Министерство образования РФ
Муниципальная средняя школа № 33
Экзаменационный реферат
по биологии на тему
«Ферменты»
Выполнил:
ученик 10 Г класса
Елизаров Александр
Научный руководитель
Захаров С. П.
Смоленск 2000
Содержание:
1. Введение.
2. Ферменты.
· История открытия.
· Природа ферментов.
а) Структуры.
б) Специфичность.
3. Состав.
4. Классификация.
5. Номенклатура.
6. Активность ферментов.
7. Механизм действия.
8. Значение.
· В организме.
· В науке.
9. Заключение.
ВВЕДЕНИЕ.
«Ферменты ( от латинского слова fermentum – закваска) – белки, которые обладают каталитической активностью и характеризуются очень высокой специфичностью и эффективностью действия. Все процессы в живом организме- дыхание, пищеварение, мышечное сокращение, фотосинтез и другие – осуществляются с помощью ферментов. Ферменты находятся во всех живых клетках и составляют большую часть всех их белков. Они во много миллионов раз ускоряют самые разнообразные химические превращения, из которых складывается обмен веществ. Под действием различных ферментов составные компоненты пищи: белки, жиры и углеводы – расщепляются до более простых соединений, из которых затем в организме синтезируются новые макромолекулы, свойственные данному типу. » Вот, всё что я знал о ферментах. Я решил пополнить свои знания и поэтому взял реферат по ферментам.
ИСТОРИЯ ОТКРЫТИЯ.
Науку составляет не только достигнутый результат, но и путь ведущий к результату путь от незнания к знанию, медленный, извилистый, скачкообразный, в каждой области зависящий от достижений смежных наук и общего развития мировоззрения. Ещё в незапамятные времена, на заре возникновения цивилизации, люди в своей практической деятельности сталкивались с различными ферментативными процессами и использовали их для своих целей. Это
спиртовое и молочнокислое брожение, применение сычуга для приготовления сыров, солода и плесневых грибов- для осахаривания продуктов. Вероятно, первым, кто попытался
создать общее представление о химических процессах в живом организме, был врач и ученый Парацельс, родившийся в Швейцарии в конце XV века. Несмотря на наивность
(с совершенной точки зрения), взгляды Парацельса во многом были прогрессивными, так как для понимания жизненных явлений он пытался привлечь реальные силы природы. Именно с этих позиций Парацельс и его последователи подошли к рассмотрению сущности ферментации, давно известного понятия обозначавшего разного рода брожения, главным
образом спиртовое и молочнокислое. В XVI и начале XVII века уже делались попытки рассматривать ферментации как химические процессы. И Василий Валентин ( первая
половина XVI века), и Андрей Либавий (1550-1616 годы) считали ферменты ( или дрожжи) особым веществом, хотя и подчиняли его действия неким не материальным силам. Другим последователем Парацельса был знаменитый голландский химик Иоганн Баптиста Ван Гельмонт (577-1644 годы). Именно он охарактеризовал фермент как агент, вызывающий химические процессы в организме и управляющий ими. Качественный скачёк в развитии учения о ферментациях произошёл в связи с исследованиями великого французского химика Антуана Лавуазье, совершившего переворот в химии и впервые внедрившего в химические исследования строгие количественные методы. К концу XVIII века уже было известно, что встречаются химические процессы, протекающие с участием какого-то агента, без которого процесс практически не идёт.
Первые успехи были достигнуты при изучении превращения крахмала в сахар. Решающая роль в этих исследованиях принадлежит работам петербургского академика К. С Кирхгофа, которые открыли новую страницу в истории и химия ферментов. В начале XIX века было открыто немало химических реакций, среди них были и некоторые ферментативные реакции. Юстус Либих был одним из наиболее крупных авторитетов среди химиков XIX века. В это время было открыто ещё несколько ферментов. В 1836 году Т.Шванн впервые обнаружил в желудочном соке фермент животного происхождения, названный им пепсином. Несколько позже, в 1857 году, А.Корвизар описал другой фермент, переваривающий белки - трипсин. В XIX веке (1897 год) Эдуард Бухнер убедительно доказал химическую природу ферментов. В 1907 году - Эдуард Бухнер был удостоен Нобелевской премии по химии. (В.И.Розенгарт Ферменты- двигатели жизни).
ПРИРОДА ФЕРМЕНТОВ.
а) После того как стало возможным исследование ферментов в бес клеточной среде, была окончательно установлена их химическая природа. Было выявлено, что все они представляют собой вещества белковой природы и, как все белки могут быть простыми и сложными в зависимости от сопутствующего компонента небелкового характера ( простетической группы).
Так мы подчёркивали, что свойство каждого белка определяется последовательностью расположения остатков аминокислот в их молекуле. Эта последовательность называется первичной структурой белка. В последние годы разработаны очень надёжные, и даже автоматизированные методы изучения первичной структуры, что дало возможность определить
полную аминокислотную последовательность для многих белков, в том числе и для ферментов. Помимо первичной структуры, определяемой последовательностью расположения аминокислот, для проявления специфических свойств белка (в ном числе ферментативной активности) важную роль играют более высокие уровни - вторичная и третичная структуры, сущность которых заключается в определённом расположение полипептидных цепей в пространстве.
Вторичная и третичная структуры белков поддерживаются сравнительно слабыми внутримолекулярными связями, и поэтому легко могут быть разрушены разными физическими и химическими воздействиями. Такое нарушение высших структур белка без повреждения его первичной структуры составляет сущность денатурации. При денатурации белок нередко утрачивает свои биологические свойства, в случае ферментов исчезает ферментативная активность. Современные методы исследования позволяют получить представление не только о первичной структуре белков. Есть ферменты, для которых полностью выяснено пространственное расположение атомов, составляющее их молекулу, то- есть расшифрованы вторичная и третичная структуры. Это достигнуто благодаря применению исключительно тонкого и сложного метода, так называемого рентгеноструктурного анализа. Некоторым белкам свойственен ещё более высокий уровень структуры - четвертичная структура. Это уже надмолекулярный уровень: функционирование такого белка нуждается не в одной, а в нескольких молекулах ( чаще всего в двух или четырёх), которые вместе образуют комплекс, обладающий всеми специфическими свойствами. Каждая отдельная молекула такого белка, составляющая четвертичный комплекс, называется субъединицей. Многие ферменты построены из субъединиц. В одних случаях субъединиц сами обладают активностью, в других их субъединиц по отдельности неактивны. Субъединицы, сопоставляющие молекулу фермента, могут быть одинаковыми, но могут и отличатся друг от друга. Представление о молекуле фермента как структуре, состоящей из субъединиц , позволяет нам объяснить одно очень интересное и практически важное явление. Существуют ферменты, различающиеся по строению, но катализирующие одну и ту же реакцию, они называются изоферментами. Такие ферменты довольно широко распространены в организме, и их выявление имеет большое значение в медицине.
б) Одно из наиболее поразительных свойств ферментов их специфичность. Специфичность ферментов проявляется по- разному и может быть выражена в разной степени. Прежде всего следует различать специфичность по отношению к субстрату и к типу химической реакции, катализируемой ферментом.
Специфичность по отношению к реакции.
Каждый фермент катализирует одну химическую реакцию или группу реакций одного типа. Наиболее ярким проявлением этого вида специфичности могут служить довольно частые случаи, когда одно и то же химическое соединение выступает как субстрат действия нескольких ферментов, причём каждый из них, катализирует специфическую для него реакцию, приводит к образованию совершенно различных продуктов (смотри приложение № 1).
· В первой реакции под действием фермента оксидазы происходит окисление аминокислот. При этом аминогруппа (NH2) отделяется в форме аммиака (NH3) и образуется соединение, содержащие кретонную группу (С=О) и называемое кетокислотой.
· Вторую реакцию катализирует декарбоксилаза. Под влиянием этого фермента из карбоксильной группы (- СООН) отщепляется углекислота (СО2) и остаётся амин.
· Третья реакция более сложна. Она катализируется ферментом трансиминазой и состоит в переносе аминогруппы с аминокислоты на кетонокислоту. Мы видим. что исходная аминокислота имеет радикал R, а образовавшаяся в результате реакции новая аминокислота- радикал R'.
Итак, один и тот же субстрат подвергается разным превращениям под влиянием различных ферментов.
Специфичность по отношению к субстрату.
Наряду с только, что описанной формой специфичности фермента по отношению к катализируемой им реакции существует и другая, тесно связанная с первой форма специфичности, выражающаяся в способности фермента атаковать субстрат только определённого химического строения. Иногда фермент способен действовать только на один единственный субстрат, тогда говорят, что он обладает абсолютной специфичностью. Значительно чаще фермент влияет на группу субстратов, имеющих сходное строение. Такую специфичность называют групповой. Особый интерес представляет так называемая стереохимическая специфичность, состоящая в том, что фермент действует на субстрат или группу субстратов, отличающихся особым расположением атомов в пространстве.
Абсолютная специфичность встречается редко.
Хорошим примером фермента , обладающего очень высокой, практически абсолютной специфичностью может служить уреаза, катализирующая гидролиз мочевины.
H2N\
C=O + H2O = CO2 + 2NH3
H2N/ вода углекислота аммиак
мочевина
Долгое время считалось, что мочевина является единственным субстратом уреазы. Но не так давно было показано, что кристаллическая уреаза может действовать и на близкого родственника мочевины - оксимочевину, отличающуюся наличием в молекуле одного атома кислорода.
HOHN\
C=O
H2N/
оксимочевина
«Правда, реакция гидролиза мочевины под влиянием уреазы протекает в 120 раз медленнее, чем гидролиз мочевины» (В. И. Розенгарт Ферменты- двигатели жизни)
Таким образом, понятие "абсолютная специфичность" является в известной мере относительным.
Групповая специфичность. Она характеризует подавляющее большинство ферментов и состоит в том, что фермент, проявляя свойственную ему специфичность по отношению к реакции, способен действовать не на один, а на несколько, иногда на большое число субстратов со сходным химическим строением. Например (смотри приложение № 1), три разных фермента, действующие на аминокислоты. все они обладают групповой специфичностью, так как действуют не на какую-нибудь одну аминокислоту, а на многие, иногда на все аминокислоты.
· Относительно групповая специфичность проявляется тогда, когда фермент безразличен к структуре соединения и имеет значение лишь тип связи. Примером служит химотрипсин, расщепляющий только пептидную связь.
Стереохимическая и оптическая специфичность имеет особое значение. Проявляется только в случае оптически активных веществ, и фермент активен только по отношению к одной стереоизомерной форме соединения. Например, L- аргиназа разлагает L-аргинин на L- орнитин и мочевину, но не действует на А- аргинин. Известным примером служит d и L- специфичность оксидаз аминокислот. Стереохимическая и оптическая активность так- же может быть абсолютной и относительной; например, карбоксипептидаза, расщепляющая карбобензокси -глицил-L- фенилаланин совсем не действует на субстрат с А- фенилаланином: с другой стороны, эстераза свиной печени разлагает метиловый эфир L- миндальной кислоты лишь вдвое быстрее, чем его А- изомер.
СОСТАВ.
После того как стало возможным исследование ферментов в бес клеточной среде, была окончательно установлена их химическая природа. Было выявлено, что все они представляют собой вещества белковой природы и как все белки, могут быть простыми и сложными в зависимости от сопутствующего компонента небелкового характера (простетической группы).
Ферменты- простые белки- построены только из аминокислот, и их каталитические свойства обусловлены свойством самой белковой молекулы. К этой группе ферментов относится большинство гидролитических ферментов. Ферменты- сложные белки- содержат в своём составе, помимо белкового компонента, ещё и небелковый- например, нуклеотиды, геминовую группу, витамины, атомы ( катионы ) металла. К таким ферментам обычно относятся ферменты окислительно-восстановительного действия. Прочность связи между белковым компонентом и простетической группой в сложных ферментах может быть различной. В некоторых случаях связь прочная, в других - простетическая группа довольно легко отделяется, например при диализе. Легко диссоциирующие простетические группы ферментов получили название коферментов. При отделении простетической группы от белковой части фермента - последний теряет свою активность. В простых ферментах активный центр образуется непосредственно группировкой аминокислотных остатков в спиральной цепи белковой молекулы. В сложных ферментах он образуется простетической группой и некоторыми прилегающими к ней остатками. Размер активных центров значительно меньше самой молекулы фермента. На один активный центр приходится масса молекулы с молекулярным весом 30000. В простых ферментах пространственная группировка этих аминокислотных остатков сама по себе определяет структуру активного центра и каталитическую активность фермента. В сложных ферментах структура активного центра определяется простетической группой и боковыми группами некоторых аминокислотных остатков, пространственная структура которых оказывает существенное влияние на специфичность и каталитическую активность небелкового компонента. Среди таких аминокислотных остатков наибольшее значение имеют SH- группы цистеина, OH- группы серина, несколько меньшее значение имеет индольная группа триптофана, карбонильные группы дикарбоновых аминокислот. Компоненты активного центра нельзя представлять последовательно расположенными на, каком - либо участке цепи. По- видимому , активный центр формируется из компонентов, удалённых в первичной структуре полипептидной цепи, но пространственно сближенных благодаря специфической укладке полипептидной цепи.
КЛАССИФИКАЦИЯ.
Сейчас известно около 2 тысяч ферментов, но список этот не закончен. В зависимости от типа катализируемой реакции все ферменты подразделяются на 6 классов:
· Ферменты, катализирующие окислительно-восстановительные реакции оксидоредуктазы;
· Ферменты переноса различных группировок ( метильных, амино- и фосфогрупп и другие)- трансферазы.
· Ферменты, осущевствляющие гидролиз химических связей - гидролазы
· Ферменты не гидролитического отщепления от субстрата различных группировок (NH3, CO2,H2O и другие)- лиазы.
· Ферменты, ускоряющие синтез связей в биологических молекулах при участии доноторов энергии, например АТФ,- лигазы.
· Ферменты, катализирующие превращение изомеров друг в друга,- изомеразы.
ОКСИДОРЕДУКТАЗЫ – ферменты, катализирующие окислительно-восстановительные процессы в организме. Они осущевствляют перенос водорода и электронов и по своим привиальным названием известны как дегидрогеназы, оксидазы и пероксидазы. Эти ферменты отличаются тем, что имеют специфические коферменты и простетические группы. Их подразделяют на функциональные группы доноров, от которых они принимают водород или электроны, и акцепторов, на которые они их передают (на СН-ОН группу, СН- NH группу, C-NH группу и другие).
ТРАНСФЕРАЗЫ – ферменты, переносящие атомные группы ( в зависимости от того, перенос какой группы они осуществляют, их соответственно называют). Среди них известны ферменты осуществляющие транспорт больших остатков, например гликозилтрансферазы и другие. Трансферазы благодаря разнообразию переносимых ими остатков принимают участие в промежуточном обмене веществ.
ГИДРОЛАЗЫ – ферменты, катализирующие гидролитическое расщепление различных субстратов (при участии молекул воды). В зависимости от этого среди них различают эстеразы, расщипляющие сложноэфирную связь между карбоновыми кислотами (липаза) тиоловых эфиров, фосфоэфирную связь и так далее; гликозидазы, расщепляющие гликозидные связи, пептид - гидролазы, действует на пептидную связь и другие.
ЛИАЗЫ. К этой группе относятся ферменты, способные отщеплять различные группы от субстрата не гидролитическим путём с образованием двойных связей или, напротив, присоединять группы к двойной связи. При расщеплении образуется Н2О или СО2 или большие остатки- например ацетил- СоА. Лиазы играют весьма важную роль в процессе обмена веществ.
ИЗОМЕРАЗЫ – ферменты, катализирующие превращение изомерных форм друг в друга, то - есть осуществляющие внутримолекулярное превращение различных групп. К ним относятся не только ферменты, стимулирующие реакции взаимных переходов оптических и геометрических изомеров, но и такие, которые могут способствовать превращению альдоз в кетозы или перемещению эфирной связи и другие.
ЛИГАЗЫ. Раньше эти ферменты не отделяли от лиаз, так как реакция последних часто идёт в двух направлениях, однако недавно было выяснено, что синтез и распад в большинстве случаев происходит под влиянием различных ферментов, и на этом основании выделен отдельный класс лигаз (синтетаз). Ферменты, обладающие двойным действием, получили название бифункциональных. Лигазы принимают участие в реакции соединения двух молекул, то есть синтетических процессах, сопровождающихся расщеплением макроэнергитических связей АТФ или других макроэргов.
«Первое подразделение ферментов на самые крупные группы (6 классов) основано не на названии субстрата, а на природе химической реакции, которую фермент катализирует. Далее, внутри классов ферменты делят на подклассы, руководствуясь строением субстрата. В подклассы объединяют ферменты данного класса, действующие на сходно построенные субстраты. На этом деление не заканчивается. Ферменты каждого подкласса разбивают на подклассы, в которых ещё строже уточняют структуру химических групп, отличающих субстраты друг от друга. Подкласс это последняя низшая ступень классификации. Внутри подклассов перечисляют уже отдельные, индивидуальные ферменты. Таким образом, вся система проста и достаточно стройна:
КЛАСС- ПОДКЛАСС- ПОДПОДКЛАСС- ИНДИВИДУАЛЬНЫЙ ФЕРМЕНТ.
В соответствии с этим принципом классификации предложена очень удобная система нумерации (индексации) ферментов. Каждый индекс состоит из четырёх цифр, разделённых точками:
1. Номер класса.
2. Номер подкласса в данном классе
3. Номер подподкласса
4. Номер, присвоенный данному индивидуальному ферменту этого подподкласса» (В. И. Розенгарт Ферменты- двигатели жизни)
Например, амилаза-фермент, гидролизующий крахмал с которой мы уже встречались неоднократно, имеет индекс 3.2.1.1. Классификация ферментов построена так, что в ней оставлены свободные места для ещё не открытых ферментов.
НОМЕНКЛАТУРА.
Ферментология очень долго не располагала, строг научной номенклатурой ферментов. Наименования ферментам давали по случайным признакам (тривиальная номенклатура), по названию субстрата (рациональная), по химическому составу фермента, наконец, по типу катализируемой реакции и характеру субстрата. Примерами тривиальной номенклатуры могут служить названия таких ферментов, как пепсин (от греч. пепсин - пищеварение), трипсин (от греч. трипсис - разжижаю) и папаин (от названия дынного дерева Carica papaja, из сока которого он выделен). По действию все эти ферменты являются протеолитическими, т. е. ускоряют гидролиз протеинов (белков). Характерное название была дано группе окрашенных внутриклеточных ферментов, ускоряющих окислительно-восстановительные реакции в клетке, - цитохромы (от лат. citos - клетка и chroma - цвет).
Наибольшее распространение получила рациональная номенклатура, согласно которой название фермента составляется из названия субстрата характерного окончания -аза. Она была предложена более столетия тому назад, в 1883 г. Э. Дюкло - учеником Л. Пастера. Так, фермент, ускоряющий реакцию гидролиза крахмала, получил название амилаза (от греч. амилон - крахмал), гидролиза жиров - липаза (от греч. липос - жир), белков (протеинов) - протеаза, мочевины - уреаза (от греч. уреа - мочевина) и т. п. Когда методами аналитической химии были достигнуты известные успехи в расшифровке химической природы простетических групп, возникла новая номенклатура ферментов. Их стали именовать по названию простетической группы, например, геминфермент (простетическая группа - гем), пиридоксаль- фермент (простетическая группа - пиридоксаль) и т.п. Затем в названии фермента стали указывать как на характер субстрата, так и на тип катализируемой реакции. К примеру, фермент, отнимающий водород от молекулы янтарной кислоты, называют сукцинатдегидрогеназой, подчеркивая этим одновременно и химическую природу субстрата, и отнятие атомов водорода в процессе ферментативного действия:
- 2Н
![]() НООС -СH2 - СН2 – CООН НООС - СН = СН – СООН
НООС -СH2 - СН2 – CООН НООС - СН = СН – СООН
Янтарная кислота Дегидрирование Малеиновая кислота
В 1961 г. Международная комиссия по номенклатуре ферментов представила V Международному биологическому конгрессу проект номенклатуры, построенный на строго научных принципах. Проект был утвержден конгрессом, и новая номенклатура прочно вошла в ферментологию. Согласно этой (Московской) номенклатуре название ферментов составляют из химического названия субстрата и названия той реакции, которая осуществляется ферментом . Если химическая реакция, ускоряемая ферментом, сопровождается переносом группировки атомов от субстрата к акцептору, название фермента включает также химическое наименование акцептора. Например, пиридоксальфермент, катализирующий реакцию переаминирования между L-аланином и -кетоглутаровой кислотой, называется L-аланин: 2-оксоглутарат аминотрансфераза. В этом названии отмечены сразу три особенности: 1) субстратом является L-аланин; 2) акцептором служит 2-окcоглутаровая кислота; З) от субстрата к акцептору передается аминогруппа.Названия ферментов по научной номенклатуре неизмеримо выигрывают в точности, но становятся в ряде случаев гораздо сложнее старых, тривиальных. Так, уреаза (тривиальное название), ускоряющая реакцию гидролиза - мочевины на оксид углерода (IV) и аммиак, по научной номенклатуре именуется карбамид - амидогидролазой:
![]() Н2N - СО - NН2 + Н2О 2NН3 + СО2
Н2N - СО - NН2 + Н2О 2NН3 + СО2
В этом названии дано точное химическое наименование субстрата и указано, что фермент катализирует реакцию гидролиза аминогруппы. Трегалаза, ускоряющая реакцию гидролиза трегалозы, называется трегалоза-1-глюко-гидролазой.. В связи со значительным усложнением научных названий в новой номенклатуре допускается сохранение наряду с новыми старых тривиальных, рабочих названий ферментов. Международной комиссией был составлен детальный список всех известных в то время ферментов, существенно дополненный в 1972 г. при пересмотре, как классификации, так и номенклатуры некоторых ферментов, где рядом с новым научным названием каждого фермента приведено старое, а также указан химизм катализируемой ферментом реакции и в некоторых случаях природа фермента. Таким образом, исключается возможность путаницы в наименовании ферментов. В 1964 г. список включал 874 фермента; в последующее время он был существенно дополнен и возрос до 1770 ферментов в 1972 г. и до 2003 ферментов в 1979 г.
АКТИВНОСТЬ ФЕРМЕНТОВ.
Для исследования или практического работника, занимающегося ферментами, определение активности ферментов - это постоянная, повседневная работа, потому что любое изучение свойств ферментов, любое применение их в практической деятельности- в медицине и в народном хозяйстве- всегда связано с необходимостью знать, с какой скоростью протекает ферментативная реакция. Что бы понять и правильно оценить результаты определения ферментативной активности, нужно совершенно отчётливо представить себе, от каких факторов зависит скорость реакции, какие условия оказывают на неё влияние. Таких условий много. Прежде всего это соотношение концентрации самих реагирующих веществ: фермента и субстрата. Далее, это всевозможные особенности той среды, в которой протекает реакция: температура, кислотность, наличие солей или других примесей, способных как ускорять, так и замедлять ферментативный процесс, и так далее. Попытаемся рассмотреть поближе эти условия.
ВЛИЯНИЕ РЕАКЦИЙ СРЕДЫ.
Для большинства известных в настоящее время ферментов определён оптимум РН, при котором они обладают максимальной активностью. Эта величина- важный критерий, служащий для характеристик фермента. Иногда это свойство ферментов используют для их препаративного разделения. Наличие оптимума РН можно объяснить тем. Что ферменты представляют собой полиэлектролиты и их заряд зависит от значения РН (Смотри приложение 2). Иногда сопутствующие вещества могут изменить оптимум РН, например буферные растворы. В некоторых случаях в зависимости от субстратов ферменты с неярко выраженной специфичностью имеют несколько оптимумов. Например, пепсин расщепляет белки яйца при РН 1,5- 2,0, синтетические субстраты- при РН 4,0. Отсюда следует, что величина (РН оптимум)- весьма чувствительный признак для данного фермента. Она зависит от природы субстрата, состава буферного раствора и поэтому не является истинной константой. Нужно иметь в виду также свойства ферментов как белковых тел, способных к кислотно-щелочной денатурации. Поэтому при определении оптимума РН, в котором сохраняется физико-химическая стабильность фермента. Кислотно-щелочная денатурация может привести к необратимым изменениям структуры фермента с утратой его каталитических свойств.
ВЛИЯНИЕ ДРУГИХ ХИМИЧЕСКИХ ФАКТОРОВ.
Присутствие в реакционной среде некоторых ионов может активировать образование активного субстрат ферментного комплекса, и в этом случае скорость ферментативной реакции будет увеличивается. Такие вещества получили название активаторов. При этом вещества, катализирующие ферментативные реакции, непосредственного участия в них не принимают. На активность одних ферментов существенно влияет концентрация солей в системе, другие ферменты не чувствительны к присутствию ионов. Однако некоторые ионы абсолютно необходимы для нормального функционирования некоторых ферментов. Известны ионы, которые тормозят активность одних ферментов и являются активаторами для других. К числу специфических активаторов относятся катионы металлов: Na+, K+,Rb+,Cs+,Mg2+, Ca2+,Zn2+,Cd2+,Cr2+,Cu2+,
Mn2+,Co2+,Ni2+,Al3+. Известно также, что катионы
Fe2+,Rb+,Cs+ только в присутствии Mg действуют как активаторы, в других случаях эти катионы не являются активаторами. В большинстве случаев один или два иона могут активировать тот или иной фермент. « Например, Mg2+- обычный активатор для многих ферментов, действующий на фосфоримированные субстраты, почти во всех случаях может быть заменён Mn2+, хотя другие металлы его заменить не могут. Следует заметить, что щелочноземельные металлы вообще конкурируют друг с другом, в частности, Са2+ подавляет активность многих ферментов, активируемых Mg2+ и Zn2+. Причина этого до настоящего времени не ясна» (Г. А. Смирнова Основы биологии). Механизм влияния ионов металлов- активаторов может быть различным. Прежде всего, металл может быть компонентом активного центра фермента. Но может действовать как связующий мостик между ферментом и субстратом удерживая субстрат у активного центра фермента. Имеются данные о том, что ионы металлов способны связывать органическое соединение с белками и, наконец, один из возможных механизмов действия металлов как активаторов- это изменение константы равновесия ферментативной реакции. Доказано, что анионы также влияют на активность ряда ферментов. Например, очень велико влияние СI- на активность А - амилазы животного происхождения. Наряду с
существованием активаторов ферментов известен ряд веществ, присутствие которых тормозит каталитическое действие ферментов или полностью инактивирует его. Такие вещества принято называть ингибиторами. Ингибиторы – это вещества, действующие определённым химическим путём на ферменты и по характеру своего действия, могут быть подразделены на обратимые и необратимые ингибиторы. Для обратимого торможения Характерно равновесие между ферментом и ингибитором с определённой константой равновесия. Система такого типа характеризуется определённой степенью торможения, зависящей от концентрации ингибитора, при этом торможение достигается быстро и после этого не зависит от времени. При удалении ингибитора с помощью диализа активность фермента восстанавливается. Необратимое торможение, прежде всего, выражается в том, что диализ не способствует восстановлению активности фермента. И в отличии от обратимого торможения усиливается со временем, так что может наступить полное торможение каталитической активности фермента при очень низкой концентрации ингибитора. В этом случае эффективность действия ингибитора зависит не от константы равновесия, а от константы скорости, определяющей долю фермента, подвергшегося торможению в данном случае.
ВЛИЯНИЕ ТЕМПЕРАТУРЫ. Температура – один из важнейших факторов внешней среды, который независимо от состояния равновесия реакции меняет её скорость. Поэтому при ферментативных реакциях при повышении температуры на 10 С процесс ускоряется в 1,5 – 2 раза. При дальнейшем повышении температуры присоединяются денатурационные процессы, характерные для всех белков и в то м числе для ферментов, поэтому наблюдается затухание скорости реакции (Смотри приложение 3). Температурным оптимумом реакции называют температуру, при которой одно её действие вызывает ускорение реакции, катализируемой данным ферментом. Для большинства ферментов животного происхождения он равен 40 – 50 С, для растительного происхождения он равен 50 – 60 С. Почти все ферменты разрушаются при температуре 80 С. Но для некоторых ферментов в настоящее время доказана возможность восстановления их каталитической активности в случае обратимого процесса денатурации белка. Известны и такие ферменты, максимальная активность которых проявляется при более низких температурах. «Например, каталаза, температурный оптимум которой лежит в пределах между 0-10С» (Г. А. Смирнова Основы биохимии). Понижение температуры снижает скорость ферментативных реакций. Большинство ферментов при 0 С ещё не утрачивают своих каталитических свойств, но при замораживании химические реакции прекращаются. При последующем оттаивании, если соблюдается определённые условия, ферментативная активность клеток может быть восстановлена.
ВЛИЯНИЕ ДАВЛЕНИЯ. При изучении действия давления на скорость ферментативных реакций необходимо, прежде всего, учитывать, как и при изучении других факторов, возможность денатурации ферментов при высоком давлении. Если константа скорости ферментативной реакции растёт с повышением давления, то образование активного комплекса происходит с уменьшением объёма и наоборот, если при увеличении давления образование активного комплекса сопровождается увеличением объёма, то константа скорости реакции снижается.
ВЛИЯНИЕ КОНЦЕНТРАЦИИ ФЕРМЕНТА И ЕГО СУБСТРАТА.
Скорость любого ферментативного процесса в значительной степени зависит от концентрации, как субстрата, так и фермента. Обычно скорость реакции прямо пропорциональна количеству фермента, при условии если содержание субстрата в в пределах оптимума или немного выше. При постоянном количестве фермента скорость возрастает с увеличением концентрации субстрата. Эта реакция подчинена закону действующих масс и рассматривается в свете теории Михаэлиса – Ментона, то есть
V=K(F) V- скорость реакции
K- константа скорости
F- концентрация фермента(Смотри приложение 4).
На графике показано соотношение скорости реакции и концентрации субстрата. В восходящей части гиперболы при низких концентрациях субстрата скорость реакции пропорциональна концентрации субстрата. В верхней части, когда концентрация субстрата высока, скорость реакции приближается к максимальному значению и почти не зависит от концентрации. Первое объяснение этой кривой было дано Генри (1901 год). Он высказал предположение, что а основе этой реакции лежит образование субстрат - ферментного комплекса. В дальнейшем эта теория была экспериментально обоснована Михаэлисом – Ментеном и не утратила своего значения до настоящего времени.
МЕХАНИЗМ ДЕЙСТВИЯ.
Предполагалось, что ферменты адсорбируют на своей поверхности реагирующие молекулы, в результате чего на участках сорбции концентрация молекул субстрата увеличивается, и это повышает вероятность протекания реакции между ними. Постепенно сложилось мнение, что фермент не сорбирует субстрат на своей поверхности, а вступает с ним во взаимодействие, причём это взаимодействие на первом этапе состоит в образовании непрочного соединения- комплекса между ферментом и субстратом. С каждой молекулой фермента ( а точнее, с каждым его каталитическим центром) реагирует одна молекула субстрата, причём реакция носит необратимый характер. Если фермент обозначить буквой Е, а субстрат буквой S, то реакцию можно написать в виде уравнения:
Именно таким образом представляли себе протекание ферментативной реакции немецкие учёные Л. Михаэлис и его сотрудница М. Ментен, которые ещё в 1913 году развили общую теорию ферментативного действия, основанную на идее образования промежуточного фермент- субстратного комплекса как первой стадии реакций. Чаще всего распаду комплекса предшествует его химическое преобразование( активирование), которое составляет ещё одну промежуточную стадию и снова усложняет уравнение реакции:
![]()
![]()
![]()
![]() E+S ES ES* E+P
E+S ES ES* E+P
Здесь активный комплекс обозначен ES* (Смотри приложение 5)
Скорости протекания отдельных стадий ферментативного процесса неодинаковы. Одни идут быстрее, другие медленнее. Скорость всей реакции будет определяться скоростью самой медленной реакции. В ферментативном процессе скорости разных стадий тоже неодинаковы. Первый этап этого процесса - образование фермент- субстратного комплекса ES представляет собой, как мы уже говорили, обратимую реакцию и в обычных условиях протекает чрезвычайно быстро, по- видимому, значительно быстрее, чем последующие стадии. Поэтому общая суммарная скорость всего процесса определяется не этой реакцией. Но эта стадия наиболее ответственна, так как сама важность каталитического действия фермента зависит от того, образуется фермент- субстратный комплекс или нет. Все последующие этапы - это только преобразование возникшего комплекса. Как же представить себе образование такого комплекса? Какие условия должны быть соблюдены для того, чтобы он возник? Если снова обратиться к схеме (Смотри приложение 5) и присмотреться к причудливой форме молекулы фермента и субстрата, то заметили, что участок молекулы фермента, на который «садится» субстрат,. Своими очертаниями как бы повторяют форму субстрата. Это символизирует строгое пространственное и химическое соответствие, существующее между активным центром фермента и субстратом. Такое соответствие совершенно необходимо для того, чтобы комплекс мог образоваться. Ещё в конце прошлого века известный немецкий химик Эмиль Фишер высказал предположение, что фермент должен подходить к субстрату как ключ к замку. Это выражение стало крылатым и дожило до наших дней. Однако образ «ключ-замок» перестал удовлетворять учёных. Этот образ предполагает жёсткость, неизменность структуры, железную прочность фермента и субстрата. Такие свойства не типичны для гибких, подвижных молекул биологических веществ. Поэтому, главным образом благодаря работам американского биохимика Д. Кашленда, возникла другая теория, дополняющая и расширяющая представления Фишера. Согласно этой гипотезе, полное соответствие между молекулой субстрата и каталитическим центром фермента возникает лишь тогда, когда они встречаются с друг другом. Субстрат вызывает в молекуле фермента такое изменение расположения химических групп в пространстве, что ранее отсутствовавшее соответствие появляется и вместе с этим появляется возможность образовать фермент- субстратный комплекс. Его возникновение связано с гибкостью белковой молекулы, с подвижностью её структуры, но оно возможно, разумеется, только в том случае когда молекула субстрата имеет пригодные для этого свойства и форму. В приложении 5 изображена схема, поясняющая возникновение наведённого соответствия между ферментом и субстратом.
Только после контакта фермента с субстратом химические группировки активного центра (А, В, С) в результате изменения их пространственного расположения приходят в состояние строгого соответствия молекуле субстрата.
Нужно иметь также в виду, что молекула субстрата, хотя она, как правило, и значительно меньше молекулы фермента, тоже обладает некоторой подвижностью и при взаимодействии с ферментом эта подвижность может способствовать более полному соответствию.
Особенность ферментов состоит в том, что об их наличии мы можем судить только по их действию. Мы умеем измерять скорость ферментативных реакций, то есть количество субстрата, подвергшегося превращению в единицу времени, например в одну минуту или в один час. Разным ферментам свойственна далеко не одинаковая молекулярная активность. Некоторое представление о реальных величинах этой активности даёт таблица (Смотри приложение 7). Из таблицы видно, насколько различна молекулярная активность различных ферментов и каких огромных величин она может достигать в отдельных случаях. «Карбоангидраза, занимающая первое место в таблице и обладающая чудовищной молекулярной активностью (36 миллионов), является самым активным из всех известных ферментов. «(В. И. Розенгарт Ферменты – двигатели
жизни).
ЗНАЧЕНИЕ.
Постоянный обмен нуклеиновыми кислотами, составляет основную часть генетического материала клетки. В ходе обмена нуклеиновых кислот наряду с синтезом происходит и распад. Этот процесс катализирует большая группа ферментов, объединенных названием нуклеаз. Цепочка нуклеиновых кислот образованна фосфорной кислотой и углеводородом; азотистые основания служат боковыми группами. Поэтому разрушение нуклеиновых кислот – это разрыв связей между остатками фосфорной кислоты и углевода. Все нуклеазы могут быть разделены на две группы: экзонуклеазы и эндонуклеазы. Экзонуклеазы действуют с одного из концов полинуклеотидной цепи и на каждом этапе отсекает по одному нуклеотиду, постепенно укорачивая цепочку. В отличие от этого эндонуклеазы сразу во многих местах разрывают связи внутри молекулы нуклеиновых кислот и поэтому приводят к быстрой деградации молекулы. Весь комплекс ферментов обмена нуклеиновых кислот выполняет важную биологическую задачу: сохранение в целостности генетического материала клетки и репарации (исправления) тех повреждений структуры ДНК, которые могут возникнуть а результате радиоактивного или ультрафиолетового облучения и других вредных воздействий.
Известно, что все проявления жизнедеятельности связаны с затратой энергии. Эта энергия освобождается при химических превращениях в клетке тех веществ, которые в виде пищи поступают в наш организм. Задача пищеварения сводится к тому, чтобы превратить главные пищевые вещества: белки, углеводы и жиры, - в такие продукты, которые непосредственно смогут быть использованы во внутриклеточном обмене. Свой путь в организме пища начинает, попадая в рот, и уже на этом этапе она сталкивается с ферментами. В слюне содержится фермент амилазы, катализирующий разложения крахмала и превращение его в сахар. Разжёванная и смоченная слюной пища проглатывается и через пищевод попадает в желудок. Слизистая оболочка желудка вырабатывает желудочный сок. В желудочном соке есть соляная кислота , придающая желудочному содержимому кислую среду. Так же в желудочном соке имеется протеолитический (расщепляющий белки) фермент – пепсин. Он как раз лучше всего действует в кислой среде. Пепсин не расщепляет белки до конца, он только ''раскладывает'' крупную белковую молекулу на части, доступные для действия пищеварительных ферментов кишечника. Из желудка пищевая кашица поступает в двенадцатиперстную кишку, где на неё изливаются соки дву самых крупных желёз человеческого организма: печени и поджелудочной железы. Сок поджелудочной железы содержит большой набор ферментов, действующих на все важнейшие пищевые вещества. Ферменты: трипсин и химотрипсин (расщепляющие белки) расщепляют пептидные цепи в разных местах. Комбинированная атака протеолитических ферментов желудочного и поджелудочного соков приводят к распаду белков на мелкие пептиды, содержащие небольшое количество аминокислотных остатков. В поджелудочном соке содержится чрезвычайно активная амилаза, она практически полностью завершает расщепление крахмала, начатое слюной. В результате крахмал превращается в солодовый сахар – мальтозу – дисахарид, состоящий из двух остатков глюкозы. Третий главный компонент пищи – жиры тоже расщепляются под влиянием поджелудочного сока. Для этой цели там содержится специальный фермент – липаза. Простейшая и наиболее распространённая форма жиров – триглицериды. Под действием липазы молекула триглицерида присоединяет три молекулы воды и распадается на составляющие его глицерин и жирные кислоты. Но заключительную работу в области пищеварения совершает кишечный сок, вырабатываемый клетками слизистых оболочек тонких кишок. Он содержит много ферментов, заканчивающих процесс окончательного разложения пищевых веществ. Осколки белковых молекул распадаются на отдельные аминокислоты; мальтоза, образовавшаяся из крахмала, и другие сложные углеводы превращаются в простые углеводы – моносахариды – вроде глюкозы. На этом заканчивается процесс пищеварения.
Одна из защитных реакций – свёртывание крови, происходит с участием ферментов. Как же происходит свертывание крови? Кровь, как известно состоит из жидкой части – плазмы и так называемых ферменных элементов, которые в ней плавают. Это кровяные клетки: эритроциты (красные кровяные тельца) и тромбоциты (кровяные пластинки). Плазма представляет собой сложный раствор многих веществ, в том числе самых разнообразных белков. Из белков плазмы для нас сейчас особый интерес представляет один – фибриноген. Пока кровь течёт по кровеносным сосудам, с фибриногеном ничего не происходит. Но стоит поранить сосуд настолько, чтобы кровь вытекала из него, как фибриноген очень быстро превращается в другой белок – фибрин. Фибрин, в отличии от фибриногена, не растворяется в плазме. В виде тонких нитей, переплетённых в густую сетку, он выпадает в осадок. В этой сетке застревают кровяные клетки и образуется плотный сгусток – тромб, препятствующий дальнейшему кровотечению. Превращение фибриногена в фибрин – процесс ферментативный, катализируемый ферментом тромбином. Тромбин – протеолитический фермент, подобный трипсину и химотрипсину. Но это фермент очень специфичный. Он действует только на фибриноген, отщепляя от его молекулы два сравнительно небольших полипептида. Оставшаяся часть молекулы фибриногена перестраивается и превращается в нерастворимый фибрин.
(тромбин)
![]() Фибриноген 2 Полипептида + фибрин
Фибриноген 2 Полипептида + фибрин
Также ферменты играют важную роль во всех проявлениях жизни. Успехи учения о ферментах внесли весомый вклад в развитие всех направлений человеческой практики.
Ферменты нашли широкое применение в медицине. Это, прежде всего, изучение таких болезней причина, которых лежит в недостаточности тех или иных ферментов. Далее это использование определения активности ферментов в биологических жидкостях и тканях для диагностики различных заболеваний. И, наконец, это применение ферментов в качестве лекарственных средств. Генетически обусловленные нарушения. Время от времени в бесконечно длинных цепях ДНК, где записаны все инструкции по синтезу белков, вдруг появляются случайные замены: вместо одного нуклеотида становится другой. Такие замены называются мутациями. Чаще всего конкретные причины мутации неизвестны. А последствия их нередко бывают роковыми. Приведем такой пример. Люди отличаются друг от друга цветом кожи, волос и глаз. Причина этого – разные пигменты, меланины, синтезируемые из некоторых аминокислот под влиянием определённых ферментов. Если образование этих пигментов не происходит из - за отсутствия одного из участвующих в реакции ферментов, возникает альбинизм – отсутствие окраски. Люди альбиносы имеют очень белые волосы и светлые глаза. Альбиносы по здоровью не уступают людям с нормальной окраской. Гораздо более тяжёлым заболеванием, нередко приводящим к гибели новорождённых, является непереносимость простых углеводов – моносахаридов (галактозы и фруктозы ). Здесь речь идёт о невозможности нормального обмена веществ в клетках из- за отсутствия необходимых ферментов. Достаточно подробно изучены врождённые болезни, связанные с недостатком ферментов, катализирующих разложение гликогена. В результате нарушения этого процесса гликоген начинает накапливаться в тканях в избыточном количестве и препятствует нормальному течению обмена веществ. Такие болезни получили название гликогенозов. Болезни, связанные с отсутствием витаминов, называют авитаминозом. Но по существу они являются ферментозами. Давно известна и когда – то была широко распространена болезнь ''бери – бери ''(сейчас её называют полиневритом – множественное воспаление нервов, в некоторых слаборазвитых странах она и теперь встречается нередко). Причина её отсутствие в пище витамина В1. Этот витамин – тиамин – в соединении с фосфорной кислотой представляет собой небелковую часть фермента декарбоксилазы. Декарбоксилаза разрушает карбоксильную группу (- СООН) некоторых органических кислот, отщепляя от неё углекислоту (СО2). В отсутствии витамина В1 декарбоксилаза образоваться не может, реакция прекращается и в нервной ткани наступают нарушения, типичные для полиневрита: параличи конечностей, боли в мышцах, слабость, контрактуры. Тяжёлое заболевание – пеллагра – связано с отсутствием в пище витамина РР – никотиновой кислоты. Упомянем ещё об одном витамине. Он называется витамином В2, а по химической природе представляет собой довольно сложную циклическую структуру – рибофлавин. Авитаминоз В2 связан с тяжёлым поражением кожи лица и глаз. Причина недостаток фермента.
Ферменты также используются в диагностике. Определение активности ферментов в биологических жидкостях и тканях стало неотъемлемым средством лабораторной диагностики различных заболеваний. Для диагностических целей ферментативную активность определяют почти исключительно в крови, значительно реже в моче и лишь в отдельных случаях в тканях. Не все ткани в одинаковой мере синтезируют разные ферменты. Для печени, например, типична высокая активность одних ферментов, для почек или скелетных мышц – других. Это явление называют органоспецифичностью ферментов. Иногда органоспецифичность выражена очень чётко: фермент содержится только в каком – нибудь одном органе и отсутствует, а других. Таким образом, врач получает возможность по повышению активности некоторых ферментов в плазме выявить заболевание, связанные с нарушением функций совершенно определенных органов. В последнее время предпринимаются всё более успешные попытки использовать ферменты и для лечения некоторых болезней. Уже давно некоторые ферменты применяют для так называемой заместительной терапии – для возмещения дефицита ферментов, возникающего при некоторых заболеваниях. Особенно успешна такая терапия при нарушениях функций желудочно-кишечного тракта, связанных с недостаточной выработкой пищеварительных ферментов. С успехом применяют ферменты в тех случаях, когда лечение требует разрушить накопившиеся в большом количестве белковые образования, мешающие нормальному функционированию тканей. Это бывает при ожогах, гнойных ранах, гнойно-воспалительных заболеваниях лёгких, когда в бронхах скапливается густая масса, препятствующая прохождению воздуха. Наметился очень перспективный путь применения ферментов для рассасывания сгустков крови, образовавшихся внутри кровеносных сосудов. Такие сгустки называются тромбами, они закупоривают сосуд и нарушают кровообращение. Велико значение ферментов в пищевой промышленности и сельском хозяйстве. Сыроварение, виноделие, производство кисломолочных продуктов, пивоварение, производство колбасных продуктов, хлебопечение, производство животных жиров, чая, уксуса, лимонной кислоты – всё это и многое другое , здесь не перечисленное – технологические процессы пищевой промышленности, в которых главным действующим лицом являются ферменты. Одна из важнейших проблем пищевой промышленности – это развитие комплексной переработки сырья и отходов пищевой промышленности и повышение эффективности этой переработки. Ферментные препараты могут сказать здесь решающее слово. Серьёзной проблемой в консервной промышленности, переработки плодов и овощей является использование семян и косточек, главная трудность которого состоит в необходимости разрушать прочную оболочку косточек. И здесь реальную пользу могут принести препараты ферментов. С помощью ферментных препаратов удаётся уменьшить расход сырья растительного и животного происхождения, идущего на приготовления пищевых продуктов. Использование ферментов в сельском хозяйстве необычайно широко и разнообразно. В растениеводстве селекция многих сельскохозяйственных культур направлена на создание сортов, обогащённых определёнными ферментами. Это имеет значение и для скорости созревания культур, и для получения более высококачественной продукции, и для повышения устойчивости растений к изменению погодных условий, к болезням, к действию вредных насекомых. Специальный интерес представляет использования ферментов в кормопроизводстве. Агрономы заботятся о том, чтобы получить полноценный растительный корм, содержащий все существенные составные части, необходимые для обеспеченья потребностей животного организма. Вот здесь роль ферментов оказалась особенно значительной, как в пищевой промышленности. Ферментативные препараты для производства кормов получают из плесневых грибов и бактерий, но задачи здесь ставят иные. Для повышения усвояемости грубых кормов необходим фермент целлюлоза, гидролизирующий клетчатку и повышающий возможность её переваривания и усвоения, особенно у таких животных, как свиньи, которые переваривают клетчатку хуже, чем крупный рогатый скот.
Мы познакомились с некоторыми аспектами практического использования ферментов в медицине, пищевой промышленности и сельском хозяйстве.
ЗАКЛЮЧЕНИЕ.
В результате проделанной работы я узнал, что ферменты это белки, катализирующие определённые химические реакции, входящие в процессы обмена веществ, отличаются чрезвычайно высокой эффективностью и специфичностью своего действия. По своему составу ферменты разделяют на простые ферменты, состоящие только из молекул белка, и сложные ферменты, состоящие из белка и небелкового компонента (простетические группы, коферменты). Каталитическое действие ферментов определяется главным образом, частью молекулы - активным центром. Действие всех ферментов происходит через стадию образования промежуточного соединения с молекулой субстрата. Ферменты играют важную роль в организме, в науке, в хозяйственной деятельности человека. Открытие разнообразных наук позволяет шире использовать ферменты.
Литература:
''Энциклопедический словарь юного биолога''
(М. С. Гиляров)
''Биофизическая химия'' (А. Г. Пасынский)
''Ферменты-двигатели жизни'' (В. И. Розенгарт)
''Основы биохимии'' (Г. А. Смирнова)
cedar_back@mail.ru
Похожие работы
... межпредметных связей между биологией и химией.Следует использовать знания,полученные учениками при изучении неорганической химиии в курсе 8-9 классов (темы:”Кислород,оксиды,горение”, ”Водород”, ”Кислоты,соли,основания”, “Строение вещества”). Формирование понятие “фермент” в курсе “Общая биология” Дальнейшее знакомство с ферментами ученики продолжают в курсе “Общая биология”. Здесь происходит ...
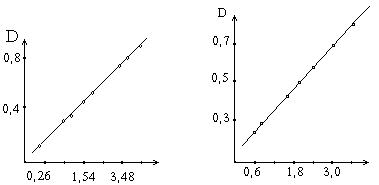
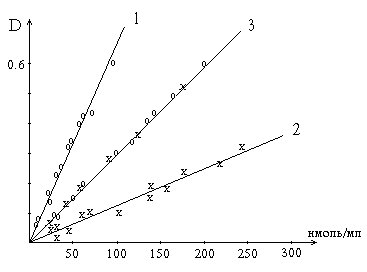
... (белка). Для целей определения ферментов могут быть использованы не только измерение поглощения света, но также измерения флюоресценции - спектрофлюорометрические методы. Такое определение активности фермента в ряде случаев по чувствительности превосходит спектрофотометрические методы на целый порядок величины. Некоторые коферменты и субстраты обладают сильной флюоресценцией. НАД и НАДФ в ...
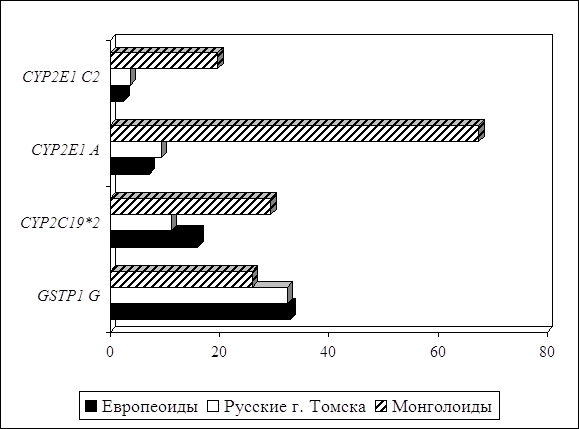
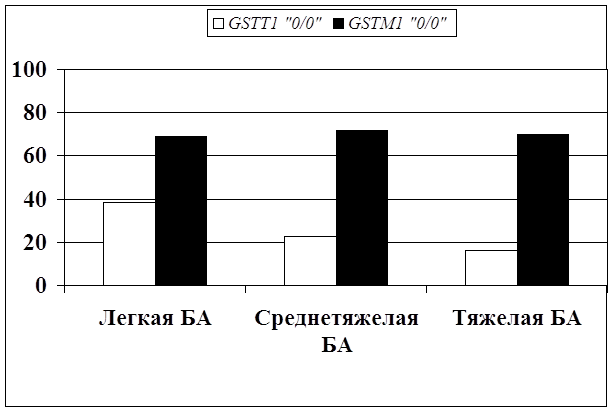
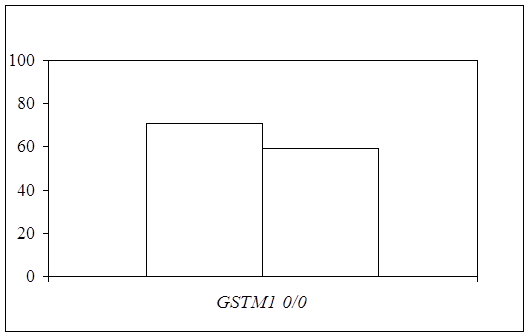
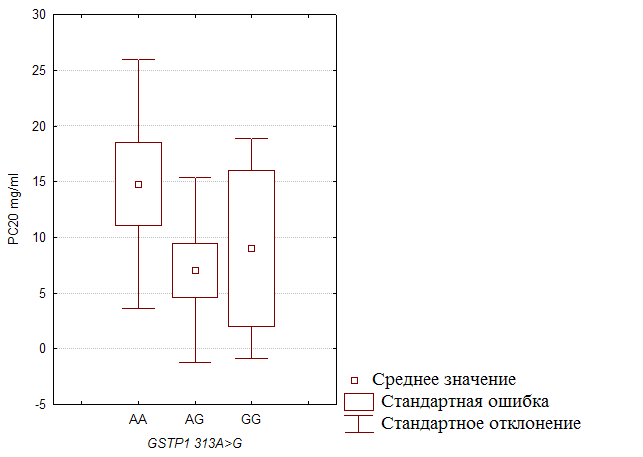
... препаратов. Установлена связь полиморфизма 313A>G гена GSTP1 с изменчивостью уровня аланинаминотрансферазы (р=0,021). 7. Выявлены различия в структуре генетической подверженности к бронхиальной астме и туберкулезу по генам ферментативной системы метаболизма ксенобиотиков: гены GSTM1, CYP2E1 и CYP2C19 связаны с бронхиальной астмой и значимыми для заболевания качественными и ...
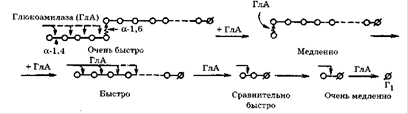

... амілази і пуллуланази або альфа-амілази і глюкоамілази, активних при високій температурі. Однак складність культивування анаеробних бактерій в заводських умовах робить цей спосіб отримання препарату амілолітичних ферментів практично непридатним. З літературних даних відомо, що найбільш термостабільними гідролітичними ферментами, такими як альфа-амілази, пуллуланази, а в ряді випадків і протеази є ...
0 комментариев