Снежинский Физико-Технический Институт НИЯУ «МИФИ»
Снежинский Политехнический Колледж
Доклад по биологии на тему:
«Естественный отбор»
Выполнила: студентка 1 курса
Группы Ф-18Д
Якунина Елена
Проверила: Будалова И.Б.
г. Снежинск 2009
Содержание
Естественный отбор
а)Дестабилизирующий отбор
б)Половой отбор
в)Групповой отбор
г)Направленный отбор (движущий)
д)Стабилизирующий отбор
е)Дизруптивный (расчленяющий) отбор
Заключение
Список литературы
Естественный отбор
Естественный отбор – результат борьбы за существование; он основывается на преимущественном выживании и оставлении потомства с наиболее приспособленными особями каждого вида и гибели менее приспособленных организмов.
Мутационный процесс, колебания численности популяций, изоляция создают генетическую неоднородность внутри вида. Но их действие ненаправленно. Эволюция же – процесс направленный, связанный с выработкой приспособлений, с прогрессивным усложнением строения и функций животных и растений. Существует лишь один направленный эволюционный фактор – естественный отбор.
Под действие отбора могут попасть либо определенные особи, либо целые группы. В результате группового отбора часто накапливаются признаки и свойства, невыгодные для отдельной особи, но полезные для популяции и целого вида (ужалившая пчела гибнет, но, нападая на врага, она сохраняет семью). В любом случае отбор сохраняет наиболее приспособленные к данной среде организмы и действует в пределах популяций. Таким образом, именно популяции – поле действия отбора.
Под естественным отбором нужно понимать избирательное (дифференциальное) воспроизведение генотипов (или генных комплексов). В процессе естественного отбора важны не столько выживание или гибель особей, сколько их дифференциальное размножение. Успех в размножении разных особей может служить объективным генетико-эволюционным критерием естественного отбора. Биологическое значение особи, давшей потомство, определяется вкладом ее генотипа в генофонд популяции. Отбор из поколения в поколение по фенотипам ведет к отбору генотипов, так как потомкам передаются не признаки, а генные комплексы. Для эволюции имеют значение не только генотипы, но и фенотипы и фенотипическая изменчивость.
В процессе экспрессии ген может оказывать влияние на многие признаки. Поэтому в сферу действия отбора могут включаться не только свойства, повышающие вероятность оставления потомства, но и признаки, которые не имеют прямого отношения к воспроизводству. Они отбираются опосредованно в результате корреляций.
а)Дестабилизирующий отборДестабилизирующий отбор - это разрушение корреляционных связей в организме при интенсивном отборе в каждом определенном направлении. В пример можно привести случай, когда отбор, направленный на уменьшение агрессивности, приводит к дестабилизации цикла размножения.
Стабилизирующий отбор сужает норму реакции. Однако в природе нередки случаи, когда экологическая ниша вида со временем может оказаться более широкой. В этом случае селективное преимущество получают особи и популяции с более широкой нормой реакции, сохраняющие вместе с тем то же среднее значение признака. Эта форма естественного отбора была впервые описана американским эволюционистом Джорджем Г. Симпсоном под названием центробежного отбора. В итоге идет процесс, обратный стабилизирующему отбору: преимущество получают мутации с более широкой нормой реакции.


Так, популяции озёрных лягушек, живущие в прудах с разнородной освещенностью, с чередованием участков, заросших ряской, тростником, рогозом, с «окнами» открытой воды, характеризуются широким диапазоном изменчивости окраски (итог дестабилизирующей формы естественного отбора). Наоборот, в водоемах с однородной освещенностью и окраской (пруды, сплошь заросшие ряской, или открытые пруды) диапазон изменчивости окраски лягушек узок (итог действия стабилизирующей формы естественного отбора).
Таким образом, дестабилизирующая форма отбора ведет к расширению нормы реакции.
б) Половой отбор
Половой отбор - естественный отбор внутри одного пола, направленный на выработку признаков, дающих преимущественно возможность оставить наибольшее количество потомков.
У самцов многих видов обнаруживаются явно выраженные вторичные половые признаки, которые на первый взгляд кажутся неадаптивными: хвост павлина, яркие перья райских птиц и попугаев, алые гребни петухов, феерические цвета тропических рыбок, песни птиц и лягушек, и т.п. Многие из этих особенностей осложняют жизнь их носителей, делают их легко заметными для хищников. Казалось бы, эти признаки не дают никаких преимуществ их носителям в борьбе за существование, и тем не менее они очень широко распространены в природе. Какую роль в их возникновении и распространении сыграл естественный отбор?
Мы уже знаем, что выживание организмов является важным, но не единственным компонентом естественного отбора. Другим важнейшим компонентом является привлекательность для особей противоположного пола. Ч.Дарвин назвал это явление половым отбором. Впервые он упомянул эту форму отбора в «Происхождении видов», а затем подробно проанализировал ее в книге «Происхождение человека и половой отбор». Он считал, что «эта форма отбора определяется не борьбой за существование в отношениях органических существ между собою или с внешними условиями, но соперничеством между особями одного пола, обычно самцами, за обладание особями другого пола».
Половой отбор - это естественный отбор на успех в размножении. Признаки, которые снижают жизнеспособность их носителей, могут возникать и распространяться, если преимущества, которые они дают в успехе размножения значительно выше, чем их недостатки для выживания. Самец, который живет недолго, но нравится самкам и поэтому производит много потомков, имеет гораздо более высокую совокупную приспособленность, чем тот, что живет долго, но оставляет мало потомков. У многих видов животных подавляющее большинство самцов вовсе не участвует в размножении. В каждом поколении между самцами возникает жесточайшая конкуренция за самок. Эта конкуренция может быть прямой, и проявляться в виде борьбы за территории или турнирных боев. Она может происходить и в косвенной форме и быть обусловленной выбором самок. В тех случаях, когда самки выбирают самцов, конкуренция самцов проявляется в демонстрации их яркого внешнего вида или сложного поведения ухаживания. Самки выбирают тех самцов, которые им больше всего нравятся. Как правило, это наиболее яркие самцы. Но почему самкам нравятся яркие самцы?

Рис. 7. Яркая расцветка птиц возникает в эволюции благодаря половому отбору.
Приспособленность самки зависит о того, насколько объективно она способна оценить потенциальную приспособленность будущего отца своих детей. Она должна выбрать такого самца, сыновья которого будут обладать высокой приспособленностью и привлекательностью для самок.
Было предложено две основные гипотезы о механизмах полового отбора.
Согласно гипотезе «хороших генов» самка «рассуждает» следующим образом: «Если этот самец, несмотря на его яркое оперение и длинный хвост, каким-то образом умудрился не погибнуть в лапах хищника и дожить до половой зрелости, то, следовательно, он обладает хорошими генами, которые позволили ему это сделать. Значит, его стоит выбрать в качестве отца для своих детей: он передаст им свои хорошие гены». Яркость окраски перьев и гребней у птиц, чешуи и хвоста у рыб является надежной характеристикой здоровья. Так, на рыбке корюшке было показано, что чем ярче окраска самца, тем более он привлекателен для самок и тем менее он заражен паразитами. Яркость гребня у петухов напрямую зависит от содержания тестостерона в крови и от степени зараженности паразитами. Чем выше уровень тестостерона, тем ярче гребень, чем больше паразитов, тем он бледнее. Известно, также, что тестостерон угнетает иммунную систему. Следовательно, самцы с ярким гребнем обладают такой эффективной иммунной системой, которая обеспечивает их устойчивость к паразитам, несмотря на высокий уровень тестостерона. Выбирая ярких самцов, самки выбирают хорошие гены для своих потомков.
Согласно гипотезе «привлекательных сыновей» логика выбора самок несколько иная. Если яркие самцы, по каким бы то ни было причинам, являются привлекательными для самок, то стоит выбирать яркого отца для своих будущих сыновей, потому что его сыновья унаследуют гены яркой окраски и будут привлекательными для самок в следующем поколении. Таким образом, возникает положительная обратная связь, которая приводит к тому, что из поколения в поколение яркость оперения самцов все более и более усиливается. Процесс идет по нарастающей до тех пор, пока не достигнет предела жизнеспособности. Представим себе ситуацию, когда самки выбирают самцов с более длинным хвостом. Длиннохвостые производят больше потомков, чем самцы с короткими и средними хвостами. Из поколения в поколение длина хвоста увеличивается, потому что самки выбирают самцов не с определенным размером хвоста, но с большим, чем в среднем размером. В конце концов, хвост достигает такой длины, когда его вред для жизнеспособности самца уравновешивается его привлекательностью в глазах самок.
Объясняя эти гипотезы, мы старались понять логику действия самок птиц. Может создаться впечатление, что мы слишком много от них ожидаем, что такие сложные расчеты приспособленности им вряд ли доступны. На самом деле, в выборе самцов самки не более и не менее логичны, чем во всем остальном их поведении. Когда животное чувствует жажду, оно не рассуждает, что ему следует попить воды, для того чтобы восстановить водно-солевой баланс в организме – оно идет на водопой, потому что чувствует жажду. Когда рабочая пчела жалит хищника, напавшего на улей, она не вычисляет, насколько этим своим самопожертвованием она повышает совокупную приспособленность своих сестер – она следует инстинкту. Точно так же и самки, выбирая ярких самцов, следуют своим инстинктами – им нравятся яркие хвосты. Все те, кому инстинкт подсказывал иное поведение, все они не оставили потомства. Таким образом, мы обсуждали не логику самок, а логику борьбы за существование и естественного отбора – слепого и автоматического процесса, который, действуя постоянно из поколения в поколение, сформировал все то удивительное разнообразие форм, окрасок и инстинктов, которое мы наблюдаем в мире живой природы.
в) Групповой отбор
Групповой отбор часто называют также групповым отбором, представляет собой дифференциальное размножение разных локальных популяций. В.Райт сравнивает популяционные системы двух типов — большую непрерывную популяцию и ряд мелких полуизолированных колоний — в отношении теоретической эффективности отбора. Предполагается, что общая величина обеих популяционных систем одинакова и организмы свободно скрещиваются между собой.
В большой непрерывной популяции отбор относительно неэффективен в смысле повышения частоты благоприятных, но редких рецессивных мутаций. Кроме того, любой тенденции к повышению частоты какого-либо благоприятного аллеля в одной части данной обширной популяции противодействует скрещивание с соседними субпопуляциями, в которых этот аллель редок. Подобным же образом благоприятные новые генные сочетания, которым удалось образоваться в какой-нибудь локальной доле данной популяции, разбиваются на части и элиминируются в результате скрещивания с особями соседних долей.
Все эти трудности в значительной степени устраняются в популяционной системе, по своей структуре напоминающей ряд отдельных островков. Здесь отбор или отбор совместно с дрейфом генов может быстро и эффективно повысить частоту какого-либо редкого благоприятного аллеля в одной или нескольких мелких колониях. Новые благоприятные сочетания генов также могут легко закрепиться в одной или нескольких мелких колониях. Изоляция защищает генофонды этих колонии от «затопления» в результате миграции из других, не обладающих столь благоприятными генами колоний, и от скрещивания с ними. До этого момента в модель был включен только индивидуальный отбор или — для некоторых колоний — индивидуальный отбор в сочетании с дрейфом генов.
Допустим теперь, что среда, в которой находится данная популяционная система, изменилась, в результате чего адаптивность прежних генотипов понизилась. В новой среде новые благоприятные гены или сочетания генов, закрепившиеся в некоторых колониях, обладают высокой потенциальной адаптивной ценностью для популяционной системы в целом. Теперь имеются все условия для того, чтобы вступил в действие групповой отбор. Менее приспособленные колонии постепенно сокращаются и вымирают, а колонии, которые более приспособлены, расширяются и замещают их по всей области, занимаемой данной популяционной системой. Такая подразделившаяся популяционная система приобретает новый набор приспособительных признаков в результате индивидуального отбора в пределах некоторых колоний, за которым следует дифференциальное размножение разных колоний. Сочетание группового и индивидуального отбора может привести к результатам, которые не могут быть достигнуты за счет одного лишь индивидуального отбора.
Установлено, что групповой отбор — процесс второго порядка, дополняющий главный процесс индивидуального отбора. Будучи процессом второго порядка, групповой отбор должен протекать медленно, вероятно, гораздо медленнее, чем индивидуальный отбор. Обновление популяций требует больше времени, чем обновление особей.
Концепция группового отбора встретила широкое признание в некоторых кругах, но была отвергнута другими учеными.. Они утверждают, что различные возможные модели индивидуального отбора способны вызывать все эффекты, приписываемые групповому отбору. Уэйд провел ряд селекционных экспериментов с мучным хрущаком (Tribolium castaneum), с тем чтобы выяснить эффективность группового отбора, и обнаружил, что жуки реагировали на отбор этого типа. Кроме того, когда на какой-либо признак одновременно действуют индивидуальный и групповой отбор и притом в одном и том же направлении, скорость изменения этого признака выше, чем в случае одного только индивидуального отбора (Даже умеренная иммиграция (6 и 12%) не препятствует дифференциации популяций, вызываемой групповым отбором.

Одна из особенностей органического мира, которую трудно объяснить на основе индивидуального отбора, но можно рассматривать как результат группового отбора, — это половое размножение. Хотя и были созданы модели, в которых половому размножению благоприятствует индивидуальный отбор, однако они представляются нереалистичными. Половое размножение — это тот процесс, который создаёт рекомбинационную изменчивость в скрещивающихся популяциях. От полового размножения выигрывают не родительские генотипы, распадающиеся в процессе рекомбинации, а популяция будущих поколений, у которых возрастает запас изменчивости. Это подразумевает участие в качестве одного из факторов селективного процесса на популяционном уровне.
г) Направленный отбор (движущий)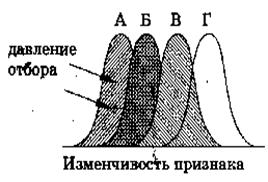
Рис. 1. Движущая форма естественного отбора
Направленный отбор (движущий) был описан еще Ч.Дарвином, а современное учение о движущем отборе разработано Дж. Симпсоном.
Суть этой формы отбора заключается в том, что он вызывает прогрессивное или направленное в одну сторону изменение генетического состава популяций, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. Он происходит в тех случаях, когда популяция находится в процессе приспособления к новой среде или же когда происходит постепенное изменение среды, а вслед за ней и постепенное изменение популяции.
При длительном изменении внешней среды преимущество в жизнедеятельности и размножении может получить часть особей вида с некоторыми отклонениями от средней нормы. Это приведет к изменению генетической структуры, возникновению эволюционно новых приспособлений и перестройке организации вида. Вариационная кривая смещается в направлении приспособления к новым условиям существования.
Такой отбор приводит к появлению новой нормы вместо старой, переставшей соответствовать новым условиям. Изменение признака может происходить как в сторону его усиления, так и в сторону ослабления. Как приобретение нового признака, так и утрата какого-либо признака – результаты действия движущей формы отбора. Редукция органов, потерявших свое функциональное значение, происходит именно таким путем. Примерами являются некоторые бескрылые птицы и насекомые, растения-паразиты без корней и листьев, ленточные черви без пищеварительной системы, потемнение окраски бабочки березовой пяденицы в развитых индустриальных районах Англии. До 1848 года описанные в Англии формы бабочек березовой пяденицы имели бледно-кремовую окраску с черными точками и отдельными темными пятнышками. В 1848 году в Манчестере была обнаружена черная форма этой бабочки, а к 1895 году популяция березовой пяденицы в Манчестере состояла на 98% из темноокрашенных бабочек. Эта «меланическая» форма появилась в результате возникновения случайных мутаций, причем фенотип мутантных особей обладал в промышленных районах большим преимуществом по сравнению со светлоокрашенными формами.
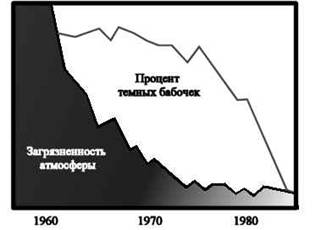
Рис 2.Зависимость частоты темных форм березовой пяденицы от степени загрязненности атмосферы
Светлоокрашенные формы были незаметны на стволах берез, покрытых лишайниками. С интенсивным развитием промышленности диоксид серы, образующийся при сжигании угля, вызывал гибель лишайников в промышленных районах, и в результате обнаружилась темная кора деревьев. На темном фоне светлоокрашенные пяденицы склевывались малиновками и дроздами, а выживали и успешно размножались меланические формы, которые на темном фоне менее заметны. За последние 100 лет у более чем 80 видов бабочек появились темные формы. Это явление известно теперь под названием индустриального (промышленного) меланизма. Движущий отбор приводит к появлению нового вида.


Рис. 3. Индустриальный меланизм. Темные формы бабочек незаметны на темных стволах , а светлые – на светлых.
Насекомые, ящерицы и ряд других обитателей травы имеют зеленую или бурую окраску, обитатели пустыни – цвет песка. Шерсть животных, живущих в лесах, например леопарда, расцвечена небольшими пятнами, напоминающими солнечные блики, а у тигра имитирует окраску и тень от стеблей камыша или тростника. Такая окраска получила название покровительственной.
У хищников она закрепилась благодаря тому, что ее обладатели незаметно могли подкрадываться к добыче, а у организмов, являющихся добычей, - вследствие того, что жертва оставалась менее заметной для хищников. Как же она появилась? Многочисленные мутации давали и дают большое разнообразие форм, отличающихся по окраске. В ряде случаев расцветка животного оказывалась близкой к фону окружающей среды, т.е. скрывала животное, играла роль покровительственной. Те животные, у которых покровительственная окраска была выражена слабо, оставались без пищи либо сами становились жертвой, а их сородичи, обладающие лучшей покровительственной окраской, выходили победителями в межвидовой борьбе за существование.
Направленный отбор лежит в основе искусственного отбора, при котором избирательное скрещивание особей, обладающих желательными фенотипическими признаками, повышает частоту этих признаков в популяции. В ряде экспериментов Фальконер выбирал из популяции шестинедельных мышей самых тяжелых особей и давал им спариваться между собой. То же самое он проделывал с самыми легкими мышами. Такое избирательное скрещивание по признаку массы тела привело к созданию двух популяций, в одной из которых масса возрастала, а в другой – уменьшалась.
После прекращения селекции ни та, ни другая группа не вернулась к первоначальной массе (примерно 22 грамма). Это показывает, что искусственный отбор по фенотипическим признакам привел к некоторому генотипическому отбору и частичной утрате обеими популяциями каких-то аллелей.
д) Стабилизирующий отбор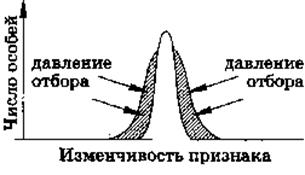
Рис. 4. Стабилизирующая форма естественного отбора
Стабилизирующий отбор в относительно постоянных условиях среды естественный отбор направлен против особей, признаки которых отклоняются от средней нормы в ту или другую сторону.
Стабилизирующий отбор сохраняет то состояние популяции, которое обеспечивает ее максимальную приспособленность в постоянных условиях существования. В каждом поколении удаляются особи, отклоняющиеся от среднего оптимального значения по приспособительным признакам.
Описано множество примеров действия стабилизующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью.

Однако наблюдения над природными популяциями птиц и млекопитающих показывают, что это не так. Чем больше птенцов или детенышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью.
Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих новорожденные с очень низким и очень высоким весом чаше погибают при рождении или в первые недели жизни, чем новорожденные со средним весом. Учет размера крыльев у птиц, погибших после бури, показал, что большинство из них имели слишком маленькие или слишком большие крылья. И в этом случае наиболее приспособленными оказались средние особи.
В чем причина постоянного появления малоприспособленных форм в постоянных условиях существования? Почему естественный отбор не способен раз и навсегда очистить популяцию от нежелательных уклоняющихся форм? Причина не только и не столько в постоянном возникновении все новых и новых мутаций. Причина в том, что часто наиболее приспособленными оказываются гетерозиготные генотипы. При скрещивании они постоянно дают расщепление и в их потомстве появляются гомозиготные потомки со сниженной приспособленностью. Это явление получило название сбалансированный полиморфизм.
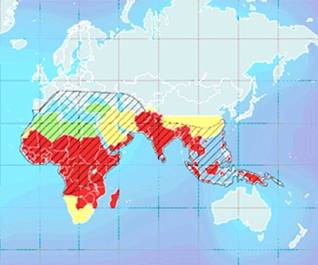
Рис.5. Карта распределения серповидно-клеточной анемии в малярийных районах. Цветами обозначены малярийные районы. В заштрихованной области обнаруживается высокая частота серповидно-клеточной анемии
Наиболее широко известным примером такого полиморфизма является серповидно-клеточная анемия. Это тяжелое заболевание крови возникает у людей гомозиготных по мутантному аллею гемоглобина (HbS) и приводит к их гибели в раннем возрасте. В большинстве человеческих популяций частота этого аллея очень низка и приблизительно равна частоте его возникновения за счет мутаций. Однако он довольно часто встречается в тех районах мира, где распространена малярия. Оказалось, что гетерозиготы по HbSимеют более высокую устойчивость к малярии, чем гомозиготы по нормальному аллею. Благодаря этому в популяциях, населяющих малярийные районы, создается и стабильно поддерживается гетерозиготность по этому летальному в гомозиготе аллею.
Стабилизирующий отбор является механизмом накопления изменчивости в природных популяциях. Первым на эту особенность стабилизирующего отбора обратил внимание выдающийся ученый И.И.Шмальгаузен. Он показал, что даже в стабильных условиях существования не прекращается ни естественный отбор, ни эволюция. Даже оставаясь фенотипически неизменной, популяция не перестает эволюционировать. Её генетический состав постоянно меняется. Стабилизирующий отбор создает такие генетические системы, которые обеспечивают формирование сходных оптимальных фенотипов на базе самых разнообразных генотипов. Такие генетические механизмы как доминирование, эпистаз, комплементарное действие генов, неполная пенетрантность и другие средства скрывания генетической изменчивости обязаны своим существованием стабилизирующему отбору.
Стабилизирующая форма естественного отбора предохраняет сложившийся генотип от разрушающего влияния мутационного процесса, чем объясняется, например, существование таких древних форм, как гаттерия, гинкго.
Благодаря стабилизирующему отбору до наших дней сохранились «живые ископаемые», обитающие в относительно постоянных условиях внешней среды:
1. гаттерия, носящая черты пресмыкающихся мезозойской эры;
2. латимерия, потомок кистеперых рыб, широко распространенных в палеозойскую эру;
3. североамериканский опоссум – сумчатое животное, известное с мелового периода;
4. голосеменное растение гинко, сходное с древесными формами, вымерших в юрском периоде мезозойской эры.
Стабилизирующая форма отбора действует до тех пор, пока сохраняются условия, повлекшие образование того или иного признака или свойства.
Здесь важно отметить, что постоянство условий не означает их неизменности. В течение года экологические условия регулярно меняются. Стабилизирующий отбор адаптирует популяции к этим сезонным изменениям. К ним приурочиваются циклы размножения, таким образом, чтобы молодняк рождался в тот сезон года, когда ресурсы пищи максимальны. Все отклонения от этого оптимального, воспроизводимого из года в год цикла, устраняются стабилизирующим отбором. Потомки, родившиеся слишком рано, гибнут от бескормицы, слишком поздно – не успевают подготовиться к зиме. Как животные и растения узнают о наступлении зимы? По наступлению заморозков? Нет, это не слишком надежный указатель. Кратковременные флуктуации температуры могут быть очень обманчивы. Если в какой-то год потеплело раньше обычного, то это вовсе не значит, что пришла весна. Те, кто слишком поспешно среагируют на этот ненадежный сигнал, рискуют остаться без потомства. Лучше дождаться более надежного знака весны – увеличения светового дня. У большинства видов животных, именно этот сигнал запускает механизмы сезонных изменений жизненно важных функций: циклы размножения, линьки, миграций и др. И.И. Шмальгаузен убедительно показал, что эти универсальные адаптации возникают в результате стабилизирующего отбора.
Таким образом, стабилизирующий отбор, отметая отклонения от нормы, активно формирует генетические механизмы, которые обеспечивают стабильное развитие организмов и формирование оптимальных фенотипов на базе разнообразных генотипов. Он обеспечивает устойчивое функционирование организмов в широком спектре привычных для вида колебаний внешних условий.
е)Дизруптивный (расчленяющий) отбор
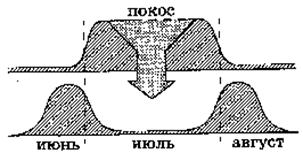
Рис. 6. Дизруптивная форма естественного отбора
Дизруптивный (расчленяющий) отбор благоприятствует сохранению крайних типов и элиминации промежуточных. В результате он приводит к сохранению и усилению полиморфизма. Разрывающий отбор действует в разнообразных условиях среды, встречающихся на одной территории, и поддерживает несколько фенотипически различных форм за счет особей со средней нормой. Если условия среды настолько изменились, что основная масса вида утрачивает приспособленность, то преимущество приобретают особи с крайними отклонениями от средней нормы. Такие формы быстро размножаются и на основе одной группы формируется несколько новых.
Моделью дизруптивного отбора может быть ситуация возникновения карликовых рас хищных рыб в малокормном водоеме. Часто щурятам-сеголеткам не хватает корма в виде мальков рыб. В этом случае преимущество получают самые быстрорастущие, которые очень быстро достигают размеров, позволяющих поедать своих собратьев. С другой стороны, в выгодном положении окажутся щурята с максимальной задержкой скорости роста, так как мелкие размеры позволяют им длительное время оставаться планктофагами. Подобная ситуация через стабилизирующий отбор может привести к возникновению двух рас хищных рыб.
Интересный пример приводит Дарвин относительно насекомых – обитателей небольших океанических островов. Они прекрасно летают или совсем лишены крыльев. По-видимому, насекомые внезапными порывами ветра уносились в море; сохранились лишь те, которые могли либо противостоять ветру, либо совершенно не летали. Отбор в этом направлении привел к тому, что на острове Мадейра из 550 видов жуков 200 нелетающие.
Еще один пример: в лесах, где почвы коричневого цвета особи земляной улитки чаще имеют коричневую и розовую окраску раковин, на участках с грубой и желтой травой преобладает желтая окраска и т.п.

Популяции, приспособленные к экологически несходным местообитаниям, могут занимать смежные географические области; например, в прибрежных областях Калифорнии растение Gilia achilleaefolia представлено двумя расами. Одна раса – «солнечная» – растет на открытых травянистых южных склонах, тогда как «теневая» раса встречается в тенистых дубовых лесах и рощах секвойи. Эти расы различаются по величине лепестков – признаку, детерминированному генетически.
Основной результат этого отбора заключается в формировании полиморфизма популяции, т.е. наличии нескольких, различающихся по какому-либо признаку групп или в изоляции популяций, отличающихся по своим свойствам, что может быть причиной дивергенции.
Заключение
Как и другие элементарные эволюционные факторы, естественный отбор вызывает изменения в соотношении аллелей в генофондах популяций. В эволюции естественному отбору принадлежит творческая роль. Исключая из репродукции генотипы с малой приспособительной ценностью, сохраняя благоприятные генные комбинации разного достоинства, он преобразует картину генотипической изменчивости, складывающуюся первоначально под действием случайных факторов, в биологически целесообразном направлении.
Список литературы
1) Власова З.А. Биология. Справочник студента – Москва, 1997
2) Грин Н. Биология – Москва, 2003
3) Камлюк Л.В. Биология в вопросах и ответах – Минск, 1994
4) Лемеза Н.А. Пособие по биологии – Минск, 1998
Похожие работы


... , которые в большей мере соответствуют данным условиям. Естественный отбор и элиминация части особей вида неразрывно связаны между собой и являются необходимым условием эволюции видов в природе. Схема действия естественного отбора в системе вида по Дарвину сводится к следующему: 1) Изменчивость свойственна любой группе животных и растений, и организмы отличаются друг от друга во многих ...
... , открытом ветрам со всех сторон, не оказалось ни одного дерева. То же было и на островах Кергеленских, где почти все растения стелются по земле, а самое высокое едва достигает 1 м. Многие растения образуют плотные дерновые подушки. Растения высокие или со слабым укоренением стеблей уничтожались здесь в процессе многовекового отбора. Сравнение действия искусственного и естественного отбора.





... их к изменившимся условиям, стремились бы сохраниться, и естественный отбор имел бы полный простор для своего улучшающего действия. 1. ЕСТЕСТВЕННЫЙ ОТБОР КАК ЭЛЕМЕНТАРНЫЙ ЭВОЛЮЦИОННЫЙ ФАКТОР Сохранение благоприятных индивидуальных различий и изменений и уничтожение вредных я назвал естественным отбором или переживанием наиболее приспособленных Ч. Дарвин В современном понимании ...
... , которая по их мнению должна снять это возражение, сама по себе не адекватна задаче объяснения эволюции. Иначе говоря, генетика не подтверждает эволюционную теорию. Естественный отбор Однако о правильности постулатов генетики должно быть сказано гораздо больше. Далекие от того, чтобы поддержать эволюционную теорию, исследования последних семидесяти лет приводят к единственному выводу: ...
0 комментариев