Расселение организмов по планете - очень широко распространённое в природе явление, которое, вероятно, существует практически с момента их возникновения на планете. В те или иные периоды существования животного и растительного мира расселение шло с различной интенсивностью. В течение ХХ столетия интенсивность такого расселения значительно увеличилась, что связано не столько с природными процессами, сколько с активной природопреобразующей деятельностью человека (Элтон, 1960).
Как отмечает А.Ф. Карпевич (1975), в области акклиматизации водных животных долгое время не существовало сколько-нибудь твёрдых теоретических принципов. Предполагалось, что на современном этапе эволюции гидробионтов процесс их расселения остановился, а комплекс приспособлений гидробионтов адаптирован к определённой среде обитания. К тому же в природе не может иметься свободных экологических ниш, поэтому удачное вселение нового вида водных животных в тот или иной биоценоз чрезвычайно маловероятно. Однако накапливавшийся по мере развития цивилизации опыт показывал, что расселение гидробионтов не только возможно теоретически, но и реально существует в природе.
Отсутствие теоретических обоснований акклиматизации приводило к низкой интенсивности и эффективности этого процесса в случае его искусственного проведения. Первый документально зафиксированный опыт по искусственному переселению рыб в России был проведён со стерлядью (Acipenser ruthenus), вселённой в 1763 г. в р. Неву. В 1857 г. было создано Русское общество по акклиматизации. Однако до 20-х гг. ХХ в. интенсивность акклиматизационных работ была низкой: с 1820 по 1850 гг. зарегистрировано лишь 5 пересадок рыб, с 1990 по 1920 гг. - 200 (Карпевич, 1975; Строганова, Задоенко, 2000).
Число интродукций резко возросло в 1920-1940 гг. За этот период осуществили более 1500 пересадок 40 видов и форм. Однако «коэффициент их полезного действия» был очень низок из-за слабых теоретических знаний закономерностей процесса акклиматизации и ряда других причин (Карпевич, 1975).
Первое теоретическое обоснование необходимости и возможности акклиматизации гидробионтов в бывшем СССР привёл Л.А. Зенкевич (1940), рассмотревший проблемы вселения в Каспийское море кормовых беспозвоночных. В 1940-1980-х гг. наибольший вклад в решение теоретических и практических вопросов акклиматизации гидробионтов внесли А.Ф. Карпевич (1947, 1948, 1960, 1962, 1968, 1975 и др.), Е.В. Бурмакин (1956, 1961), П.А. Дрягин (1953, 1954), Ф.Д. Мордухай-Болтовской (1960), Т.С. Расс (1962, 1965) и ряд других исследователей. В Азово-Черноморском регионе к числу наиболее значимых, на наш взгляд, исследований следует отнести работы С.И. Дорошева (1964), Ю.И. Абаева (1971).
Подробную трактовку терминов, используемых при рассмотрении вопросов, связанных с акклиматизацией, приводит А.Ф. Карпевич (1975, 1998). Согласно её взглядам:
Интродукция - это перенос организмов с целью введения их в новую область, водоём, культуру. Она является первым этапом процесса акклиматизации, но не обязательно интродукция заканчивается акклиматизацией интродуцента.
Акклиматизация - процесс приспособления интродуцированных особей и их потомства к новым условиям среды, а также формирования в них новой популяции вида на основе ограниченного генофонда и под действием естественного отбора, в результате чего в биологии и морфо-физиологическом облике последующих поколений переселенца возникают изменения.
Вселение - процесс переноса особей в тот или иной водоём.
Зарыбление - регулярный выпуск молоди одного и того же вида на нагул в определённые водоёмы.
Натурализация - конечная фаза процесса акклиматизации, когда вселенец приспособился к новым условиям, определилась его ниша и взаимоотношения с аборигенами в экосистеме водоёма, установилось подвижное равновесие численности новой популяции, и выявилась возможность её использования в кормовых или промысловых целях.
Поэтапная акклиматизация - незавершённая акклиматизация, когда некоторые этапы развития вселенца не могут завершиться в условиях заселяемого водоёма и проходят в других водоёмах или при непосредственном влиянии человека.
Реакклиматизация - интродукция особей вида в целях восстановления его утраченного ареала.
Аутоакклиматизация (самоакклиматизация) - самостоятельное вселение водных организмов с последующей их акклиматизацией в новом водоёме. А.Ф. Карпевич (1975) в этом процессе выделила две составляющие - палеоспонтанное расселение и самоакклиматизацию без участия человека и неоспонтанное расселение и акклиматизацию при прямом либо косвенном участии человека.
Безусловно, трактовка указанных терминов достаточно разнообразна.
Например, Л.А. Зенкевич (1940) под акклиматизацией понимал успешное существование и развитие какого-либо живого организма в новом ареале в естественных условиях и различал акклиматизацию в естественной среде и в процессе искусственного культивирования.
В Биологическом энциклопедическом словаре (1989) приводятся следующая трактовка этого понятия: «Акклиматизация - приспособление организмов к новым или изменившимся условиям существования, в которых они проходят все стадии развития и дают жизнестойкое потомство» (с. 14).
По мнению Н.З. Строгановой и И.Н. Задоенко (2000), акклиматизация водных организмов - это «биолого-биотехнический процесс, представляющий собой вселение объектов, доставленных из одних водоёмов, регионов, стран в другие, где их ранее не было или они исчезли, с целью их полной или частичной натурализации, а также других форм хозяйственного использования…» (с.51).
Водоём, из которого перемещается организм, называют водоёмом-донором, а водоём, куда организм перемещается - водоёмом-реципиентом. Направления инвазии новых видов называют векторами и обычно подразделяют на естественные и антропогенные. Первые обычно характеризуются относительно невысокими темпами расселения, вторые – более быстрыми (Алимов и др., 2000; Горелов, 2000).
Процесс акклиматизации вида всегда сопровождается определёнными взаимодействиями с элементами местных биоценозов. В связи с этим Л.А. Зенкевичем (1940) были выделены два типа акклиматизации - акклиматизация внедрения и акклиматизация замещения. Первая форма акклиматизации происходит при существовании в водоёме свободной экологической ниши, занимаемой акклиматизантом, в результате чего он практически не конкурирует с местными видами. При акклиматизации замещения вселенцы вторгаются в экологические ниши местных видов и вступают с аборигенными формами в конкурентные отношения за те или иные факторы среды.
Представления Л.А. Зенкевича (1940) о типах акклиматизации были дополнены А.Ф. Карпевич (1975), выделившей также акклиматизацию отторжения, акклиматизацию пополнения и акклиматизацию конструирования. В первом случае акклиматизант вступает в конкурентные отношения с местными видами, но уступает им в этой борьбе и либо гибнет, либо оказывается очень малочисленным. При акклиматизации пополнения вселенцы пополняют состав обеднённого населения водоёмов. При акклиматизации конструирования переселенцев подбирают для построения пищевых цепей, сообществ или фаун водоёмов.
Численность вида-акклиматизанта в водоёме не остаётся постоянной. На это обратил внимание ещё Л.А. Зенкевич (1940). А.Ф. Карпевич (1975) выделила пять «узловых фаз процесса акклиматизации и натурализации вида в новых условиях» (с.119):
1. Выживание переселённых особей в новых для них условиях - период физиологической адаптации;
2. Размножение особей и начало формирования популяции;
3. Максимальная численность переселенца - фаза «взрыва»;
4. Обострение противоречий переселенца с биотической средой;
5. Натурализация в новых условиях.
Однако следует отметить, что традиционные подходы к степени полезности акклиматизационных работ в последнее время начали существенно пересматриваться. Так, Ю.Ю. Дгебуадзе (2000) оперирует понятием «биологическая инвазия», под которым понимает все случаи проникновения живых организмов в экосистемы, расположенные за пределами их естественного ареала. Он указывает, что в бывшем СССР долгое время считалось, что при экстенсивном ведении хозяйства можно существенно повысить выход рыбной продукции за счёт акклиматизационных работ, в связи с чем проводились масштабные переселения гидробионтов. При этом при подборе видов-акклиматизантов в первую очередь учитывали их продукционные и потребительские качества, потенциальные возможности их адаптации к абиотическим факторам нового водоёма. Одним из важнейших критериев также являлось наличие свободных экологических ниш. Но большая часть проводимых акклиматизационных мероприятий заканчивалась неудачей (а значит исследователи не учитывали какие-то важнейшие для акклиматизации факторы). Так, Е.В. Бурмакин (1963) указал, что в период с 1763 по 1957 гг. в 73% пересадок результат акклиматизации либо был отрицательным, либо не был выявлен вообще. Можно привести и ряд подобных примеров в отношении рыб, вселённых в водоёмы Северо-Западного Кавказа. Неудачей, несмотря на огромные финансовые расходы, закончилось вселение в регион белорыбицы, аральского усача, малоэффективны работы по заселению Азовского моря молодью каспийских осетровых. Акклиматизации белого и пёстрого толстолобиков, белого и чёрного амура, веслоноса не произошло. Их численность в естественных водоёмах может поддерживаться только путём искусственного воспроизводства. В тоже время в водоёмах появляются и быстро расселяются различные самоакклиматизанты.
Многие исследователи (Efford et al., 1997; Carlton, 1999; Mooney, 1999; Алимов, 2000; Матишов, 2000 и др.) в настоящее время сходятся во мнении, что вселение чужеродных видов в природные сообщества (даже с благими целями) представляет своего рода «биологическое загрязнение», сопоставимое по своим последствиям с другими видами загрязнения, а в некоторых случаях превышающее их. Новые виды могут подавлять или даже полностью вытеснять аборигенные формы, занимающие сходные экологические ниши, что приводит к снижению уровня биоразнообразия. Следует также учитывать другие негативные последствия вселения - распространение новых болезней и паразитов, ухудшение качества воды (Алимов и др., 2000; Горелов, 2000), «каскадный эффект» в сообществах (Carpenter et al., 1985).
Примеров негативного влияния вселенцев на местные экосистемы множество и, скорее всего, они даже перевешивают положительное влияние.
Приведём лишь некоторые примеры, касающиеся рыб.
1. Хрестоматийным примером стали последствия вселения в Великие американские озёра морской миноги (Petromyzon marinus). Долгое время она обитала только в оз. Онтарио, в притоках которого нерестилась. Проникновению миноги в другие озёра системы препятствовал Ниагарский водопад. Однако в 1829 г. для обеспечения судоходства был построен обводной канал, обеспечивший проход миноги в остальные водоёмы. В результате её хищничества был нанесён огромный ущерб местным ихтиоценозам и, прежде всего запасам сиговых, лососевых, чукучановых, уловы которых снизились в сотни раз (Элтон, 1960).
2. В 1950-х гг. британские ихтиологи решили вселить в африканское озеро Виктория нильского окуня (Lates niloticus), одного из самых крупных хищных видов рыб Африки. Целью подобной акклиматизации являлось занятие ниши крупного хищника. Из-за паводка несколько рыб, содержавшихся в прудах неподалёку от озера, попали в него. К началу 1970-х гг. окунь расселился по всему водоёму и существенно подорвал запасы и видовое разнообразие местных цихлид. Так, уловы хаплохромисов упали с 1200-2200 кг/га в середине 1970-х гг. до 200-400 кг в середине 1980-х. Местное население перешло от употребления в пищу вяленых на солнце хаплохромисов к добыче окуня. Перед употреблением в пищу этих крупных рыб нужно было термически обработать. Для этого люди начали вырубку лесов по берегам, что, усилило сток в воды озера биогенов и привело к его эвтрофированию, «цветению» воды. Конечным итогом акклиматизации нильского окуня стала трансформация местной экосистемы, резкое падение видового разнообразия ихтиофауны (Дгебуадзе, 2000).
Наиболее мощным антропогенным фактором расселения гидробионтов во всемирном масштабе в настоящее время считают их перевозки с помощью водного транспорта с балластными водами или на корпусах судов (Алимов и др., 2000). Это явление усилилось в последнее время, т.к. интенсифицировался сам процесс перевозок и изменились конструкции судов. Теперь балластные воды закачиваются в т.н. «второе дно», что позволяет попадающим вместе с ними гидробионтам избегать негативного воздействия токсических веществ и легче переносить перевозки. Мощностей для переработки балластных вод в большинстве российских портов недостаточно, что приводит к их сбросу непосредственно в море. С очень высокой долей вероятности можно утверждать, что именно с балластными водами судов в Чёрное море были завезены гребневики, брюхоногий моллюск рапана. Возможно, с ними же в море попали пойманные недавно у берегов Южного Крыма (Болтачёв, 2004) северная путассу (Micromesistius poutassou) и белопёрая рыба-бабочка.
В континентальных водоёмах России скорее всего важнейшими факторами расселения гидробионтов являлись целенаправленные акклиматизационные мероприятия и соединение каналами рек из различных речных систем (Алимов и др., 2000).
Акклиматизация - приспособление живых организмов к новым, непривычным для них климатогеографическим условиям. Без акклиматизационной подготовки проблематичен успех восхождения, особенно когда оно совершается на высоту более 5000. Влияние высоты практически у всех ощущается в первые дни после подъема уже на уровень 1500-2000 м, а основные проявления у многих начинают сказываться с 2500-3000 м. Это головная боль, тошнота, рвота (т.н. горная болезнь).
Кроме того не подготовленный к кислородному голоданию организм легче подвержен различным заболеваниям, работоспособность значительно снижается, а волевые качества сводятся к нулю. Часто эти симптомы проявляются при спуске c 3000-3500м, у других усиливаются на остановках и уменьшается при движении. Но полностью проходят только при понижении высоты и(или) приеме соответствующих лекарств. Как правило (если вы спустились) к утро эти симптомы проходят, но необходимо сделать день отдыха.
Кислород необходим всем органам и тканям человеческого тела при обмене веществ. Его расход прямо пропорционален активности организма. Перенос кислорода осуществляется красными кровяными тельцами -эритроцитами содержащими вещество - гемоглобин (оксигемоглобин), который благодаря разницы парциального давления кислорода в крови и в альвеолах насыщается кислородом и переносит его по организму. С высотой уменьшается парциального давления кислорода в воздухе (а не вообще количества кислорода, что является одним из широко распространенных заблуждений) это вызывает уменьшение насыщения кислородом крови .
Поэтому первой реакцией (первой фазой) организма на понижении внешнего давления является учащение пульса, повышение кровяного давления и усиление легочной вентиляции (т.н. гипервентиляция), что собственно и является причиной головной боли и пр.. Наступает расширение капилляров в тканях организма и прежде всего в мускулатуре. Кроме того, в кровообращение включается резервная кровь из селезенки и печени. Это компенсаторные процессы, протекающие в первой фазе акклиматизации, которая завершается, как правило, через 7-14 дней.
Во второй фазе акклиматизации недостаток кислорода начинает влиять на костный мозг, который начинает производить повышенное количество эритроцитов и вводить их в кровообращение. Так увеличивается гемоглобин в крови. Нормальное количество красных кровяных телец в 1 куб.мм - около 4,5-5,0 млн. у мужчин и 4,0-4,5 млн. у женщин. За период акклиматизации количество красных кровяных телец возрастает до 7,0 и даже до 8,0 млн. в 1 куб.мм, что приводит к увеличению гемоглобина. Установлено, что вторая фаза акклиматизации полностью завершается через три недели после начала работы на маршруте. Именно это должно определять планируемую длительность акклиматизационного периода и влиять на тактику акклиматизации. Достижение большой высоты может быть осуществлено надежно только после полной адаптации.
С физиологической точки зрения самая лучшая система заключается в постепенном достижении с грузом все большей высоты (лучше с кратковременным пребыванием на достигнутой высоте, например, ночевка), чередующемся со спуском вниз в базовый лагерь. Пребывание и отдых в базовом лагере один-два дня восстанавливают силы и закрепляют полученную акклиматизацию при сохранении относительного равновесия между акклиматизацией и детериорацией - исчерпывающим истощением организма, хотя альпинист никогда не достигнет уровня своей работоспособности при равнинных условиях. Отсюда следует необходимость стремления альпиниста к достижению очень высокой работоспособности в условиях равнины, а лучше среднегорья. Её за один год не приобретешь. Она вырабатывается, как правило, многолетней (в зависимости от генетических особенностей организма) тренировкой по общей и специальной физической и функциональной подготовках при регулярном периодическом медицинском контроле (в том числе с хотя бы ежегодной диспансеризацией для углубленного обследования) и повседневном постоянном самоконтроле самочувствия (пульс лежа после пробуждения, вес тела и т.п.).
Как показывает опыт высотных экспедиций, для лучшего восстановления сил при сохранении достигнутой акклиматизации после завершения установки промежуточных и штурмового лагерей целесообразен 2-4-дневный отдых на высоте на 500-1000 м ниже высоты базового лагеря. По современным представлениям 5300 м - это граница высокогорной зоны, в которой может еще наступить полная акклиматизация. В интервале от 7000 до 7800 м нельзя говорить о полной акклиматизации, поэтому организм начинает расходовать собственные резервы без возможности их пополнения. Альпинист может находиться в этой зоне до 4-5 дней, после чего необходимо спускаться ниже. Зона свыше 7800 м называется “зоной высотной смерти”. Пребывание в ней в течение 2-3 дней без кислородного аппарата вызывает быструю детериорацию. При разработке стратегии и тактики бескислородного высотного восхождения изложенное целесообразно принимать во внимание
Похожие работы
... похожи друг на друга. Поэтому целесообразно выделять группы сходных ареалов, типизировать ареалы по величине, характеру очертаний (и разрывов - для дизъюнктивных), географической приуроченности. Общая классификация типов распространения организмов пока отсутствует. Обычно такую классификацию проводят, исходя из конкретных целей исследования. Самые большие ареалы часто называют космополитическими. ...
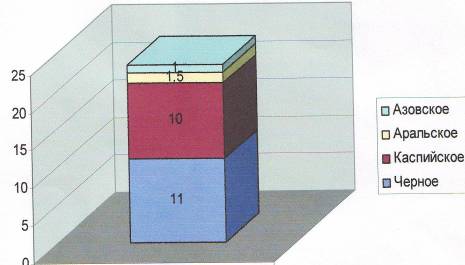
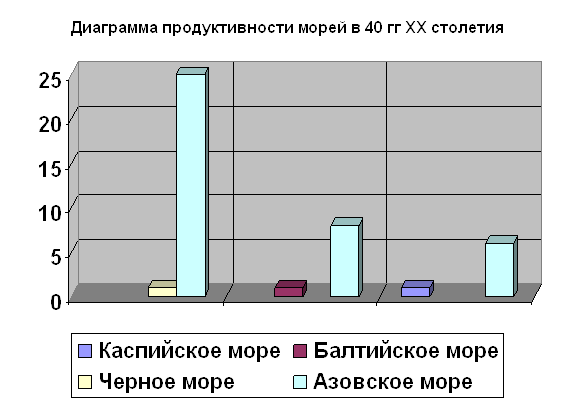
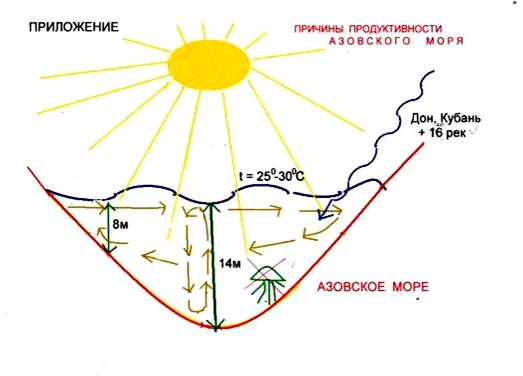
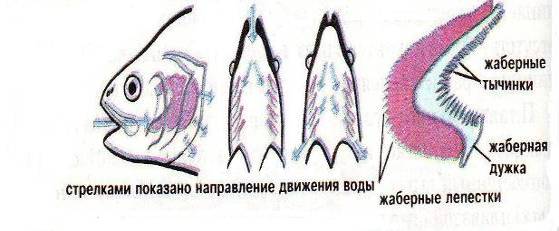
... во внешнем строении. Однако нужно помнить, что для точного установления вида недостаточно только морфологического критерия. Нужно учитывать и другие критерии вида. 2.2 Биология рыб семейства кефалевых Азово – Черноморского бассейна Рыбы – это группа холоднокровных позвоночных животных обтекаемой формы, живущих исключительно в воде. Для них характерны наличие парных (грудных и брюшных) и ...
... было бы ожидать в связи с обилием карстующихся пород. Более широко они развиты в южной части страны, где отсутствует сплошная мерзлота. Так, на Лено-Ангарском и Лено-Алданском плато имеется масса карстовых воронок, колодцев, слепых долин и т. д. С активным физическим выветриванием в условиях резко континентального климата связано обилие глыбово-каменистых россыпей, каменных потоков - курумов и ...
... биологических и химических средств защиты леса от хвое- и листогрызущих насекомых». Наставление разработано на основе обобщения результатов исследований по совершенствованию технологий изменения средств защиты леса от вредителей на базе современной авиационной техники. Данный документ рассматривает такие важные для лесозащитной практики вопросы, как определение целесообразности авиационной борьбы ...
0 комментариев