С.Ю. Афонькин, Г.П. Пинаев
В сумерки на чердаке сарая заработало штурвальное колесо. Один за одним натягивались крепкие веревочные провода, передавая туда, куда надо, и те, что надо, сигналы.
А.Гайдар. «Тимур и его команда»
Основы коммуникации
Для нормальной жизнедеятельности любого организма составляющие его клетки, подобно людям в социуме, должны чутко реагировать на меняющуюся ситуацию, регулируя свою работу в зависимости от окружающих условий и текущих потребностей многоклеточного государства. Изменение функций клетки происходит при появлении или исчезновении в ней определенных белков или изменении активностей уже существующих. Регуляция активностей белков происходит путем изменения их пространственной структуры за счет присоединения или диссоциации ионов металлов, фосфатных, гидроксильных или метильных групп, а также взаимопревращения SH-групп и S–S-связей, способных образовывать внутрибелковые сшивки. Эти группы и малые молекулы играют роль своеобразных молекулярных «зажимов» и «фиксаторов», добавление или удаление которых обратимо изменяет активности ферментов.
Клетка изменяет активности своих ферментов в ответ на получаемые ею из внешней среды специфические сигналы. Эти сигналы представляют собой определенные низкомолекулярные вещества (лиганды), связывающиеся со специальными участками клеточной поверхности – рецепторами. В организме человека лигандами являются, например, нейротрансмиттеры, которые выделяются в синаптических щелях нервными клетками в ответ на нервный импульс, а также вещества, секретируемые другими клетками в окружающую их среду.
В первом случае сигнал воспринимается нервной клеткой и по нервному волокну приходит точно по адресу к другой клетке. Этот тип регуляции быстрый и обеспечивается нервной системой.
Во втором случае, называемом гуморальной регуляцией, сигнальное вещество может действовать на целую группу клеток. Если оно действует в ближайшем окружении от выделившей его клетки, говорят о локальных химических медиаторах (от лат. localis – местный, medius – посредничающий). Один из примеров такого медиатора – белок гистамин, который выделяют так называемые тучные клетки в ответ на повреждение окружающих их тканей. В результате действия гистамина увеличивается просвет близлежащих кровеносных сосудов, и к месту травмы устремляются отряды лимфоцитов, буквально протискивающихся через стенки капилляров. Поглощая сигнальные вещества, которые сами же и выделяют, клетки осуществляют самоконтроль и самонастройку на определенную работу.
Возможна также и гуморальная регуляция состояния всего организма, когда сигнальное вещество синтезируется определенным типом ткани, попадает в кровь и разносится с кровотоком по всему телу. Такой тип сигнальной коммуникации обеспечивают гормоны. Однако и на гормоны реагируют только клетки, имеющие соответствующие рецепторы.
Химические вещества, способные связываться с наружными клеточными рецепторами и влиять на функционирование клетки, называют первичными медиаторами, или первичными мессенджерами (англ. messenger – посыльный). Межклеточная сигнализация удивительным образом напоминает основы коммуникации, разработанные в человеческом обществе.
Провода телефонной сети похожи на хитросплетение нервных волокон. Сообщение по ним проходит от одного абонента к другому. Роль локальных химических медиаторов играют устные сообщения. Они поступают только к ближайшим слушателям. Без специальных технических ухищрений до всего общества они не дойдут.
Самонастройку и самоконтроль осуществляет каждый человек, отдавая устный или письменный приказ самому себе. Кровеносное русло выполняет функции почтовой связи, которая помимо именных писем и бандеролей ежедневно разносит тысячи рекламных листочков, которые опускаются в каждый абонентный ящик. Реагируют же на эти сообщения о починке телевизоров или продаже сахара мешками только люди, которые имеют потребность в данной услуге – «рецептирующие» информацию о ней.
Загадка цАМФ
Каким же образом первичные мессенджеры оказывают влияние на активность белков и, тем самым, на активность клетки? Для этого внешний по отношению к клетке сигнал должен превратиться во внутренний. Ключевую роль в таком процессе играют образующиеся внутри клетки вторичные мессенджеры, которых на удивление мало. Главенствующую роль среди них играет циклический аденозинмонофосфат (цАМФ), открытый в 1958 г. Э.Сазерлендом и Т.Роллом.
Это соединение образуется из знаменитой АТФ – аденозинтрифосфорной кислоты, которую часто называют «энергетической разменной монетой» клетки. Как известно, АТФ состоит из азотистого основания аденина, пятиуглеродного циклического сахара и трех остатков фосфорной кислоты (рис. 1). Химические связи между фосфорными остатками богаты энергией. Практически все внутриклеточные процессы, начиная от синтеза белков и кончая мышечным сокращением, получают энергию за счет отщепления одной или двух фосфатных групп АТФ.
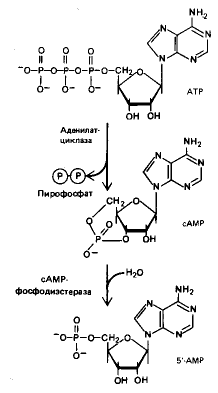
Рис. 1. Синтез и расщепление цАМФ
Циклический аденозинмонофосфат образуется с помощью фермента аденилатциклазы, который отщепляет от АТФ два остатка фосфорной кислоты, а последний, третий, остаток замыкает через два атома кислорода на сахар рибозу, входящий в состав аденозина (рис. 1). Это удивительное вещество играет роль универсального вторичного мессенджера в клетках практически всех организмов, как эукариотических, так и прокариотических (рис. 2).
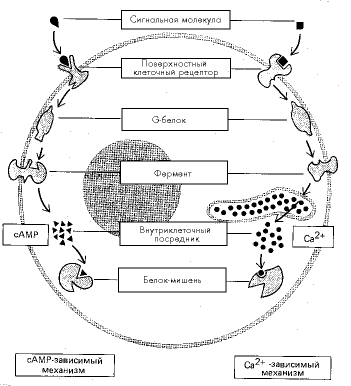
Рис. 2. Главные механизмы образования внутриклеточных мессенджеров
В качестве примера рассмотрим, как цАМФ влияет на образование гликогена, который является формой хранения глюкозы в животных клетках.
В момент опасности из надпочечников в кровь выбрасывается большое количество адреналина. Двигаясь по кровеносной системе, этот гормон достигает мышечных клеток, имеющих рецепторы адреналина. Связывание адреналина приводит к изменению пространственной структуры рецептора, что, в свою очередь, активирует фермент аденилатциклазу, расположенную на внутренней поверхности клеточной мембраны. Аденилатциклаза начинает превращать АТФ в цАМФ, и внутриклеточная концентрация последнего быстро возрастает. При достижении определенного уровня цАМФ активирует фермент протеинкиназу, который присоединяет остатки фосфорной кислоты (фосфорилирует) к аминокислотам серину и треонину в ферменте гликогенсинтетазе. Как следует из названия, этот фермент занимается в клетках синтезом гликогена. Фосфорилирование изменяет пространственную структуру фермента, в результате чего он инактивируется и новые порции гликогена уже не образуются. Протеинкиназа, активированная цАМФ, фосфорилирует также еще один фермент – киназу фосфорилазы. Киназа, в свою очередь, фосфорилирует гликогенфосфорилазу, которая в результате начинает отщеплять от гликогена молекулы глюкозы (рис. 3). Появившаяся же в мышцах глюкоза играет роль топлива, на котором они успешно работают.
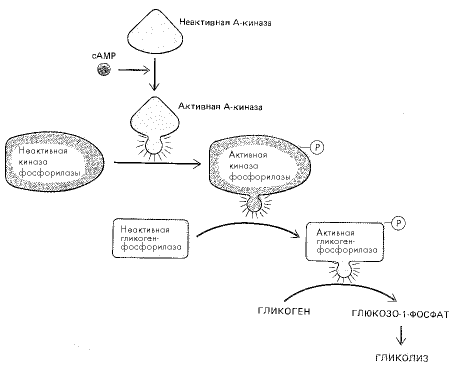
Рис. 3. Схема стимулирования распада гликогена повышением уровня цАМФ
Цитоскелет сигнализирует
Регулируемая цАМФ каскадная схема взаимодействий ферментов кажется непростой, а в действительности устроена еще более сложно. В частности, связавшиеся с первичными мессенджерами рецепторы оказывают влияние на активность аденилатциклазы не непосредственно, а через так называемые G-белки (рис. 4), работающие под контролем гуанинтрифосфорной кислоты (ГТФ).
А что происходит, когда почему-либо нарушается нормальная связь событий? Примером может быть заболевание холерой. Токсин холерного вибриона оказывает воздействие на уровень ГТФ и влияет на активность G-белков. В результате уровень цАМФ в клетках кишечника больных холерой оказывается постоянно высоким, что вызывает переход больших количеств ионов натрия и воды из клеток в просвет кишечника. Следствие этого – изнуряющие поносы и потеря воды организмом.
В норме под воздействием фермента фосфодиэстеразы цАМФ в клетке быстро инактивируется, превращаясь в нециклический аденозинмонофосфат АМФ. Течение другого заболевания – коклюша, вызываемого бактериями Bordetella pertussis, сопровождается образованием токсина, который тормозит превращение цАМФ в АМФ. Отсюда возникают и неприятные симптомы болезни – покраснение горла и доходящий до рвоты кашель.
На активность фосфодиэстеразы, превращающей цАМФ в АМФ, влияют, например, кофеин и теофиллин, что обуславливает стимулирующее действие кофе и чая.
Многообразие эффектов цАМФ и способов регуляции его концентрации в клетках делает его универсальным вторичным мессенджером, играющим ключевую роль в активации различных протеинкиназ.
В разных клетках цАМФ может приводить к совершенно различным эффектам. Это соединение не только принимает участие в распаде гликогена и жиров, но также увеличивает частоту сердечных сокращений, влияет на расслабление мускулатуры, контролирует интенсивность секреции и скорость поглощения жидкостей. Оно является вторичным мессенджером для целого спектра различных гормонов: адреналина, вазопрессина, глюкагона, серотонина, простогландина, тироид-стимулирующего гормона; цАМФ работает в клетках скелетной мускулатуры, сердечной мышцы, в гладких мышцах, почках, печени, в тромбоцитах.
Резонно возникает вопрос: почему же различные клетки реагируют на цАМФ по разному? Можно сформулировать его и иначе: почему при повышении концентрации цАМФ в разных клетках активируются различные протеинкиназы, которые фосфорилируют разные белки? Эту ситуацию можно проиллюстрировать с помощью такой аналогии. Представьте себе, что к двери офиса постоянно подходят различные визитеры – лиганды и первичные мессенджеры. При этом они звонят в один-единственный звонок: раздается сигнал – вторичный мессенджер. Как при этом служащим заведения определить, кто именно пожаловал с визитом и как надо реагировать на данного посетителя?
Загадка ионов кальция
Рассмотрим сначала, что происходит со вторым чрезвычайно распространенным вторичным мессенджером – кальцием, вернее его ионами. Впервые их ключевая роль в ряде биологических реакций была показана еще в 1883 г. когда Сидней Рингер заметил, что изолированные мышцы лягушки не сокращаются в дистиллированной воде. Чтобы в ответ на электрическую стимуляцию мышца сократилась, ей необходимо присутствие в окружающей ее среде ионов кальция.
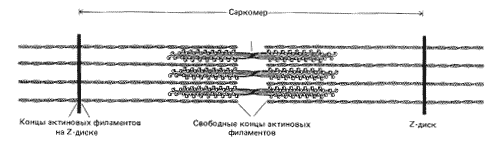
Рис. 5. Схема строения скелетной мышцы (саркомера)
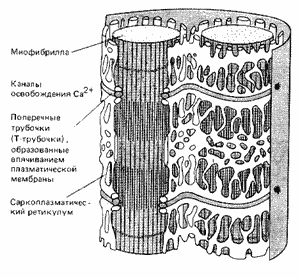
Рис. 6. Система мембран, передающая сигнал от плазмалеммы мышечной клетки ко всем миофибриллам
Теперь последовательность основных событий, происходящих при сокращении скелетной мускулатуры, хорошо известна (рис. 5). В ответ на электрический импульс, который доходит до мышцы по аксону нервной клетки, внутри мышечной клетки – миофибриллы – открываются резервуары ионов кальция – мембранные цистерны, в которых концентрация ионов кальция может быть выше, чем в цитоплазме, в тысячу и более раз (рис. 6). Высвободившийся кальций соединяется с белком тропонином С, который связан с выстилающими внутреннюю поверхность клетки актиновыми филаментами. Тропонин (рис. 7) играет роль блокатора, препятствующего скольжению миозиновых нитей по актиновым филаментам. В результате присоединения кальция к тропонину блок отсоединяется от нити, миозин скользит по актину, и мышца сокращается (рис. 8). Как только акт сокращения заканчивается, специальные белки – кальциевые АТФазы – закачивают ионы кальция обратно во внутриклеточные резервуары.
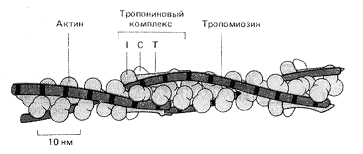
Рис. 7. Схема расположения на актиновом филаменте тропонина и тропомиозина
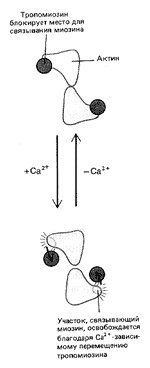
Рис. 8. Актиновый филамент в поперечном разрезе
На концентрацию внутриклеточного кальция оказывают влияние не только нервные импульсы, но и другие сигналы. Например, это может быть уже знакомый нам цАМФ. В ответ на появление адреналина в крови и соответствующее повышение концентрации цАМФ в клетках сердечной мышцы в них высвобождаются ионы кальция, что приводит к учащению сердцебиения.
Вещества, оказывающие влияние на кальций, могут содержаться также непосредственно в клеточной мембране. Как известно, мембрана состоит из фосфолипидов, среди которых один – фосфоинозитол-4, 5-дифосфат – играет особую роль. Помимо инозита молекула фосфоинозитол-4, 5-дифосфата содержит две длинные углеводородные цепи, состоящие из 20 и 17 атомов углерода (рис. 9). Под воздействием определенных внеклеточных сигналов и под контролем уже знакомых читателям G-белков они отсоединяются, в результате чего образуются две молекулы – диацилглицерин и инозитолтрифосфат. Последний участвует в высвобождении внутриклеточного кальция (рис. 10). Такого рода сигнализация используется, например, в оплодотворенной икре шпорцевой лягушки.
Проникновение первого же из множества спермиев в готовую для оплодотворения икринку вызывает образование в ее мембране инозитолтрифосфата. В результате ионы кальция высвобождаются из внутренних резервуаров и оболочка оплодотворенной яйцеклетки мгновенно разбухает, отсекая путь внутрь яйцеклетки менее удачливым или менее расторопным сперматозоидам.
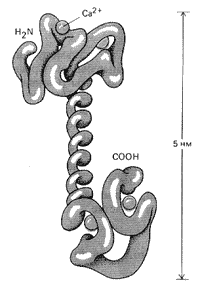
Рис. 11. Структура молекулы кальмодулина
Как же такое простое вещество, как ион кальция, может регулировать активность белков? Выяснилось, что он связывается внутри клетки со специальным белком кальмодулином (рис. 11). Этот достаточно крупный белок, состоящий из 148 аминокислотных остатков, как и цАМФ, обнаружен практически во всех изученных клетках.
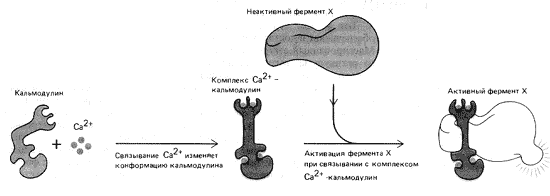
Рис. 12. Механизм активации Ca2+-зависимого ферментау
Присоединяющийся к кальмодулину кальций активирует его аналогично тому, как цАМФ стимулирует работу протеинкиназ. Именно так, например, происходит инициация сокращения гладкой мускулатуры. Высвободившиеся в ответ на внешний сигнал ионы кальция связываются с кальмодулином, который после этого взаимодействует с ферментом киназой и активирует ее (рис. 12). Комплекс киназа–кальмодулин связывается с актином, приводя его в рабочее состояние. В результате гладкие мышцы сокращаются. Опосредованный кальцием путь сигнала к поперечно-полосатой скелетной мускулатуре более длителен, зато гладкие мышцы в отличие от поперечно-полосатых могут значительно дольше находиться в сокращенном состоянии. Именно поэтому мускулы-замыкатели двустворчатых раковин могут часами сжимать свои створки.
В клетках разных тканей активированный кальцием кальмодулин связывается с различными белками-мишенями, влияя на их работу. Такое поведение кальмодулина вызывает вопрос, который возникал и при обсуждении влияния цАМФ и активируемой им протеинкиназы на активность белков, а именно: почему в разных клетках одни и те же активированные кальцием молекулы кальмодулина присоединяются к различным белкам?
Загадка стероидных гормонов
Совершенно аналогичная проблема возникает и при изучении гидрофобных стероидных гормонов, структура которых похожа на структуру жирорастворимого вещества холестерина. Термин «гидрофобный» указывает на их плохую растворимость в воде (от гр. hydor – вода и phobos – страх). Такие гормоны, будучи жирорастворимыми, легко проходят через состоящие из фосфолипидов клеточные мембраны. Оказавшись внутри клетки, стероидные гормоны связываются с соответствующими рецепторами. Рецепторы изменяют свою пространственную форму (конформацию) и, проникая в ядро через его ядерную мембрану, соединяются с определенными последовательностями нуклеотидов в ДНК, тем самым «включая» или «выключая» транскрипцию определенных генов. Такая последовательность событий доказана для стероидного гормона кортизона – его комплекс с рецептором связывается с соответствующим единственным геном, который удалось выделить и клонировать. Какая цепь превращений вызывается действием других гормонов и как происходит их специфическая «посадка» на определенный участок ДНК, во многом пока не ясно.
Не ясно и другое. Показано, что один и тот же гормон, связываясь со своим специфическим рецептором, вызывает различные ответы в разных клетках.
Забытое пространство
Число разнообразных сигналов, которые клетка может получать извне, очень велико. Это слабые электрические импульсы, гормоны, медиаторы, различные ростовые факторы и другие воздействия. Количество же вторичных мессенджеров, с помощью которых все это множество внешних сигналов влияет на внутриклеточные процессы, удивительно мало. Это цАМФ, ионы кальция, специальные молекулы типа высокофосфорилированных нуклеотидов (РРАРР – фосфат-фосфат-аденин-фосфат-фосфат) или инозитолтрифосфат.
Как же с помощью этих вторичных посредников клетка умудряется понять, какие именно сигналы их вызвали и каким образом необходимо на них реагировать? На этот вопрос трудно ответить прежде всего потому, что все клеточные компоненты (молекулы и ансамбли молекул) строго определенным образом скомпонованы в пространстве клетки.
Изучая по отдельности детали сложного часового механизма, который устроен несравнимо проще клетки, нелегко понять, как все эти шестеренки, маховички и пружины влияют друг на друга в работающем хронометре. Между тем именно такую задачу приходится решать исследователям клетки. Чтобы понять существо отдельных явлений или процессов, нужно разрушить клетку, выделить из нее белки, изучить их свойства и только потом попытаться установить их роль в том или ином процессе. При этом допускаются упрощения. Так, обычно принимается, что водорастворимые белки свободно диффундируют в цитоплазме наподобие крупинок в супе и никак не связаны между собой. Между тем само устройство некоторых клеточных органелл предполагает, что комплексы взаимодействующих ферментов должны образовывать специально сконструированные архитектурные ансамбли. Например, множество белков, катализирующих окислительные реакции в дыхательной цепи, располагаются на внутренних мембранах митохондрий в строго определенном порядке. Именно такая пространственная организация позволяет им с успехом осуществлять передачу богатых энергией электронов.
Но клетка содержит множество белков, связи которых друг с другом более лабильны и изменчивы во времени. По-видимому, для регуляции их пространственного взаимодействия требуется не прочное «заякоривание», а более тонкий и гибкий механизм. В частности, обеспечивать такую пространственную организацию могут белки цитоскелета. Они образуют настолько ажурные и динамичные структуры, что их изучение стало возможным лишь относительно недавно.
Строительные леса цитоскелета
Первые свидетельства сложности внутренней архитектуры цитоплазмы были получены еще в XIX в., когда в результате серебрения срезов тканей в клетках стали различать явственно проступающие сетеподобные структуры. Однако к изучению их состава и устройства удалось приступить лишь в 60-е гг. XX в., когда в биологии стали широко применяться такие тонкие методы исследований, как электронная микроскопия, ультрацентрифугирование и электрофорез.
В цитоплазме были обнаружены сложные структуры, образующие цитосклет. Выяснилось, что тяжи цитоскелета построены в основном из тонких (диаметром 7 нм) актиновых филаментов и длинных, толстых (диаметром 25 нм) и жестких микротрубочек, состоящих из - и -тубулина. Эти белки оказались очень лабильными, способными формировать легко изменяющиеся динамичные пространственные структуры. В частности, глобулярные белки актина не только легко и быстро полимеризуются в длинные вытянутые нити – филаменты (рис. 13). Они взаимодействуют с целым набором других вспомогательных белков, в результате чего возникает определенным образом организованная пространственная сеть филаментов.
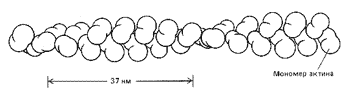
Рис. 13. Организация глобулярных молекул актина в актиновом филаменте
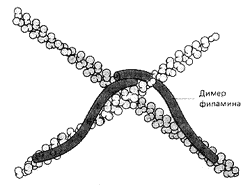
Рис. 14. Создание сети с помощью молекул филамина
Вспомогательные белки филамин и -актинин выполняют функции своеобразных скобок, сшивающих филаменты актина в структуру, напоминающую рыболовную сеть (рис. 14). Белок фибрин связывает актиновые филаменты в толстый пучок вроде веника или снопа. Тропомиозин стабилизирует уже сформированные тяжи актина (рис. 7). Гельзолин действует словно секатор, разрезая длинные филаменты на отдельные кусочки. Профилин, как нянька, сопровождает актиновые глобулы к местам их присоединения к образовавшимся ранее фрагментам нитей, виллин служит инициатором полимеризации актина в растворе, а тимозин, наоборот, не позволяет глобулярному актину соединятся в нити. Наконец, миозин способен активно подтягивать филаменты навстречу друг другу (рис. 5). Таким образом актиновые филаменты можно уподобить арматуре строительных лесов, которые можно резать, надставлять и соединять под любыми углами и в любых положениях. Их можно также соединять вместе, создавая тянущее напряжение всей конструкции.
Тубулин оказался белком не менее замечательным, чем актин. Образованные им микротрубочки способны разбираться с одного конца и собираться с другого. Белок нексин помогает им объединяться в сложные ансамбли, а белок динеин может скользить по тубулиновой микротрубочке, как дрезина по рельсам. Поскольку динеин в то же время способен жестко прикрепляться к микротрубочке другой своей стороной, это обеспечивает взаимное скольжение трубочек друг относительно друга.
В некоторых случаях филаменты и микротрубочки образуют четкий шаблон для построения определенных органоидов. Так происходит, например, в развивающемся сперматозоиде: актиновые филаменты формируют спиралевидную конструкцию, структуру которой потом, после ее распада, в точности повторяют возникшие на этом месте митохондрии. Высокоорганизованные летательные мышцы насекомых строятся в эмбриогенезе по тубулиновым шаблонам.
Помимо актиновых филаментов и тубулиновых микротрубочек в состав цитоскелета входят также промежуточные филаменты (диаметром 7–11 нм), которые исследованы значительно хуже, но, как полагают, являются не менее лабильными образованиями.
Белки цитоскелета принимают деятельное участие в движении клетки, поскольку для его осуществления требуется постоянное изменение ее формы. Мышечное сокращение, амебоидное движение, перешнуровывание клетки во время деления, фагоцитоз основаны на взаимодействии актина и миозина, а биение ресничек и жгутиков сперматозоидов происходит благодаря скольжению микротрубочек друг относительно друга.
Белки цитоскелета незаменимы там, где надо создать сложную пространственную и относительно стабильную форму. Например, внутри микроворсинок эпителиальных клеток кишечника и почек проходят пучки актиновых филаментов. Принципиально такие же, но гораздо более мощные пучки находятся в стереоцили волосковых клеток в улитке внутреннего уха. Похожая на двояковогнутую шайбу форма эритроцита поддерживается благодаря взаимодействию актина с белками спектрином и анкирином.
Подобные примеры можно было бы множить, однако важнее задаться вопросом: только ли к фиксации формы клетки сводится роль ее цитоскелета? Может быть, он играет существенную роль в образовании функциональных комплексов водорастворимых ферментов, а образованная цитоскелетом сеть служит для приема и передачи информации? Исходя из физико-химических свойств белков цитоскелета, это, в принципе, возможно. Вспомните ловчую сеть пауков. Она не только образует хитрые ловушки для насекомых. Натяжение и дрожание паутинок сигнализируют их конструктору о пойманной добыче или непредвиденной поломке ажурной конструкции.
Паутина сигнализирует
Указания на роль цитоскелета в передаче информации в клетки начали накапливаться с 1980-х гг. К этому времени уже было известно явление так называемого кеппинга.
Вернемся к началу этой статьи. Когда сигнальные вещества – лиганды – взаимодействуют со своими рецепторами, образовавшиеся комплексы собираются на поверхности клетки в компактную группу. Затем клеточная мембрана изгибается и комплексы лигандов с рецепторами втягиваются внутрь клетки (интернализируются), где происходит их утилизация. В этом процессе принимает участие актин, филаменты которого связываются с внутриклеточной частью пронизывающего мембрану рецептора. Играет ли при этом актин роль только лишь стягивающей сеточки, необходимой для образования на мембране впадины, или же его роль в этом процессе более сложна, остается пока не выясненным.
Актиновые филаменты способны прикрепляться не только к рецепторам, но и к клеточной мембране в районе так называемых фокальных контактов, образующихся в местах соприкосновения клетки с субстратом. Являются ли эти контакты лишь местами крепления к субстрату или же они одновременно информируют клетку об окружающих ее молекулах, частицах?
Проведенная выше аналогия устройства цитоскелета с сетью паутины становится почти наглядной, если учесть, что микротрубочки и промежуточные филаменты тянутся от ядра к периферии клетки. Из исследований культивируемых вне организма клеток хорошо известно, что большинство из них при осуществлении активной работы распластываются на той или иной подложке (рис. 15). В этих условиях в клетках формируется сложная трехмерная сеть филаментов.
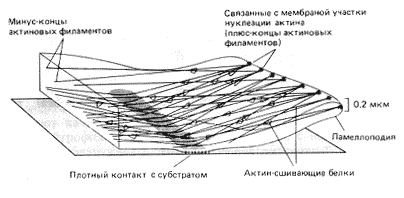
Рис. 15. Схема расположения актиновых филаментов в тонких пластинчатых отростках (ламеллоподиях) клеток, растущих в культуре
Эти наблюдения хорошо согласуются с данными о том, что митохондрии и лизосомы передвигаются в клетке не случайным образом, а вдоль микрофиламентов. Часть белоксинтезирующего аппарата клетки тоже связана с цитоскелетом. Если разрушить микротрубочки, то расположение таких важных органоидов как пузырьковидные элементы аппарата Гольджи, в которых проходят конечные стадии созревания готовых для экскреции белков нарушается, они оказываются размещенными в клетке случайным образом, а не в определенном порядке. Некоторые водорастворимые ферменты, участвующие в гликолизе, связаны с актиновыми филаментами. Хорошо известно, что в транспорте белков в нервных клетках также участвуют актиновые филаменты.
Следовательно, для синтеза определенных белков (а значит, и для выполнения определенных функций), клетка должна привести свой цитоскелет в рабочее состояние, которое обеспечивает необходимую пространственную организацию клеточных реакций и процессов. С этим выводом хорошо согласуется тот факт, что при различных стрессовых воздействиях клетка в первую очередь разбирает основные компоненты своего цитоскелета, а затем формирует их заново, в соответствии с реакцией на полученный сигнал. Такая перестройка обеспечивает переключение с одного режима работы на другой.
Будет ли клетка по-разному формировать свой молекулярный скелет в ответ на активацию различных поверхностных рецепторов? Опыты с фибробластами и эпителиальными клетками, распластывающимися на стекле, покрытом различными белками, дают на этот вопрос однозначный ответ.
Если на стекло нанести белок внеклеточного матрикса – фибронектин, то распластавшиеся на нем фибробласты принимают полигональную форму и в них активно формируются состоящие из актина так называемые стрессфибриллы. Другой нанесенный на стекло белок внеклеточного матрикса – ламинин – вызывает активное движение фибробластов вследствие образования у них узких спицеподобных микрошипов и плоских тонких ламеллоподий (рис. 15). Стрессфибриллы в этой ситуации не образуются вовсе. Наконец, нанесенные на стекло антитела к эпидермальному фактору роста (веществу, стимулирующему активное деление клеток кожи) вызывают в распластывающихся клетках эпидермиса образование полусфер из актина.
Хотя детали этих процессов остаются пока не ясными, очевидно, что клетки по-разному формируют свой цитоскелет в зависимости от тех или иных сигналов, полученных из окружающей среды.
Скептически настроенный читатель может возразить на это, что реорганизация цитоскелета не имеет прямого отношения к проведению внутриклеточных сигналов, а скорее является следствием, реакцией клетки на эти сигналы. Однако такое утверждение, в свою очередь требует доказательств.
Интересные, в этом плане, результаты были получены при работе с клетками CHO (от англ. Chinese Hamster Ovary cells), выделенными из китайских хомячков. При определенных условиях они становились раковыми. Вернуть их в нормальное состояние помогало добавление бутирата цАМФ – формы цАМФ, которая легко проникает внутрь клеток. Если же предварительно раковые клетки CHO обрабатывались разрушающим цитоскелет цитохалазином, то нормализации не происходило. Из этих экспериментов следует, что для успешного использования цАМФ в качестве вторичного мессенджера клетка должна иметь работоспособный цитоскелет.
Очень важны данные исследователей, работавших с протоонкогенами Ras, Rho и Cdc42 (т.е. с генами, повреждение которых почти неизбежно превращает нормальную клетку в раковую). Ген Ras активируется (т.е. начинается синтез закодированного в нем белка) уже упоминавшимся эпидермальным фактором роста, Rho – мощным стимулятором клеточного деления лизофосфатидиловой кислотой, а Cdc42 – пептидным гормоном брадикинином. В экспериментах было показано, что действие гена Ras связано с образованием в клетках сети микрофиламентов и ламелл, похожих на амебные псевдоподии. Ген Rho ответственен за формирование стрессфибрилл, а Cdc42 вызывает образование филлоподий.
Полученные результаты становятся совсем уже интригующими в свете последних открытий, связанных с изучением особых трансмембранных белков интегринов. Как правило, эти клеточные рецепторы состоят из двух цепей – альфа и бета, причем существует 15 вариантов первой и 98 вариантов второй. Собираясь в разных комбинациях, они образуют как минимум 20 различных типов интегринов, некоторые из которых способны связываться с белками внеклеточного матрикса, например, ламинином и фибронектином. При этом происходит активация уже знакомых нам протоонкогенов Ras и Cdc42!
Каким же образом полученный интегринами сигнал поступает в ядро? Сами интегрины не обладают фосфатазной активностью, т.е. не способны активировать белки с помощью присоединения к ним фосфатных групп. Зато торчащую внутрь клетки часть молекулы интегрина окружает целый комплекс белков, таких, например, как таллин, винкулин, паксиллин, а также фермент FAC-киназа, связанный с фокальными контактами – своеобразными «пуантами», на которых клетка передвигается по субстрату. От этого белкового ансамбля внутрь клетки отходят актиновые нити, которые совместно с микротрубочками и промежуточными филаментами могут доходить от мембранной периферии клетки до ее ядра. Полагают, что так передается сигнал в ядро клетки.
Это предположение подтверждается данными экспериментов, проведенных с искусственно выращиваемыми мышиными фибробластами, трансформированными онкогеном Ras. В этих опытах было показано, что в ответ на внешний сигнал – эпидермальный фактор роста – клетка синтезирует белок – фактор транскрипции NFkb, который участвует в самой первой стадии синтеза белка – считывании (транскрипции) РНК с ДНК, находящейся в ядре. Фактор транскрипции находится в цитоплазме и связан непосредственно с элементами цитоскелета, а именно со стрессфибриллами и фокальными контактами. Это означает, что соединение, играющее существенную роль в регуляции работы ядерных генов, буквально вплетено в цитоскелетную сеть!
Возвращаясь к загадкам вторичных мессенджеров, можно предположить, что цитоскелет принимает активное участие в проведении внутриклеточных сигналов, за счет образования пространственных комплексов между рецепторами, соответствующими протеинкиназами и активируемыми ими белками. Аналогичную роль цитоскелет может играть и в ситуации с кальмодулином, поскольку определенное его количество в клетке связано с пучками актиновых филаментов и промежуточных филаментов. Функции цитоскелета в активации ядерных генов менее ясны, но указания на такую возможность уже получены.
Кто в клетке король?
На этом можно было бы и закончить рассказ о возможной роли цитоскелета во внутриклеточной сигнализации, если бы не еще одно любопытное соображение. Дело в том, что в ядре клетки содержится информация о первичной структуре всех белков, включая белки цитоскелета, но нет абсолютно никаких указаний на их взаимное расположение. Если образованные белками пространственные структуры чрезвычайно стабильны, можно говорить о самосборке, которая действительно происходит, например, в случае образования белковых оболочек вирусов или при формировании прямо в пробирке тубулиновых микротрубочек. Однако когда дело касается таких лабильных и способных к сложным перестройкам структур, как цитоскелет, только к самосборке их образование не свести. Что же определяет ту или иную конфигурацию цитоскелета?
Явно не ядро – как не определяет состояние общества накопленная в библиотеках информация. Ее можно почерпнуть оттуда, но что и когда будет востребовано определяется самим обществом. Вероятно, так же действует и клетка, достаточно автономно от ядра формируя свою пространственную структуру.
Известно, что миниатюрные безъядерные фрагменты фибробластов живут часами, активно ползая и приобретая различные формы. Живущие неделями в кровяном русле тромбоциты также лишены ядра, что не мешает им принимать активное участие в таком важном процессе, как свертывание крови при травмах сосудов. Пространственная структура клетки может самовоспроизводиться и даже оказывать воздействие на ее внешние реакции. Интегрины, например, обладают уникальной для трансмембранных (т.е. пронизывающих мембрану насквозь) рецепторов способностью реагировать на внутренние клеточные сигналы, изменяя при этом свое сродство к тем или иным внешним лигандам. Известно также, что именно цитоскелет определяет ориентацию белков внеклеточного матрикса и тем самым оказывает влияние на соседние клетки.
Таким образом, ответ на вопрос о том, что же есть жизнь, во многом зависит от понимания того, каким образом длительно существуют во времени и самовоспроизводятся сложные пространственные ансамбли макромолекул, прекрасным примером которых является цитоскелет
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://bio.1september.ru
Похожие работы
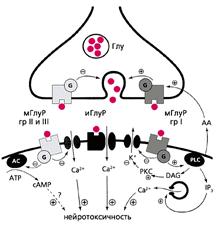
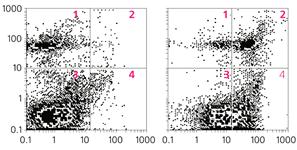
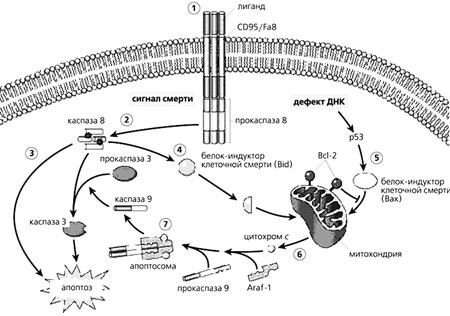
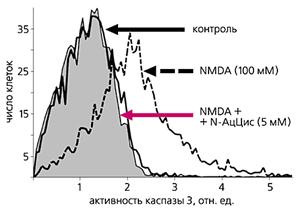
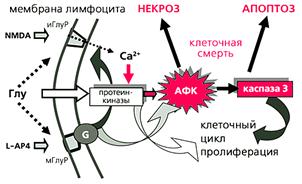
... многих стадиях клеточного цикла и в процессах пролиферации [4]. Особенно важны эти реакции для клеток иммунной системы. Значит, в ряде случаев активность каспазы 3 не обязательно означает начало апоптоза, а может быть связана с пролиферацией лимфоцитов. Глутаматные рецепторы иммуннокомпетентных клеток История открытия и изучения глутаматных рецепторов накопила массу примеров их причастности к ...
0 комментариев