самки. Внутреннее оплодотворение – характерная особенность всех хрящевых рыб.
Самки имеют парные яичники, подвешенные на брыжейках в тех же местах, где у самцов находятся семенники. У неполовозрелых самок яичники по внешнему виду похожи на семенники. Вольфов канал у самок выполняет только функцию мочеточника. Мюллеровы каналы хорошо развиты; каждый из них лежит на брюшной поверхности соответствующей почки. У большинства хрящевых рыб передние концы мюллеровых каналов – они у самок выполняют функцию яйцеводов–огибают передний конец печени и, сливаясь, образуют общую воронку яйцевода; она лежит у брюшной поверхности центральной доли печени и у половозрелых самок имеет широкие бахромчатые края. У немногих видов каждый яйцевод заканчивается воронкой самостоятельно. В области передней части почек каждый яйцевод образует расширение – скорлуповую железу; заметно расширенная задняя часть яйцевода называется «маткой». Яйцеводы правой и левой сторон открываются в клоаку самостоятельными отверстиями по бокам мочевого сосочка.
Непосредственной связи между яичником и яйцеводом нет. Созревшее яйцо через разрыв стенки фолликула выпадает в полость тела, скатывается по печени и попадает в резко увеличивающуюся к этом) моменту воронку яйцевода. Благодаря перистальтическим сокращениям стенок яйцевода яйцо постепенно перемещается по яйцеводу, оплодотворяется, затем в области скорлуповых желез покрываете студенистой белковой оболочкой и поверх нее – почти у всех яйце кладущих видов – плотной рогоподобной оболочкой, часто имеющее выросты и жгуты. Такая оболочка защищает яйцо и развивающегося эмбриона от обезвоживания в морской воде, от многи) хищников и механических повреждений. При помощи выростов i нитей яйца подвешиваются на водорослях, цепляются за неровности подводных скал. Яйца крупные, содержат много желтка. У разнозубых акул длиной около 1,5 м длина яйцевой капсулы примерно 10 см ромбообразные скаты длиной 1 м откладывают яйца 10-х 6 см; удлиненные яйца химер достигают 20 см. Плодовитость хрящевых рыб невелика. Одновременно обычно откладывается 1–2, редко 10–12 яиц через некоторое время откладка повторяется. Лишь полярная акула достигающая 5–8 м, откладывает до 500 яиц длиной около 8 см.
Большие запасы желтка в яйце хрящевых рыб обусловливаю отсутствие полного дробления яйца: делится лишь сравнительно белая желтком часть на анимальном полюсе. Образуется плавающий на желтке зародышевый диск, задний край которого претерпевает гаструляцию и потом здесь же обособляется зародыш. Остальные части зародышевого диска обрастают желток, образуя желточный мешок. Сформировавшийся зародыш связан с желточным мешком пупочным канатиком, по которому проходят кровеносные сосуды. Е мере ассимиляции желтка зародышем размеры желточного мешка уменьшаются, а его остатки перед вылуплением втягиваются в брюшную полость зародыша. Развитие плотной, оболочки, затрудняющее доступ кислорода в яйцо, привело к появлению у зародышей многих видов наружных жабр; перед вылуплением они рассасываются.
Эмбриональное развитие идет медленно. У разнозубых акул оно продолжается до 7, у кошачьих акул – до 8–9, у ромбовидных скатов – от 4 до 14 и у химер – до 9–12 месяцев. Вылупляющийся из яйца детеныш отличается от взрослой рыбы только размерами и вполне способен к самостоятельной жизни.
Для многих акул и скатов характерно яйцеживорождение: оплодотворенные яйца задерживаются в маточных частях яйцеводов. Особых связей между зародышем и материнским организмом не возникает: развитие идет за счет энергетических запасов яйца, но кислород зародыш, видимо, получает путем осмоса и диффузии из материнского организма. Рождаются вполне зрелые, способные к самостоятельной жизни детеныши. У морской лисицы Alopias vulpes развивается 2– 4 детеныша, при рождении достигающие 1–1,5 м длины, самка катрана – Squalus acanthias через 6–7 месяцев беременности приносит от 12 до 30 акулят длиной 20–26 см, а тигровая акула – Galecerdo cuvieri – до 30–80 детенышей длиной 45–50 см. У сельдевых акул p. Lamna зародыши, использовав запас своего желточного мешка, поедают скопившиеся в матке неоплодотворенные яйца; самка рождает 3–5 детенышей длиной до 75 см.
Наконец, у некоторых акул и скатов можно говорить о настоящем живорождении, когда между зародышем и материнским организмом возникают тесные связи. У куньих акул p. Mustelus одновременно развивается до 20, а у молот-рыбы Sphyrna – даже до 30–40 зародышей. Образующийся вокруг желтка и снабженный кровеносными сосудами зародыша желточный мешок после использования желтка прирастает к стенке матки; возникает своеобразная плацента: кровеносные сосуды зародыша и матери лежат близко друг от друга; путем осмоса и диффузии кислород и питательные вещества из кровотока матери попадают в кровоток зародыша, а продукты распада – в кровь матери. Интересно заметить, что подобную «плаценту» акул впервые описал еще Аристотель. У некоторых скатов на стенках матки образуются пальцеобразные выросты, врастающие в брызгальца зародыша; железистые клетки этих выростов выделяют в глотку зародыша питательную жидкость, которую он заглатывает. Одновременно скаты вынашивают до 6–12 детенышей, но гигантская манта – только одного.
Внутреннее оплодотворение, крупные размеры яиц, содержащих значительные запасы питательных веществ, прочная яйцевая наружная оболочка, наконец, широкое распространение яйцеживорождения и даже живорождения – приспособления, резко снизившие эмбриональную и постэмбриональную смертность. Это обеспечивает существование при в общем невысокой – по сравнению с другими водными животными – плодовитости. Такая «экономичность» размножения, несомненно, способствовала сохранению хрящевых рыб и помогла им занять заметное место в сложных, насыщенных жизнью современных биоценозах Мирового океана.
Центральная нервная система и органы чувств. По сравнению с круглоротыми у хрящевых рыб не только возрастают относительные размеры головного и спинного мозга, но усиливается дифференцировка отделов, усложняются связи между ними.
Возрастают относительные размеры переднего мозга. Проходящая сверху продольная борозда внешне как бы делит его на полушария, но внутри этого подразделения еще нет. Очень велики обонятельные доли. Нервные клетки образуют скопления не только в обонятельных долях и в полосатых телах на дне переднего мозга, но сплошным слоем выстилают полость желудочков. Функционально передний мозг представляет высший центр переработки информации, получаемой от органов обоняния. Связи переднею мозга со средним и промежуточным мозгом обеспечивают его участие в регуляции движения и поведения.
Массивный промежуточный мозг имеет хорошо развитые зрительные бугры. На его спинной стороне есть эпифиз, а на брюшной – гипофиз. Образуется хиазма зрительных нервов. Промежуточный мозг служит первичным зрительным центром, участвует в переработке информации от других органов чувств, играет роль в координации движений, а через гормональную деятельность связанных с ним желез участвует в регуляции метаболизма и его сезонных перестройках.
Средний мозг развит хорошо, но его относительные размеры значительно меньше по сравнению с костными рыбами. Сверху он подразделен на две зрительные доли, в которых заканчиваются проводящие тракты зрительного анализатора. Возникают связи среднего мозга с мозжечком, продолговатым и спинным мозгом. Мозжечок велик и прикрывает часть среднего и продолговатого мозга. У акул по сравнению со скатами не только возрастают относительные размеры мозжечка, но на его поверхности образуется сложная система извилин. Мозжечок поддерживает мышечный тонус, равновесие и общую координацию движений; в нем замыкаются рефлексы, связанные с рецепторами органов боковой линии. Продолговатый мозг удлинен и имеет четко выраженную ромбовидную ямку – объемистую полость четвертого желудочка. Это центр регуляции рефлекторной деятельности спинного мозга и вегетативной нервной системы, координирующий работу скелетно-мышечной, кровеносной, дыхательной, пищеварительной и выделительной систем. Здесь расположены ядра вестибулярного аппарата и органов боковой линии, а связи с другими отделами головного мозга позволяют ядрам продолговатого мозга принимать участие в выработке ответа организма на сигналы и других органов чувств. У рыб, имеющих электрические органы, их разряды регулируются деятельностью специальных электрических ядер продолговатого мозга.
От головного мозга хрящевых рыб отходят 10 пар хорошо развитых и симметрично расположенных головных нервов; XII пара головных нервов развита слабо.
Продолговатый мозг незаметно переходит в спинной мозг, который у хрящевых рыб имеет вид округлого тяжа, лежащего в канале, образованном верхними дугами и верхними вставочными пластинками. По сравнению с круглоротыми у хрящевых рыб возрастает число нервных клеток спинного мозга и отчетливо выражены брюшные рога серого вещества, тогда как спинные лишь слабо намечены. Развиваются и усложняются восходящие и нисходящие проводящие пути; проводящие пути боковых стволов достигают продолговатого мозга и мозжечка. Это приводит к большей координации головным мозгом деятельности спинного мозга. Несмотря на это спинной мозг хрящевых рыб сохраняет еще значительную автономность: обезглавленные акулы под действием сильных стимуляторов могут совершать ненаправленные движения. У так называемых электрических рыб в спинном мозге имеются особые нервные клетки, волокна которых идут к электрическим органам, обеспечивая координацию спинным мозгом работы электрических органов; в свою очередь она контролируется электрическими центрами продолговатого мозга. Это обеспечивает синхронность разрядов отдельных элементов электрического органа, резко повышая мощность общего разряда.
Симпатическая нервная система хрящевых рыб, как и у кругло-ротых, представлена цепочкой симпатических ганглиев, между которыми еще нет прямой продольной связи; связь между ганглиями осуществляется пока через спинной мозг и через соединения нервов отдельных ганглиев в сплетениях на внутренних органах.
Органы чувств хрящевых рыб много сложнее и совершеннее органов чувств круглоротых.
Один из основных органов рецепции – обоняние. Объемистые обонятельные мешки парные; каждый из них лежит в хрящевой обонятельной капсуле и сообщается с внешней средой через ноздрю. Внутренняя поверхность мешков несет многочисленные складки, покрытые обонятельным эпителием, связанным с окончаниями обонятельного нерва. Парные ноздри открываются наружу на нижней стороне рыла впереди ротовой щели. У большинства видов от каждой ноздри к краю ротовой щели идет довольно глубокая назооральная борозда, прикрытая складкой кожи. По этой борозде вода от ротовой щели проходит к ноздре. Поэтому рыба может ощущать запах схваченной добычи. Эксперименты показывают высокую чувствительность органов обоняния. Акулы в некоторых случаях ощущают запах добычи за 400–500 м, обнаруживают мясо, положенное в плотный мешок. Вероятно, при помощи органов обоняния одиночно живущие особи разного пола находят друг друга в сезон размножения, улавливая выделения ректальной железы. Ямковидный орган, расположенный в ротовой полости, как показали недавние наблюдения, служит детектором солености.
Очень важны для ориентации и кожные сейсмосенсорные органы. У хрящевых рыб они представлены двумя типами образований. По боковой поверхности тела в толще кожи проходит канал боковой линии; на голове и у некоторых, особенно придонных видов, на брюхе канал сильно ветвится. Многочисленные мелкие отверстия сообщают полость канала с внешней средой. На его стенках расположены скопления чувствующих клеток: они имеют реснички, а основание каждой клетки оплетено окончанием боковой ветви блуждающего нерва. Реснички чувствующих клеток воспринимают даже слабые токи воды. У примитивных акул и у химер настоящего канала нет. Орган боковой линии у них имеет вид узкой и глубокой кожной борозды, также местами ветвящейся. Органы боковой линии воспринимают механические перемещения частиц воды и, возможно, инфразвуковые колебания. Они играют важную роль при видовом общении, при поисках пищи и избегании опасности и при ближней ориентации. В последнем случае боковая линия дает возможность «эхолокации» – улавливания отраженных от дна и подводных предметов волн, вызванных движением самой рыбы.
На голове имеются скопления лоренциниевых ампул. Каждая из них представляет погруженную в толщу кожи маленькую соединительнотканную капсулу, от которой отходит тонкая трубочка, открывающаяся отверстием на поверхности кожи. На стенках капсулы расположены скопления чувствующих клеток, а полость капсулы и трубочки заполнены слизью. Высказываются предположения, что лоренщшиевые ампулы выполняют роль термодатчиков, улавливающих перепады температуры внешней среды до 0,05° С. В то же время от воспринимают электрические поля напряжением до 0,1–0,01 mB/civ и позволяют находить неподвижную, но живую добычу Эти предположения требуют доказательств.
При характеристике отряда электрических скатов слабые разряды, и используют его для ориентации. Слабо развитые электрические органы обнаружены и у других скатов; они генерируют слабы' электрические поля. Разряды напряжением 0,2–2 В следуют друг з; другом с частотой 20–300 импульсов в 1 с. Улавливая изменение электрического поля, рыба, видимо, может определить размеры и положение посторонних предметов, попадающих в поле. Предполагают что рецепторными органами, улавливающими изменения электрического поля, служат не только лоренциевы ампулы, но и рецептор канала боковой линии.
Электрические органы скатов представляют собой сильно измененные участки поперечнополосатой мускулатуры. Особенно они велики у электрических скатов, составляя до четверти вес рыбы. Мышечные волокна преобразуются в электрические пластинки отделенные друг от друга студенистой соединительной тканью. К каждой пластинке снизу подходит и сильно ветвится веточка нерва, верхняя часть пластинки заполнена разветвлениями кровеносных сосудов. Нижняя часть пластинки электрически отрицательна, верхняя – положительна. Скопления 35–40 рядов электрических пластинок образуют призмы, разделенные друг от друг стенками волокнистой соединительной ткани. Каждый орган состоит из 350–600 призм, содержащих в совокупности до 100–200 тыс. электрических пластинок. К органу подходят мощные ветви головных нервов. Электрический орган действует по принципу Лейденской банки, накапливая электричество, образующееся в процессе обмена в электрических пластинках. Мощные разряды при защите и нападении, достигающие у скатов напряжения в 60–300 В при силе тока до 5 А, происходят под воздействием импульсов из электрических ядер продолговатого мозга. По некоторым данным скаты могут генерировать с интервалами в доли секунды до 100 разрядов без заметного снижения их мощности.
Глаза хрящевых рыб крупные, с уплощенной роговицей и круглым хрусталиком, имеют типичное для всех рыб строение. Около глаза кожа образует невысокий валик – неподвижное кольцевидное веко. У некоторых акул есть подвижная мигательная перепонка. У акул и химер глаза подвижны, у скатов часто склера прирастает к глазнице. Глаза используются как рецепторы близкой ориентации: контуры предметов и их движение крупные виды различают, видимо, не далее 10–15 м; у мелких видов дальность видения меньше. Зрение у хрящевых рыб черно-белое; цветов они не различают.
Орган слуха и равновесия хрящевых рыб представлен только внутренним ухом, заключенным в хрящевую капсулу. Парные – по одной с каждой стороны – слуховые капсулы врастают в заднебоковую стенку мозгового черепа. Внутреннее ухо – собственно перепончатый лабиринт – включает круглый и овальный мешочки и соединенные с последним хорошо развитые три полукружных канала, лежащие во взаимно перпендикулярных плоскостях. Полукружные каналы выполняют функцию органа равновесия; круглый и отчасти овальный мешочки – органы слуха. Принципы работы органа слуха и равновесия уже были описаны. Слух хрящевых рыб изучен недостаточно. Они воспринимают преимущественно низкие звуки. Слух, несомненно, играет важную роль в их общении и ориентации. Улавливают так называемые «механические» звуки, возникающие при движении, захватывании и разгрызании добычи. Некоторые виды способны издавать и специальные сигнальные звуки, обеспечивающие защиту индивидуального участка, взаимное общение партнеров по стае и облегчающие встречу при размножении.
Вкусовые почки обнаружены в слизистой оболочке ротовой полости и глотки. Осязательные тельца на участках тела с сильно развитой плакоидной чешуей, видимо, отсутствуют или развиты слабо; более обильны они на участках голой кожи. Усики на рыле пилоносных акул, вероятно, выполняют осязательную функцию и, возможно, несут и вкусовые почки.
Поведение, образ жизни и распространение хрящевых рыб
В эволюции усложнение подвижности и возрастание скорости движений вырабатывалось одновременно с совершенствованием рецепторов – органов чувств – и анализаторов центральной нервной системы. Обогащалось восприятие внешнего мира, усложнялась нервная деятельность. Особенности поведения и характер нервной деятельности хрящевых рыб пока изучены слабо. Можно говорить о том, что в поведении преобладают безусловно рефлекторные действия. Они принимают форму сложных инстинктов, обеспечивающих поиск и добывание пищи, размещение в наиболее благоприятных местах, миграции, размножение. В какой степени играет в жизни хрящевых рыб индивидуальный опыт, пока сказать трудно. Видимо, выработкой индивидуального опыта в некоторых случаях можно объяснить появление акул-людоедов – особей, «предпочитающих» нападать на человека.
Несомненно существует, но пока мало выяснена внутривидовая организация, различающаяся у разных видов. Гигантские акулы – мирные планктоноеды – «пасутся» на богатых планктоном участках океана группами до 20–30 особей. Почти столь же большими группами их обнаруживали и на дне в неактивный период. С другой стороны, сходная по питанию китовая акула, видимо, встречается только поодиночке. Более подвижные хищники – рейдеры чаще держатся группами по 2–3 особи, но при появлении крупной добычи быстро собираются в стаи, которые вскоре вновь распадаются. Высказывалось предположение, что рогатые скаты, особенно гигантская манта, держатся семейными группами. Придоннопелагические мелкие акулы обычно охотятся и кочуют стаями, размеры которых меняются в широких пределах.
Для донных акул, для большинства скатов и химер характерен малоподвижный, более или менее оседлый образ жизни. Скаты большую часть времени проводят полузакопавшись в грунт. Животные, видимо, имеют свои индивидуальные участки, которые иногда охраняют от вторжения особей своего вида. Размеры участков зависят от размеров рыбы, ее пищевой специализации и численности пищевых объектов, характера грунта и т.п. В некоторых случаях рыбы живут всего в нескольких метрах друг от друга, в других – на большом расстоянии.
Миграции хрящевых рыб изучены слабо. Они, прежде всего, связаны с сезонным изменением температуры воды и определяемым им перераспределением кормовых объектов. У части видов в разной степени выражены и миграции, связанные с размножением. Многие виды весной подходят к берегам и на мелководья, где в это время, видимо, обильнее пища; в этих же районах идет спаривание и деторождение. Скаты и химеры зимой держатся на более глубоких участках, чем летом. Но полярная акула, наоборот, зимой приближается к берегам и заходит в морские заливы, а летом держится в открытом море. Во время миграций у части видов резко возрастает величина стай, иногда достигающих колоссальных размеров. Так, за один замет невода у берегов Англии было поймано сразу около 20 тыс. катранов, а в Азовском море и Керченском проливе ловили сразу до 10 тыс. скатов – морских котов Dasyatis pectinata.
Географическое распространение. Многие хрящевые рыбы приурочены к теплым экваториальным водам Мирового океана. Например, в водах у Южной Японии встречаются представители примерно половины ныне живущих родов хрящевых рыб. Немногие виды – обитатели умеренных и холодных вод. Скат Raja hyperborea и полярная акула встречаются в водах, имеющих минусовую температуру. Некоторые виды распространены очень широко: от полярных вод до тропиков; такие виды с почти космополитическим распространением в тропиках обычно держатся на больших глубинах, где вода холоднее. Встречаются и виды с ограниченными ареалами.
Экономическое значение хрящевых рыб
Большинство видов хрящевых рыб не образует плотных скоплений, что затрудняет их добычу. Поэтому объектами специального промысла служат лишь немногие виды и в ограниченных районах. Значительно чаще многие виды хрящевых рыб добываются попутно. По весу хрящевые рыбы составляют около 1,5–2% мировой добычи рыбы. Использование хрящевых рыб очень различно в разных странах; в Австралии и Японии многие виды охотно используются в пищу, в Америке и Европе те же или близкие виды перерабатываются преимущественно на кормовую муку или экспортируются в те страны, где на них есть спрос. Довольно часто свежее или копченое мясо акул продается под видом других рыб.
Из печени акул добывают жир. Он содержит большое количество витамина А и используется в пищу и в медицинских целях как рыбий жир. Во время второй мировой войны, когда резко упал промысел трески, до 75% вырабатываемого в США витамина А получали из акульего жира. Промысел многих видов акул проводился необычайно интенсивно. Но после разработки технологии получения синтетического витамина А промысел их резко сократился. Из акульего жира вырабатывают специальные смазки для ряда точных приборов, используют его в косметике и др. Находит применение и кожа многих видов акул и скатов. Она используется для шлифовки; после специальной обработки из нее шьют высококачественные галантерейные и обувные изделия: сумки, портфели, туфли.
Для уменьшения случаев нападения акул на людей идет поиск специальных репелентов – веществ, отпугивающих акул. Предложенные пока препараты малоэффективны. В ряде мест – у пляжей Австралии, Калифорнии и в некоторых других районах – ведется усиленный промысел только с целью уменьшения численности крупных акул. Это позволяет обеспечить большую безопасность купающимся. Крупные виды акул местами служат объектами спортивного лова.
Недостаток животных белков в пищевых рационах населения многих стран мира вызывает необходимость более широкого использования в пищу объектов морского промысла, в том числе и хрящевых рыб. При этом следует учитывать, что поздние сроки наступления половой зрелости и относительно невысокая плодовитость хрящевых рыб делает их весьма уязвимыми: при интенсивном промысле их численность восстанавливается много медленнее, чем у многих других промысловых рыб.
Похожие работы

способлениями от избытка взвеси в воде у обитателей мутных вод являются уменьшение глаз (т. е. сокращение наиболее уязвимых участков поверхности тела) и усиленное выделение слизи, которая, осаждая муть, обеспечивает чистоту воды вокруг тела рыбы и таким образом улучшает условия дыхания. Глава I. Строение и некоторые физиологические особенности рыб · Строение и функции организма рыбы ...



... которая при их отсутствии неминуемо опускалась бы книзу при плавании. Короткое коническое рыло сжато с боков, у мелких особей оно имеет вид клюва или короткого хобота . Характерную особенность гигантской акулы составляют очень большие жаберные щели, охватывающие голову от спинной стороны до горла. Каждая жаберная дуга несет по переднему краю 1000—1300 длинных роговых жаберных тычинок, образующих ...
... . Эволюция органического мира значительно ускоряется в эпохи интенсивного тектонического развития земной коры и обновления органического мира. Заключение. В результате работы над рефератом по теме «Живое прошлое Земли» мы познакомились с геологической историей Земли, основными этапами ее развития и ключевыми моментами повлиявшими на появление жизни на Земле и возникновение новых форм фа
... . Различают водоемы: пресные - с минерализацией 1000 мг/л (1 ‰), солоноватые - с соленостью от 1 до 25 ‰, соленые - с соленостью от 25 до 50 ‰ и минеральные (или соляные) с соленостью более 50 ‰. По отношению к солености рыбы делятся на две большие экологические группы: евригалинные — живут при переменной солености, стеногалинные — при постоянной. По приуроченности к характерным экологическим ...
0 комментариев