Вступ
1. Класифікація антигенів
2. Властивості антигенів
3. Поняття антигенної детермінанти або епітопа
4. Послідовні та переривчасті антигенні детермінанти, їх властивості
Висновок
Список використаних джерел
Вступ
Антиген - функціональне поняття. Ним може бути будь-яка речовина, що здатна викликати імунну відповідь.
Потужними антигенами є білки. Це зрозуміло, оскільки більшість процесів імунного розпізнання - це білок-білкові взаємодії. Білковими і пептидними антигенами є токсини бактерій та змій, білки вірусів та тканинні антигени. Добре розпізнаються вуглеводи, що входять до складу клітинних стінок бактерій. Більшість мембранних білків вищих тварин - це глікопротеїди, тому антигени груп крові, наприклад, є полісахаридними. Нуклеїнові кислоти самі по собі не є імуногенними, але у комплексі з білками стають такими. Наприклад, аутоімунна відповідь при захворюванні на червону вовчанку включає відповідь на нуклеопротеїди. Ліпіди можуть бути ефективно розпізнаними, якщо представлені з поверхневою клітинною молекулою CD1.
1. Класифікація антигенів
Класифікація антигенів.
1. Повні і неповні (гаптени). Антиген, якому властива антигенність, називається гантеном. Гаптени викликають імунну відповідь тільки після кон'югації з високомолекулярними носіями. Гаптенами можуть бути прості хімічні сполуки, наприклад, глюкоза або тринітрофенол. Вони стають імуногенними лише після з'єднання з білковим носієм, наприклад, з білковою молекулою. При відповіді на гаптен, який поєднаний з білком - носієм, В-лімфоцити розпізнають його, а Тh-лімфоцити - білкову молекулу.
2. Розчинні та корпускулярні. Розчинні (чужорідні білки, токсини, продукти деградації вірусів та бактерій) і корпускулярні (бактерії, віруси, чужорідні еукаріотичні клітини) антигени по-різному сприймаються імунною системою і, викликають різні форми імунної відповіді. В основі такого розділення лежить характер представлення антигенів клітинам імунної системи.
3. Екзогенні та ендогенні.
4. Т-залежні і Т-незалежні. Антигени, які викликають продукцію антитіл, можна поділити на тимусзалежні та тимуснезалежні. У процесі відповіді на тимусзалежні антигени В-лімфоцити потребують "допомоги" Т-лімфоцитів-хелперів. Серед цих антигенів можна виділити дві основні групи: цитокіновоопосередковані та власне Т-лімфоцитарні антигени.
Відповідь на тимуснезалежні антигени, яких є менше, ніж попередніх, не потребує допомоги з боку Г-лімфоцитів. До них серед інших належать: ліпополісахариди оболонок бактерій (LPS), декстран і очищений білок туберкуліну. Певні тимуснезалежні антигени містяться в багатьох вірусах, бактеріях і грибках.
Тимуснезалежні антигени теж можна поділити на дві групи:
1. Антигени з властивостями поліклональних активаторів В-лімфоцитів і здатністю до стимуляції імунної відповіді (наприклад, у новонароджених мишенят, у яких імунна реактивність ще не розвинена). Ці антигени передають В-лімфоцитам як сигнал І, так і сигнал II. Сюди належать LPS і декстран. Їх позначають символом ТІ-1 (thymus independent).
2. Антигени, які не мають рис поліклональних активаторів Я-лімфоцитів і не здатні викликати відповідь у новонароджених мишенят. Це полівалентні антигени, які діють вони не тільки безпосередньо на В-лімфоцити, але й опосередковано через цитокіни, що виділяються під їх впливом іншими клітинами, наприклад NK-клітини. До цієї групи належать Ficoll і полісахарид пневмококів. їх позначають символом ТІ-2. Ці антигени не є цілком "тимуснезалежні". За певних обставин, вони можуть безпосередньо стимулювати Т-лімфоцити.
5. Гетеро-, ізо(ало)- та аутоантигени.
2. Властивості антигенів1. Треба розрізняти поняття антигенності та імуногенності.
Антигенність (антигенна специфічність) - це здатність комплементарно зв'язуватися із антиген-специфічними рецепторами В і Т клітин. Вона притаманна майже всім відомим речовинам. Антигенами можуть бути пептиди, амінокислоти, вітаміни, і навіть АТФ, дінітрофенол чи іони металів. Але якщо просто ввести до організму одну з цих низькомолекулярних речовин, імунної відповіді не відбудеться.
Імуногенність - це здатність викликати імунну відповідь, тобто стимулювати цілу низку подій, необхідних для активації імунних клітин. Імуногенність залежить як від структури антигену (молекулярна вага, просторова будова), так і від стану імунної системи реципієнта (репертуар білків гістосумісності та Т-клітинних рецепторів).
Силу імунної відповіді до слабкого антигену можна підвищити за допомогою ад'ювантів - речовин, що сприяють неспецифічній стимуляції імунної системи: мінеральні олії (ад'юванти Фройнда), окис алюмінію.
2. Розчинні антигени сприймаються і представляються як екзогенні (зовнішні), і результатом їх розпізнання є активація В лімфоцитів і синтез антитіл. Антигени бактерій і вірусів часто синтезуються всередині інфікованих клітин і тому сприймаються імунною системою як ендогенні (внутрішні). Результатом їх розпізнання є активація цитотоксичних Т лімфоцитів, що знищують інфіковані клітини разом із інфекційним агентом.
3. Т-залежні антигени, їх більшість, потребують участі у своєму розпізнанні Т-лімфоцитів. Деякі антигени, що містять фрагменти, які багато разів повторюються (полісахариди бактерій), здатні давати достатній сигнал В лімфоцитам без участі Т і називаються Т-незалежними. Вони викликають тільки синтез антитіл.
4. По відношенню до організму реципієнта антигени діляться на ауто- (власні), ало- або ізо- (свого виду) та гетеро- або ксеноантигени (іншого виду). Потужність імунної відповіді півищується від ауто-до гетероантигенів.
Виявляється, що в межах одного антигену може знаходитися багато місць, до яких можуть приєднуватись антитіла. Це, так звані, епітопи або антигенні детермінанти. Епітопи однієї молекули можуть бути ідентичні або різні, можуть зв'язуватися з антитілами однакової або різної специфічності. Антиген, котрий містить багато епітопів, називається полівалентним. У молекулі білка найбільш антигенними є випуклі, рухомі щодо даної молекули фрагменти, які мають від'ємний заряд. Ні антиген, ні місце антитіла, що зв'язує антиген, не є статичними структурами. Вони характеризуються певною еластичністю щодо своєї форми.
Використовують також термін тканинні антигени. Мається на увазі комплекс антигенів, притаманних певному органу чи тканині. Базується на різних фізіологічних функціях органів, що визначає їх певну біохімічну особливість. Пухлинні антигени - це специфічні розчинні чи асоційовані з клітинами речовини, що з'являються в організмі під час пухлинного росту. Часто їх називають раково-ембріональними, оскільки вони характерні також для ембріонального розвитку і з'являються в організмі жінки під час вагітності. Такими антигенами є, наприклад, альфа-фетопротеїн і трофобласт-специфічний глобулін, їх визначення має значення для діагностики як вагітності, так і пухлинного росту.
3. Поняття антигенної детермінанти або епітопа
Ще у 30-х роках було показано, що молекула білка може зв'язати декілька молекул антитіл одночасно. У 50-х роках стало ясно, що антитіла взаємодіють із діскретними ділянками на поверхні білкової молекули. їх назвали антигенними детермінантами. Було сформульовано проблему: що складає антигенну детермінанту? Які властивості дозволяють тій чи іншій ділянці білка бути розпізнаній як чужерідній і викликати імунну відповідь?
Спочатку, як модель, було використано короткі синтетичні пептиди. Виявилося, що лінійні гомополімери амінокислот (тит (А1а-А1а)n) неімуногенні, але після кон'югації з білком-носієм поводять себе як гаптени, тобто мають антигенну специфічність. Розгілковані гетерополімери амінокислот високоімуногенні і викликають синтез антитіл до поверхневих ділянок молекули. Пептиди, взяті у впорядкованій або денатурованій формі, мали різну антигенну специфічність Якщо синтетичний антиген ніс заряджені групи, то антитіла до нього мали протилежний заряд.
Було зроблено висновків, що антигенні детермінанти знаходять ся на поверхні молекули, мають певну конформацію і несуть аміно кислотні залишки, здатні утворювати нековалентні зв'язки із антитілом.
Головні роботи по антигенній структурі глобулярних білків було проведено в 70-80-ті роки XX сторіччя. В результаті їх було з'ясовано що антигенна детермінанта чи епітоп - це відокремлена область на поверхні білкової молекули. У склад її входять 6-7 амінокислотних залишків. Не було знайдено зв'язків із якимось певними амінокислот ними залишками: у склад антигенних детермінант входили ті амінокислоти, що звичайно розташовані на поверхні білка. Виявилося, ще кожна антигенна детермінанта описує на поверхні білка лінію завдовжки 23-25 А і має детерміновані N і С кінці.
4. Послідовні та переривчасті антигенні детермінанти, їх властивостіРозрізняють послідовні (лінійні) та переривчасті (конформаційні) антигенні детермінанти.
Послідовні - визначаються порядком амінокислот. Антитіла де таких епітопів легко взаємодіють із лінійним пептидом такої їх послідовності. У чистому вигляді зустрічаються у фібрилярних білків та пептидів. У глобулярних білків поверхневі послідовні ділянки мають певну конформацію. Антитіла, отримані до пептидів, часте впізнають нативні білки, тобто можуть певним чином пристосовуватися до конформації поверхневих фрагментів.
Переривчасті антигенні детермінанти складаються із амінокислотних залишків, розташованих далеко один від одного у поліпептидному ланцюгу, але зближених за рахунок третинної структури білка перш за все дисульфідних зв'язків. Такі антигенні детермінанти не можна змоделювати лінійним пептидом.
Не всі амінокислоти, що входять до складу епітопу, мають однакове значення для розпізнання: як правило, специфічність визначається 1-2 залишками (імунодамінантними), а інші відіграють роль у підтримці належної конформації епітопу.
Як приклади, розглянемо антигенну структура міоглобіну кашалота та лізоциму курячого яйця - перших детально вивчених білкових антигенів.
Міоглобін - гемовмісний білок м'язів з молекулярною масою 18 КДа, складається із 153 амінокислотних залишків, не містить дисульфідних зв'язків. В молекулі міоглобіну було визначено п'ять лінійних епітопів: фрагменти 16-21, 56-62, 94-99, 113-119 і 146-151. У їх склад входили гідрофільні полярні амінокислоти.: Lys, Arg, Glu, His.
Лізоцим - фермент, що міститься у секреторних рідинах організму ссавців і у білку пташиних яєць, з молекулярною масою 14 КДа, має чотири дисульфідні зв'язки. У складі лізоциму було визначено три переривчасті антигенні детермінанти, що відповідали фрагментам:
22-34 і 113-116, зближених дисульфідним зв'язком 30-115;
62-68 і 74-96, зближених зв'язками 76-94 і 64-80;
6-13 і 126-129, зближених зв'язком 6-127.
Для вивчення цих антигенних детермінант було запропоновано спеціальний експериментальний підхід - синтез, що імітує поверхню. Так, для імітації переривчастого епітопу залишки, що було ідентифіковано як імунодомінантні, зшивали в єдиний пептид, поєднуючи окремі фрагменти за допомогою гліцинового спейсора:
116 113 114 34 33
Lys Asn Arg Phe Lys
Lys-Asn-Arg-Gly-Phe-Lys
Такий пептид ефективно блокував зв'язування специфічних антитіл із білком, тобто був схожий на природний переривчастий епітоп.
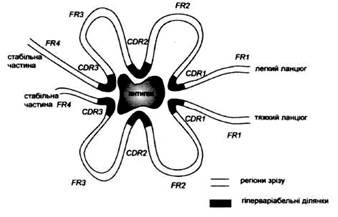
У 80-ті роки стало ясно, що вся поверхня білка може бути антигенною, тобто якщо для імунізації використовувати синтетичні пептиди, то можна отримати антитіла до будь-якої поверхневої ділянки. Однак при імунізації цілим білком антитіла утворювалися тільки до певних ділянок. Використання моноклональних антитіл чітко визначеної специфічності показало, що кожна антигенна детермінанта фактично складається із декількох потенційно антигенних ділянок, що перекриваються. Тепер такі епітопи стали називати більш вдалим терміном імунодомінантні області.
Природно, постало питання, які фактори визначають імунодомінантність.
Виходячи із визнаної функції імунної системи відрізняти "своє" від "чужого", першим принципом, покладеним в основу імунодомінантності, був принцип чужорідності антигену по відношенню до білків реципієнта. Щоб з'ясувати справедливість цього принципу вивчали серії гомологічних білків, тобто білків, що зустрічаються у багатьох організмів і відрізняються окремими амінокислотними замінами. Ідеальними для таких експериментів виявилися цитохроми с.
Цитохроми с - це гемовмісні білки дихального ланцюга мітохондрій з молекулярною масою 13 КДа, складаються із близько 100 амінокислотних залишків. Вони з'явились дуже рано в еволюції живого світу, перші цитохроми с зустрічаються у бактерій. Структура білка виявилася настільки вдалою, що збереглася у принципі до вищих тварин. Цитохроми ссавців відрізняються між собою окремими амінокислотними залишками, тобто можуть бути розглянуті як точкові мутанти. Було знайдено прямий зв'язок між імуногенністю цитохрому с та кількістю залишків, що відрізняли антиген від гомологічного цитохрому с реципієнта. Але стосовно специфічності антитіл, що вироблялися, цей зв'язок не виявився абсолютним. Так, кролі, імунізовані власним цитохромом, модифікованим глутаровим альдегідом, виробляли антитіла проти епітопів власного цитохрому. Коли тварин різних видів імунізували одним типом цитохрому, то антитіла вироблялися проти одних і тих самих ділянок. Тоді стали розглядати інший принцип імунодомінантності - зв'язок із структурними особливостями антигену, доступністю, зарядом, специфічним розташуванням на згині подіпептидного ланцюга. Було запропоновано алгоритми пошуку імунодомінантних ділянок за принципами гідрофільності та атомної рухливості. Подальші експерименти виявили зв'язок гідрофільності і рухомості із еволюційною варіабільністю: амінокислотні заміни, що закріпилися в еволюції, не повинні порушувати біологічні функції цитохрому с і тому локалізувалися саме у поверхневих, найбільш гнучких ділянках, де поява іншої амінокислоти найбільш безпечна і може бути компенсована за рахунок гнучкості молекули.
В результаті цих досліджень було дійшли висновку, що хоча вся поверхня білка в принципі може бути антигенною, за природної імунізації нативним білком антитіла утворюються тільки до певних епітопів, імунодомінантність яких визначається їх структурними особливостями, перш за все, гідрофільністю і атомною рухомістю (гнучкостю).
Антитіла (і В лімфоцити) зв'язують нативний антиген і впізнають на його поверхні так звані В-епітопи. Але ж у процесі імунної відповіді антиген впізнається і Т лімфоцитами. Більше того, саме специфічність Т лімфоцитів визначає ті імунодомінантні ділянки, що буде впізнано як В-епітопи. Ділянки антигену, що розпізнаються Т лімфоцитами, називаються Т-епітопами. їх положення і структура визначаються не так легко, як для В епітопів, тому що Т клітини впізнають антигени зовсім по-іншому.
1. Для розпізнання Т лімфоцитами антиген має бути процесованим (розщепленим). Процесинг відбувається всередині спеціалізованих клітин під дією протеолітичних ферментів. Спектр пептидів, що утворюються, залежить від типу протеаз, які відрізняються у різних типів клітин.
2. Процесований пептид має бути представленим у комплексі із білками головного комплексу гістосумісності: відбір антигенного пептиду залежить від структури цих білків, які є високо поліморфними і відрізняються навіть у різних особистостей одного виду.
3. Розпізнання представленого пептиду залежить від репертуару Т-клітинних рецепторів, який є результатом позитивного та негативного відбору у певного індивіда.
В результаті, Т-епітоп - це не обов'язково поверхнева структура; не конформаційно-залежний, а лінійний пептид. Його положення не пов'язане із гідрофільністю чи рухомістю поліпептидного ланцюга. Воно залежить як від структури нативного білка (потенційні сайти протеолізу, пептидні мотиви, що відповідають сайтам зв'язування білків гістосумісності), так і від стану імунної системи індивідуального реципієнта (репертуар білків гістосумісності і Т-клітинних рецепторів). Т-епітопи більше пов'язані із сайтами чужорідності антигену по відношенню до білків реципієнта, ніж В-епітопи, оскільки репертуар Т-рецепторів проходить більш суворий негативний відбір.
Визначення будови і локалізації В і Т епітопів представляє не тільки фундаментальний інтерес. Воно необхідне для створення ефективних вакцин і імунодіагностикумів.
Висновок
Імунна система здатна впізнати майже будь-яку речовину із середовища, що оточує макроорганізм. Для цього антиген має бути належним чином представлений імунним клітинам. В лімфоцити і антитіла впізнають конформаційно-залежні поверхневі епітопи, розташовані у місцях найбільшої гідрофільності та гнучкості поліпептидно-го ланцюга. Т лімфоцити впізнають внутрішні лінійні пептидні фрагменти, які утворюються в результаті протеолізу (процесингу) нативного антигену.
Антигенами називають речовини, здатні викликати імунну відповідь. Вони мають наступні властивості: а) імуногенність (здатність запустити специфічну відповідь), б) антигенність (здатність специфічно з'єднуватися з вільними або зв'язаними імуноглобулінами на поверхні В-лімфоцити, а також з рецепторами Г-лімфоцитів).
Антигени можуть бути неперервні, коли контактуючі з антитілом амінокислоти антигену містяться в одному відрізку білкового ланцюга, або уривчасті, коли ці амінокислоти перебувають в білковому ланцюгу на віддалі, але зближаються при певній конформації білка.
Розрізняють послідовні та переривчасті антигенні детермінанти. Послідовні - визначаються порядком амінокислот. Антитіла де таких епітопів легко взаємодіють із лінійним пептидом такої їх послідовності. Переривчасті антигенні детермінанти складаються із амінокислотних залишків, розташованих далеко один від одного у поліпептидному ланцюгу, але зближених за рахунок третинної структури білка перш за все дисульфідних зв'язків. Такі антигенні детермінанти не можна змоделювати лінійним пептидом.
Список використаних джерел
1. Большая медицинская энциклопедия. Т. 9. Гл. ред. Б.В. Петровский. – М.: Совецкая энциклопедия, 1978. – 483 с.
2. Вершигора А.Ю. Основы иммунологии. — К.: Вища шк. Головное изд-во, 1980. — 504 с.
3. Земское В.М. Основы общей микробиологии, вирусологии и иммунологии. — М.: Колос, 1977. — 311 с.
4. Иммунология: В 3-х т. Т.1. Пер. с англ. / Под ред. У. Пола. – М.: Мир, 1987 – 1988. – 476 с.
5. Петров Р.В. Иммунология. — М.: Медицина, 1982. — 368 с.
6. Скок М.В. Основи імунології. Курс лекцій. – К.: Фітосоціоцентр, 2002. – 152 с.
7. Якобисяк М. Імунологія. – Вінниця: Нова книга, 2004. – 672 с.
Похожие работы
... беременности у крыс / М. А. Волошин, О. Г. Кущ // Актуальные вопросы эволюционной, возрастной и экологической морфологии. – Белгород, 2006. – С. 34. (Самостійно проведено дослідження, обробка результатів, написання тез). АНОТАЦІЯ Кущ О.Г. Закономірності будови плаценти і лімфоїдної тканини, асоційованої з нею, протягом третього періоду вагітності (анатомо-експериментальне дослідження). – Рук
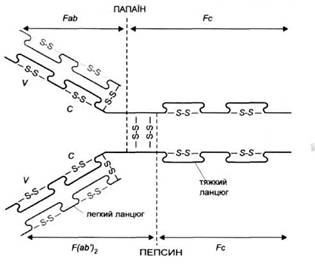
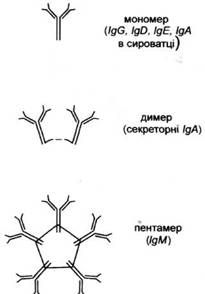
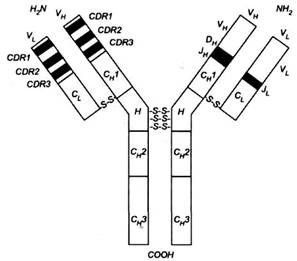

... і з крові багатьох донорів, призначаються при імунодефіцитах та важких вірусних і бактерійних захворюваннях. Еквівалентом і попередником IgG у птахів, плазунів і земноводних є так звані антитіла lg Y. 2.4 Імуноглобулін М Антитіла цього класу синтезуються в початковій фазі імунної відповіді; також ці антитіла перші синтезуються в онтогенезі. При первинній імунній відповіді першими виді ...
... спектру дії Наявність загального родоспецифічного антигена. Все це визначає самостійне положення цих мікроорганізмів серед прокаріот [5]. 3. Характеристика будови та морфології представників роду Chlamydia 3.1 Морфологічні особливості Хламідії є групою прокаріотних мікроорганізмів і мають вид дрібних грамнегативних коків. Добре видні в не зафарбованому стані при мікроскопії ...
... живлення, дихання, ріст і розвиток, розмноження, реакції на зовнішні подразники, пластичність, інтенсивність взаємодії з факторами середовища. Фізіологія являється науковою основою промислового використання мікроорганізмів у мікробіологічних виробництвах біологічно активних речовин (БАР), ферментів, вітамінів, антибіотиків, амінокислот, органічних кислот. Мікроорганізми здатні до синтезу ...
0 комментариев