МИНИСТЕРСТВО ВЫСШЕГО И СРЕДНЕГО СПЕЦИАЛЬНОГО ОБРАЗОВАНИЯ РОССИИ
МОСКОВСКИЙ ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ ФИЗИКО-ТЕХНИЧЕСКИЙ ИНСТИТУТ
ДИПЛОМНАџЯ РАБОТА
АТФ индуцированное изменение внутриклетоЧной концентрации кальцияЯ в нейронах неокортекса крыс.
НАУЧНЫЙ РУКОВОДИТЕЛЬ академик Магура И.С.
КОНСУЛЬТАНТ к. б. н. Войтенко Н.В.
рецензент к.б.н. Исаев Д
ДИПЛОМНИК кругликов И.А.
Москва 1998
Оглавление
1. ÂÂÅÄÅÍÈÅ.......................................................................................................................... 4
2. ÎÁÇÎÐ ËÈÒÅÐÀÒÓÐÛ....................................................................................................... 5
2.1 Ãîìåîñòàç êàëüöèßÿ â íåðâíûõ êëåòêàõ............................................................... 5
2.1.1 Êàëüöèåâûå êàíàëû ïëàçìàòè÷åñêîé ìåìáðàíû.................................................... 5
2.1.2 Êàëüöèåâûå áóôåðû............................................................................................... 7
2.1.3 Êàëüöèåâûå êàíàëû ýíäîïëàçìàòè÷åñêîãî ðåòèêóëóìà...................................... 8
2.1.4 Êàëüöèåâûå íàñîñû............................................................................................... 11
2.1.5 Êàëüöèåâûå îáìåííèêè........................................................................................... 13
2.1.6 Ñà2+-ñâÿçûâàþùèå îðãàíåëëû............................................................................. 13
2.2 Вëèßÿíèå ÀÒÔ íà êàëüöèåâûé ãîìåîñòàç........................................................... 14
2.2.1 Ñòðîåíèå è ñâîéñòâà ÀÒÔ................................................................................... 15
2.2.2 Íîìåíêëàòóðà è ñóáêëàññèôèêàöèÿ ïóðèíîðåöåïòîðîâ...................................... 16
2.2.3 Ð1 ïóðèíîðåöåïòîðû............................................................................................... 16
2.2.4 Ð2 ïóðèíîðåöåïòîðû............................................................................................... 17
2.2.5 Ðåêëàññèôèêàöèÿ ïóðèíîðåöåïòîðîâ.................................................................. 20
3. ÎÁÚÅÊÒ È ÌÅÒÎÄÛ ÈÑÑËÅÄÎÂÀÍÈÉ........................................................................... 21
3.1 Ïîäãîòîâêà ïðåïàðàòà............................................................................................... 21
3.2 Õàðàêòåðèñòèêè êàëüöèåâîãî çîíäà.................................................................. 22
3.3 Îêðàñêà ñðåçîâ ôëóîðåñöåíòíûì êðàñèòåëåì................................................... 24
3.4 Ñòðóêòóðà ýêñïåðèìåíòàëüíîé óñòàíîâêè äëÿß äâóõ-âîëíîâîãî èçìåðåíèÿ êîíöåíòðàöèè êàëüöèÿß............................................................................................................................ 24
3.5 Ðàñòâîðû è ñìåíà ðàñòâîðîâ.................................................................................. 27
4. ÐÅÇÓËÜÒÀÒÛ ÈÑÑËÅÄÎÂÀÍÈÉ................................................................................. 28
5. ÎÁÑÓÆÄÅÍÈÅ................................................................................................................... 39
6. ÂÛÂÎÄÛ............................................................................................................................. 41
7. ÑÏÈÑÎÊ ËÈÒÅÐÀÒÓÐÛ.................................................................................................. 42
1. ВВЕДЕНИЕ
Молекула АТФ давно известна как повсеместно распространенный источник энергии для внутриклеточного метаболизма. Но ее свойства как нейротрансмитера были обнаружены сравнительно недавно. Сегодня уже не осталось никаких сомнений, что АТФ является нейротрансмитером в автономных нейромышечных соединениях, ганглиях и центральной нервной системе. К примеру, было показано, что АТФ вовлечена в генерацию болевых сигналов через Р2Х1 и Р2Х2 рецепторы. Однако роль и распределение пуринорецепторов в коре головного мозга и особенно в моторной коре до сих пор остается слабо изученной. Поэтому изучение механизмов действия АТФ в коре головного мозга представляет несомненный интерес. Мы изучали действие АТФ посредством измерения концентрации внутриклеточного кальция, - одного из найболее важных и универсальных регуляторов клеточных функций.
Цель работы состояла в изучении механизмов генерации АТФ - индуцированных внутриклеточных кальциевых сигналов в нервных клетках моторной коры.
2. ОБЗОР ЛИТЕРАТУРЫ 2.1 Гомеостаз кальция в нервных клетках
Концентрация цитозольного кальция в эукариотических клетках регулируется трансмембранным транспортом и цитоплазматическим связыванием кальция. Движение ионов кальция через мембрану контролируется: 1) двумя семействами Са2+ каналов, как то: кальциевыми каналами плазмалеммы и кальциевыми каналами, расположенными в мембране эндо(сарко)плазматического ретикулума (ЭР или СР), которые формируют пути входа кальция в цитоплазму; 2) выводом кальция из цитоплазмы благодаря активности кальциевых насосов плазмалеммы и/или кальциевых обменников; и 3) аккумуляцией ионов кальция внутриклеточными кальциевыми депо и митохондриями. Последние служат системами кальциевого буфера, способными аккумулировать и накапливать ионы кальция, поддерживая таким образом гомеостаз кальция в цитоплазме.
2.1.1 Кальциевые каналы плазматической мембраныНервные клетки экспрессируют различные типы кальциевых каналов плазмалеммы, которые могут быть активированы различными воздействиями. Основываясь на механизмы активации, кальциевые каналы могут быть разделены на несколько типов потенциал - управляемых и рецептор - управляемых каналов.
Потенциал - управляемые каналы вносят существенный вклад как в регуляцию входа кальция в цитоплазму, так и в нейрональный электрогенез. Белки, образующие кальциевые каналы, состоят из 5 субъедениц (a1, a2, b, g, d). Главная субъеденица a1 формирует собственно канал и содержит места связывания для различных модуляторов кальциевых каналов. Было обнаружено несколько структурно различных a1 субъедениц кальциевых каналов в нервных клетках млекопитающих (обозначенных как A, B, C, D и E). Функционально кальциевые каналы различных типов отличаются друг от друга активацией, кинетикой, проводимостью одиночного канала и фармакологией. В нервных клетках описано до 6 типов потенциал - управляемых кальциевых каналов (T-, L-, N-, P-, Q-, R- каналы). Активность потенциал - управляемых каналов плазмалеммы регулируется различными внутриклеточными вторичными посредниками и мембраносвязанными G-белками (33,26).
Второй важный путь потока ионов кальция через мембрану связан с активацией агонист - управляемых каналов. Многие агонист - управляемые каналы обладают значительной кальциевой проницаемостью при физиологических условиях. Такую кальциевую проницаемость обнаруживают нейрональные ацетилхолин(Ach)-управляемые, глутамат -управляемые (NMDA и AMPA/Каинат типы) и пуринорецепторы (10,18,49,34). Кроме кальциевых каналов плазмалеммы, управляемых внешними воздействиями, в эукариотических клетках было открыто также несколько типов кальциевых каналов, контролируемых внутриклеточными вторичными посредниками. В частности, IP3-управляемые кальциевые каналы были обнаружены в нейронах Пуркинье мозжечка (34), а IP4-управляемые кальциевые каналы - в клетках эндотелия (35).
Третий, недавно обнаруженный, особый тип Са2+ каналов, который контролируется заполненностью внутриклеточных кальциевых депо, осуществляя таким образом прямую связь между освобождением Са2+ в цитоплазму из депо и вход в нее Са2+ через плазмалемму.
2.1.2 Кальциевые буферыБольшая часть ионов Са2+, входящих в клетку, практически немедленно связывается цитоплазматическими местами связывания кальция. Показано, что только менее 1% ионов кальция, которые проникают в цитозоль, остается в несвязанном состоянии (11). Цитозольные кальциевые буферы представлены главным образом Са2+-связывающими белками, такими как парвальбумин, кальмодулин, тропонин-С, кальретинин, кальциунеурин, белок S-100 (25). Кроме того, цитозольная буферная емкость может быть опосредована АТФ, которая способна связывать значительное количество Са2+ (64). 20-50% цитозольных кальциевых буферов могут быть удалены из цитоплазмы при перфузировании клетки, что показывает их мобильность, в то время как оставшаяся часть Са2+-связывающей емкости относится к фиксированным буферам. Мобильные кальциевые буферы могут играть важную функциональную роль, способствуя диффузии ионов Са2+ в цитоплазме и распространению Са2+ сигнала по клетке. Внутриклеточное введение эндогенных Са2+ буферов (кальбиндина D28k и парвальбумина) через микропипетку приводило к увеличению скорости нарастания [Ca2+]i на несколько порядков и существенно влияло на кинетику изменения [Ca2+]i, что подтверждает роль мобильных Са2+ буферов с низким молекулярным весом в эффективном регулировании внутриклеточной концентрации кальция.
2.1.3 Кальциевые каналы эндоплазматического ретикулумаСа2+ каналы ЭР являются олигометрическими протеинами, встроенными в мембрану ЭР. Эти каналы можно относительно легко выделить из клетки для дальнейшего структурного анализа благодаря тому, что белки канала связываются специфически и с высоким сродством с IP3 (для IP3-управляемых каналов) и с рианодином (для Са2+-управляемых каналов).
Са2+-управляемые Са2+ каналы ЭР. Эти кальциевые каналы были впервые выделены из скелетных и сердечных мышц. Оказалось, что Са2+ каналы ЭР в этих мышечных тканях имеют молекулярные различия и закодированы различными генами. Са2+ каналы ЭР в сердечных мышцах непосредственно связаны с высокопороговыми Са2+ каналами плазмалеммы (L-тип) через кальцийсвязывающие белки, образуя, таким образом, функционально активную структуру - «триаду». В скелетных мышцах деполяризация плазмалеммы прямо активирует освобождение Са2+ из саркоплазматического ретикулума благодаря тому, что Са2+ каналы плазмалеммы служат потенциал - чувствительными передатчиками активирующего сигнала непосредственно Са2+ каналам ЭР через связывающие белки (44). Таким образом, Са2+ депо скелетных мышц обладают механизмом освобождения Са2+, вызываемым деполяризацией (RyR1-тип). В отличие от скелетных мышц, саркоплазматические Са2+ каналы кардиомиоцитов не связаны с плазмалеммой, и для стимуляции освобождения Са2+ из депо требуется увеличение концентрации цитозольного кальция (RyR2-тип). ДНК, кодирующая белки двух типов каналов Са2+ освобождения, была клонирована из тканей человека и кролика, что дало возможность экспрессировать Са2+-управляемые Са2+ каналы в модельные клеточные системы. Белки, встроенные в липидный бислой, формируют чувствительные к рианодину каналы, активируемые ионами Са2+ (50 нмоль/л) в присутствии АТФ (29). Кроме этих двух типов Са2+-активируемых Са2+ каналов, недавно был идентифицирован третий тип Са2+ каналов ЭР (RyR3-тип), который является продуктом другого гена. Этот третий тип Са2+ каналов ЭР, как было показано, не чувствителен к кофеину (21). Эксперименты, проведенные на нервных тканях, продемонстрировали присутствие всех трех типов Са2+-управляемых Са2+ каналов ЭР в мозге млекопитающих, однако RyR2-тип является доминантным (38). Са2+-управляемые Са2+ каналы ЭР являются гомотетрамерами, состоящими из мономеров с молекулярным весом 500 КД (39)
IP3-управляемые Са2+ каналы ЭР. Существование IP3-управляемых Са2+ каналов впервые было обнаружено в нейронах Пуркинье. Позже было показано, что они встроены в мембрану эндоплазматического ретикулума. Структура IP3-управляемых Са2+ каналов сходна со структурой Са2+-управляемых Са2+ каналов ЭР. Они также являются гомотетрамерами с молекулярным весом мономера 260 КД. 50% этих каналов активируется 15 мкмоль/л IP3 и блокируется рутением красным и La3+. IP3-управляемые Са2+ каналы были выделены из мозга млекопитающих, и их аминокислотная последовательность была расшифрована. Было показано, что семейство генов, экспрессирующих IP3-управляемые Са2+ каналы, состоит из трех или четырех различных генов; они характеризуются различной чувствительностью к IP3 и по-разному распределены в мозге млекопитающих (45). Порог активации этих каналов варьирует между 0.2 - 0.5 мкмоль/л в нейронах Пуркинье мозжечка и возрастает до 9 мкмоль/л в астроцитах.
2.1.4 Кальциевые насосыСуществует два семейства Са2+ насосов, ответственных за устранение ионов Са2+ из цитоплазмы: Са2+ насосы плазмалеммы и Са2+ насосы эндоплазматического ретикулума. Хотя они относятся к одному семейству белков (так называемому P-классу АТФ-аз), эти насосы обнаруживают некоторые различия в строении, функциональной активности и фармакологии.
Кальциевый насос плазмалеммы. Са2+ насос плазмалеммы, который удаляет ионы Са2+ из цитоплазмы в межклеточное пространство, был открыт в 1966 году. Молекулярные свойства Са2+ насосов плазмалеммы описаны в нескольких обзорах (18), однако достоверных данных о скорости вывода Са2+ и регуляции Са2+ насосов в нервных клетках немного. Недавно был разработан двухфлуоресцентный микрокапельный метод (58), позволяющий одновременно измерять [Ca2+]i и выход Са2+ наружу на одиночных клетках. Исследования, проведенные с помощью данного метода на нейронах моллюска и секреторных клетках, показали, что активность Са2+ насоса плазмалеммы контролируется непосредственно [Ca2+]i: увеличение концентрации цитоплазматического кальция активирует Са2+ насос (58). В нейронах моллюска около 40% ионов кальция, входящих в клетку в ответ на деполяризацию мембраны, выводится из нейрона уже во время фазы нарастания [Ca2+]i, отражая таким образом активацию кальциевого насоса плазмалеммы увеличением концентрации цитозольного Са2+ (58).
Кальциевый насос эндоплазматического ретикулума. Во многих эукариотических клетках, наряду с Са2+ насосом плазмалеммы, существует кальциевый насос сарко(эндо)плазматического ретикулума (SERCA). В настоящее время описано по крайней мере 3 различных изоформы SERCA-насосов в клетках млекопитающих. SERCA1-подтип сосредоточен исключительно в быстрых скелетных мышцах, SERCA2-насосы широко распространены в других тканях. Значимость SERCA3-насосов менее ясна (13). Белки SERCA2-насосов разделяются на две различные изоформы: SERCA2а, характерные для кардиомиоцитов и гладких мышц, и SERCA2b, характерные для тканей мозга. Предполагается, что насосы SERCA различными способами регулируются цитоплазматической и интралюминальной концентрациями Са2+: Увеличение [Ca2+]i активирует захват ионов кальция в ЭР, в то время как увеличение свободного кальция внутри ЭР ингибирует насосы SERCA (12). Насосы SERCA эффективно и селективно блокируются тапсигаргином в наномолярных концентрациях (37) и микромолярными концентрациями циклопиазоновой кислоты. Однако, тапсигаргин вызывает также блокаду потенциал - управляемых кальциевых каналов плазмалеммы, как это показано на клетках коркового слоя надпочечников и на сенсорных нейронах (Shmigol et al., 1995), поэтому его следует использовать с некоторой осторожностью.
2.1.5 Кальциевые обменникиДополнительным механизмом, ответственным за вывод ионов кальция из цитоплазмы, является натрий-кальциевый обменник, который выводит Са2+, используя энергию натриевого электрохимического градиента. Наличие Na+- Са2+ обменника было показано в различных типах возбудимых и невозбудимых клеток; в клетках нервной системы он был обнаружен в конце 60-х годов (9). В нейронах моллюска, помещенных в среду с пониженным натрием (т.е. с обратным натриевым градиентом), наблюдалось увеличение [Ca2+]i, что является результатом работы обменника в инвертированной форме. Однако, вклад Na+- Са2+ обменника в регуляцию [Ca2+]i в нейронах млекопитающих до сих пор не оценен. В некоторых работах было показано, что обменник принимает незначительное участие в удалении цитоплазматического Са2+, в то время как в других работах представлены данные о том, что обменник играет существенную роль в переносе Са2+ через мембрану (57).
2.1.6 Са2+-связывающие органеллыКроме быстрого связывания цитозольного Са2+ внутриклеточными Са2+-связывающими белками, ионы кальция, попадающие в цитозоль, могут аккумулироваться аппаратом Гольджи или клеточным ядром, захватываться митохондриальными Са2+ депо, имеющими достаточно невысокое сродство к Са2+, или быстрыми депо, связанными с ЭР или СР, имеющими высокое сродство к Са2+. Однако если [Ca2+]i превышает 0,5 мкмоль/л, наблюдается существенное перераспределение [Ca2+]i в область митохондрий. Буферные системы митохондрий принимают участие в удалении избыточного Са2+ из цитоплазмы в клетках кишечника, некоторых типах нервных клеток (59) и в секреторных клетках после повышения [Ca2+]i, стимулированного агонистами. Связывание кальция митохондриями обеспечивается активностью систем, расположенных на внутренней митохондриальной мембране. Са2+ поступает в митохондрии по электрохимическому градиенту; разность потенциалов, обеспечивающая транспорт кальция, создается переносом электронов во время клеточного дыхания и связанного с ним переносом протонов. Перенос электронов по дыхательной цепи является основным механизмом, обеспечивающим энергетику транспорта кальция. Подавление дыхательной цепи карбонил-цианид-м-хлорофенил-гидразоном (СССР) эффективно блокирует аккумуляцию кальция митохондриями (41).
2.2 Влияние АТФ на кальциевый гомеостазПоследние исследования показали, что АТФ занимает прочное место в ряду нейромедиаторов центральной и периферической нервной систем (Burnstock 1990). Не вызывает сомнения, что АТФ является не только важнейшим внутриклеточным метаболитом, но и служит важным объектом межклеточного взаимодействия.
2.2.1 Строение и свойства АТФ
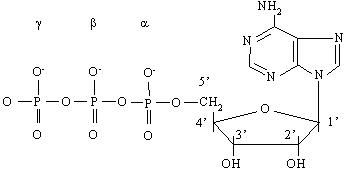 |
АТФ (см. рис.1) представляет собой нуклеотид и как всякий нуклеотид состоит из трех компонентов: азотистого основания, сахара пентозы и фосфата. В качестве азотистого основания в нуклеотидах присутствуют производные пурина и пиримидина. Фосфаты соединены в полифосфатную цепь, количество которых в естественных нуклеотидах не превышает трех. Однако синтезированы нуклеотиды, содержащие линейные цепи из более чем 3-х фосфатов, к примеру аденозинтетра- и аденозинпентафосфаты.
Названия нуклеотидов, содержащих в качестве сахара рибозу, складываются из названия соответствующего нуклеозида, приставки, обозначающей количество фосфатных групп в нуклеотиде и слова фосфат. Для наиболее распространенных нуклеотидов приняты сокращенные названия, например АТФ для аденозинтрифосфата, ГТФ - для гуанозинтрифосфата, ИМФ - инозинмонофосфата.
В области нейтральных значений pH нуклеиновые основания и рибоза в растворе не заряжены (Мартин, Мариам, 1982). Нуклеотиды, из-за наличия фосфатов, представляют собой сильные кислоты. АТФ содержит четыре ОН группы, способные к ионизации, три из которых имеют pKa ниже 3, а pKa четвертой - 6,5 (Ленинджер, 1976). Таким образом, при pH 7,4 подавляющее большинство молекул АТФ представляют собой четырехзарядные анионы АТФ4- , кроме того, в растворе присутствует небольшое количество АТФ3-.
2.2.2 Номенклатура и субклассификация пуринорецепторов.Первое разделение пуринорецепторов на Р1 и Р2 типы основывалось на следующем критерии: нуклеозиды такие как аденозин активировали Р1 пуринорецепторы, в то время как АТФ стимулировала Р2 пуринорецепторы, а метилксантины (кофеин, теофиллин) являются селективными антагонистами Р1 пуринорецепторов. Также Р1 пуринорецепторы связаны с аденилатциклазой, а активация Р2 пуринорецепторов может приводить к выработке простогландинов (Burnstock 1978).
2.2.3 Р1 пуринорецепторы.В 1979 году (Van Calker et al 1979) показали, что аденозиновые, Ð1 - пуринорецепторы можно подразделить на две группы. Рецепторы одной из них обладали очень высоким сродством к аденозину (константа диссоциации Кd = 3 - 10 нМ). При взаимодействии аденозина с этой группой рецепторов наблюдалось ингибирование аденилатциклазы и, следовательно, уменьшался уровень внутриклеточного цАМФ. Этот класс рецепторов был назван А1 -рецепторами (Ri рецепторы по (Londos et al 1980)). Аденозиновые рецепторы, относящиеся ко второй группе, имели более низкое сродство к аденозину (Кd комплекса аденозина с рецептором 5-10 мкМ). Активация этого типа рецепторов приводила к стимуляции аценилатциклазы. Эти рецепторы Ван Колкер назвал А2-рецепторами (Ra рецепторы по (Londos et al 1980)). Были обнаружены также и мощные антагонисты для А1 - R-N6-Фенилизопропиладенозин (R-PIA). Для А2 более сильным антагонистом был 5’-N-этилкарбоксамидаденозин (NЕСА). Также существуют работы, в которых описываются аденозиновые рецепторы, не связанные с аденилатциклазой эти пуринорецепторы было предложено назвать А3 (Ribeiro and Sebastiao 1986). Предпосылкой к разделению А2 пуринорецепторов на А2а и А2b подтипы стало открытие разного сродства NECA к Р1 пуринорецептору, - высокого в стриатуме (А2а) и низкого в фибробластах (А2b) (Bruns et al 1986).
2.2.4 Р2 пуринорецепторыНакопленные фармакологические доказательства, как-то: тип ответа, порядок активности агонистов, десенситизация, вызываемая АТФ и ее структурными аналогами, послужили основой для первого субделения Р2 пуринорецепторов. В 1985 году Бернсток и Кеннеди (10) указали на неоднородность популяции Р2-пуринорецепторов. Этот вывод был сделан исходя из того, что в некоторых тканях (например продольная мышца слепой кишки) АТФ вызывал расслабление гладких мышц, а в других (мышечная стенка мочевого пузыря) - сокращение. Первый подтип рецепторов был обозначен как Р2у, а второй - Р2х. Для Р2х пуринорецепторов наиболее активным агонистом был a, b - метилен АТФ (a,b-мАТФ), а для Р2у - 2 - метилтио АТФ (2-МеSАТP). Так как порядок активности лигандов может зависеть от скорости их гидролиза до неактивных соединений, которая может значительно различаться в зависимости от ткани, более четким доказательством для подразделения Р2 - рецепторов на подтипы служит наличие селективных блокаторов. Сейчас известны селективные антагонисты как для Р2х, так и Р2у-рецепторов для Р2х - a,b-мАТФ (десенситизирующее действие), арилазидаминопропионил АТФ (АНАПП3), пиридоксалфосфат-6-азофенил-2’,4’-дисульфидная кислота (PPADS), сурамин и другие; для Р2у пуринорецепторов - реактив голубой 2, сурамин. В 1986 году Гордон (23) предложил выделить из класса Р2 пуринорецепторов еще два подтипа Р2t и Р2z. Первый рецептор располагается в тромбоцитах и управляет их агрегацией. Он, в отличие от всех остальных подтипов P2 - рецепторов, активируется АДФ, и блокируется АТФ. Р2z рецепторы расположены в тучных клетках. В них АТФ в очень больших концентрациях (> 100 мкМ), а точнее четырехзарядный анион АТФ4-, вызывал кальцийзависимую секрецию гистамина. Другие пуриновые нуклеотиды, включая негидролизуемые аналоги АТФ, не активировали рецептор. Также был обнаружен неселективный антагонист данного типа рецепторов DIDS - аналог PPADS (Soltoff et al 1993). В 1991 году O’Connor предложил так называемый нуклеотидный рецептор одинаково чувствительный как к АТФ, так и к УТФ, но не чувствительный к 2 - MeSATP. Этот рецептор получил обозначение Р2u. Возможным антагонистом данного рецептора является сурамин. Также было обнаружено, что аденин динуклеотид полифосфаты также присутствуют в фармакологической периферии (Hoyle 1990). Hildermann (1991) индентифицировал сайт связывания для диаденозин тетрафосфата ( Ap4A) в мозге крысы, который получил название дипуринергического рецептора, а позднее Р2d (Pintor et al 1993). Антагонисты данного вида пуринорецепторов пока не обнаружены.
| пуринорецепторы | ||||||||||||||||||||||||||||
| Р1 - пуринорецепторы | Р2 - пуринорецепторы | |||||||||||||||||||||||||||
| A1 | A2a | A2b | A3 | P2x | P2y | P2t | P2z | P2u | P2d | |||||||||||||||||||
Таблица 1. Классическая схема субклассификации пурино-рецепторов.
2.2.5 Реклассификация пуринорецепторов.Из - за всевозрастающих трудностей и несогласований в классической схеме классификации Ð2 пуринорецепторов и увеличивающемся числе подтипов рецепторов, стало очевидным, что классическая схема требует пересмотра. В 1994 году Abbracchio and Burnstock предложили новую схему классификации Р2 - пуринорецепторов. Из всех Р2 - пуринорецепторов они вычленили три основных семейства: Р2Х семейство, связанное с ионотропными каналами, которое включало четыре подтипа; Р2У - связанное с активацией G - белков, включающее семь подтипов и семейство Р2Z - семейство неселективных пор. Их гипотеза основывалась в основном на изучении литературных источников и анализе фармакологического профиля новообнаруженныж агонистов. В дальнейшем теория подтвердилась клонированием различных подтипов Р2 - пуринорецепторов. В настоящее время семейство Р2Х насчитывает шесть, а Р2У - семь подклассов. Благодаря интенсивным исследованиям, практически не остается сомнений в том, что данные семейства будут расти и дальше (Collo et al 1996).
| Р2 - пуринорецепторы | ||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| P2Z - неселективные поры | ||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| Р2Х - семейство ионотропных рецепторов | Р2У - семейство метаботропных рецепторов | |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| Р2Х1 | Р2Х2 | Р2Х3 | Р2Х4 | Р2Х5 | Р2Х6 | Р2У1 | Р2У2 | Р2У3 | Р2У4 | Р2У5 | Р2У6 | Р2У7 |
| ||||||||||||||||||||||||||
Таблица 2. Современная классификация Р2 типа пуринорецепторов.
3. ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЙ 3.1 Подготовка препарата
Исследования проводились на пирамидальных нейронах моторной области новой коры в тонких срезах мозга, выделенного из 14 дневных крыс. После декапитации мозг помещался в холодный солевой раствор (0 - 40С). Процедура от начала декапитации до выделения мозга длилась не более 60-90 секунд. Затем мозг закреплялся полиакриламидным клеем на подложке вибротома (Campden, Campden Instruments LTD, U.K.); камера вибротома заливалась холодным солевым раствором. Срезы нарезались сагитально толщиной 250 - 300 мкм; скорость подачи лезвия - 1 см/с с частотой 10 Гц. После приготовления срезы помещались в раствор постоянно насыщаемый карбогеном (5% СО2 + 95% О2). Перед загрузкой флуоресцентным красителем срезы инкубировались в постоянно оксигенируемом растворе 30 минут при температуре 32 градуса. Окраска среза осуществлялась в течении 30 - 35 минут в СО2 насыщенном термостате при температуре 35 градусов. После окраски срезы отмывались 1 - 1,5 часа в постоянно оксигенируемом растворе при комнатной температуре. Все эксперименты проводились при температуре 32 градуса.
3.2 Характеристики кальциевого зонда|
|
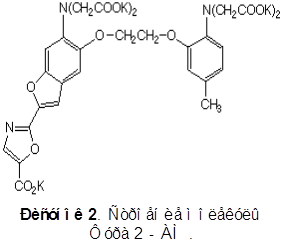
Для количественного определения концентрации кальция в пирамидальных нейронах моторной области новой коры использовался краситель Фура-2 AM (рисунок 2) (16).
|
|
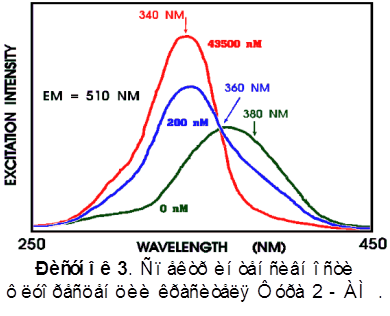
На рисунке 3 представлен спектр зонда Фура -2 AM. При связывании с кальцием происходит характерное изменение спектра возбуждения этого красителя: при возбуждении светом с длиной волны 390 нм происходит уменьшение флуоресценции, а при возбуждении светом, с длиной волны 340 нм - увеличение. Однако, 340 нм является уже ультрафиолетовым светом, и для ее использования необходим микроскоп с кварцевыми линзами. Поскольку в нашем случае был использован обычный микроскоп, то вторая волна была выбрана длиной 390 нм. 360 нм - это изобестическая точка зонда Фура -2, т.е. флуоресцентный сигнал при возбуждении светом этой длины волны не зависит от концентрации Ca2+ и есть функция лишь концентрации зонда.
Измерение флуоресценции при возбуждении двумя длинами волн позволяет легко рассчитать [Ca2+]i в клетке по формуле (24):
[Ca2+]i= Kd * b * (R - Rmin)/(Rmax-R)
где Кd - константа диссоциации комплекса Фура -2 с кальцием, R=F360/F390 - текущее отношение флуоресцентных сигналов, Rmin = F360/F390|Ca0 - то же отношение в растворе с низкой концентрацией Ca2+, Rmax = F360/F390|CaҐ - то же отношение в растворе с высокой концентрацией Ca2+, b = F390|Ca0/F390|CaҐ - отношение флуоресцентных сигналов в низкой и высокой концентрации Ca2+ при возбуждении длиной волны 390 нм. Параметры Rmin, Rmax и b определяли экспериментально. Для этого были приготовлены базовый раствор (в ммоль/л) : KCl - 100, Tris-Cl - 10 (pH=7.2), Фура -2 - 0.005; раствор с низкой концентрацией Ca2+ готовился без добавления Ca2+ с добавкой EGTA - 10; раствор с высокой концентрацией Ca2+ - с добавкой CaСl2 - 10. Отношение величин флуоресценции определялось в каплях приведенных выше растворов, помещенных на дно экспериментальной камеры. В результате для нашей системы Rmin= 0.33, Rmax= 8.9 и b=12.8. Параметр Кd был взял из работы [Grinkiewicz et al, 1985] и равнялся 224 нмоль/л.
3.3 Окраска срезов флуоресцентным красителемДля введения кальций - чувствительного зонда в клетку, срезы инкубировались в растворе Тироде, содержащим эстерифицированную незаряженную форму красителя Фура-2 ацетоксиметилэстер в концентрации 5 мкмоль/л, растворенную в диметилсульфоксиде с добавлением детергента Плуроник F-127 (0.02%). В такой форме краситель проникает в клетку, затем эндогенными эстеразами эфирные группы отщепляются, зонд становится заряженным и покинуть клетку не может. Окраска производилась 20 минут при температуре 35оС. Концентрация зонда в клетке определялась путем титрования окрашенных клеток раствором красителя, который добавлялся во внеклеточный раствор. Оцененная таким способом концентрация зонда в клетке была в диапазоне 30 - 70 мкмоль/л.
3.4 Структура экспериментальной установки для двух-волнового измерения концентрации кальцияПринципиальная схема установки представлена на рисунке 4. Основными элементами ее являются: источник света, система смены фильтров, микроскоп, ФЭУ, модуль предварительной обработки сигнала и компьютер.
Источником возбуждающего света служит ртутная лампа мощностью 50 ватт. Поскольку в качестве кальций - чувствительного зонда использовали индикатор Фура -2 АМ, являющийся по возбуждению двухволновым (см. выше), то для количественного определения [Ca2+]i необходимо было одновременно индуцировать флуоресценцию обеими длинами волн. В нашей конфигурации периодическая смена фильтров возбуждения была реализована при помощи вращения колеса с вмонтированными в него двумя интерференционными фильтрами 360 и 390 нм. Частота вращения была 5 Гц. Управление системой смены фильтров осуществлялось при помощи кальций - измерительного модуля фирмы Luigs und Neumann, Германия. Этот же модуль являлся интерфейсом предварительной обработки.
Оптическая часть установки была смонтирована на базе микроскопа Axioskop, Zeiss. Разделение потоков возбуждающего света и флуоресценции производилось при помощи дихроического зеркала. Свет, прошедший через фильтр для возбуждения флуоресценции, отклонялся дихроическим зеркалом и при помощи высокоаппертурного иммерсионного объектива (40*, апертура 0.75) фокусировался на объекте. Флуоресцентный свет проходил через соответствующий интерференционный фильтр и подавался на фотоэлектронный умножитель (ФЭУ). Для уменьшения уровня шумов фиксированная диафрагма (1 мм) была расположена перед ФЭУ. В результате флуоресцентный сигнал собирался с фокальной плоскости диаметром 50 мкм, что позволило значительно улучшить соотношение сигнал/шум.
Модуль предварительной обработки позволял производить компенсацию автофлуоресценции и, при необходимости, усиливать сигнал, после чего он оцифровывался при помощи цифрового интерфейса TIDA и обрабатывался компьютером при помощи программ Tida 5.0 и Wintida, разработанных в Гейдельберге, Германия.
|
|
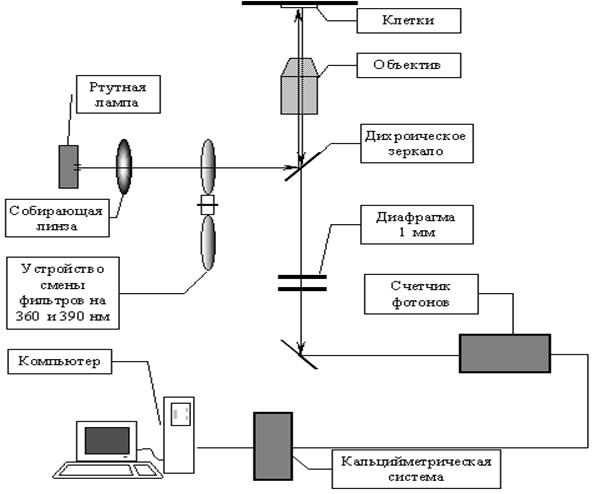
Рисунок 4. Принципиальная схема экспериментальной установки для двухволнового измерения кальция.
3.5 Растворы и смена растворовПри работе со срезами все растворы готовились на основе раствора Тироде (в ммоль/л): NaCl - 125, KCl - 5.4, MgCl2 - 1, CaCl2 - 2.5, NaHCO3 - 26, KH2PO4 - 1.6, глюкоза - 10, постоянно аэрировавшийся газовой смесью 95% О2 + 5% СО2 (pH=7.4). Бескальциевый раствор готовился эквимолярной заменой CaCl2 на MgCl2 и добавкой 1 ммоль/л EGTA, растворы с повышенным содержанием калия или кофеина - заменой NaCl на соответствующее количество KСl или кофеина.
Смена растворов производилась протоком, скорость которого можно было варьировать от 10 до 20 мл/мин. Объем экспериментальной камеры составлял 0.5 мл. Все эксперименты были проведены при температуре (32 °С).
4. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
|
|
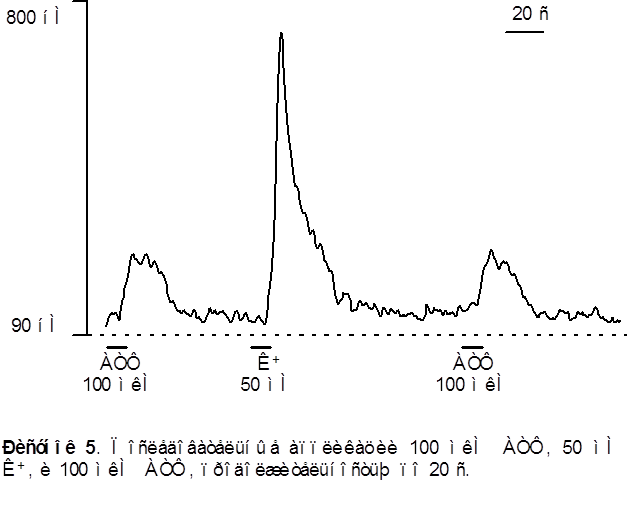
Аппликация АТФ в большинстве нейронов моторной коры (98 из 102) вызывает быстрое и кратковременное повышение концентрации цитозольного кальция [Ca2+]i . Концентрация [Ca2+]i достигает своего максимума на протяжении 6-8 секунд и затем восстанавливается до базального уровня при продолжающемся присутствии АТФ. Отмывание АТФ приводит к восстановлению [Ca2+]i до уровня покоя. Характерный вид АТФ - индуцированного кальциевого транзиента представлен на рисунке 5.
|
|
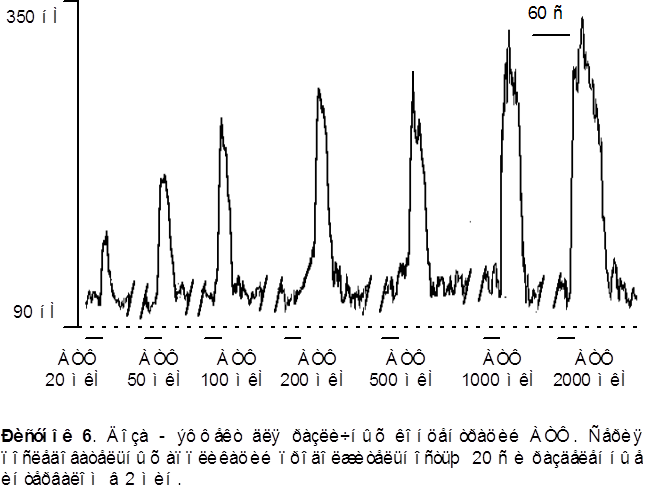
Амплитуда АТФ - индуцированного кальциевого транзиента варьировалась в пределах от 50 до 450 нМ. Среднее значение выброса [Ca2+]i вызванного приложением АТФ в концентрации 100 мкМ находилось в пределах 200-250 нМ.
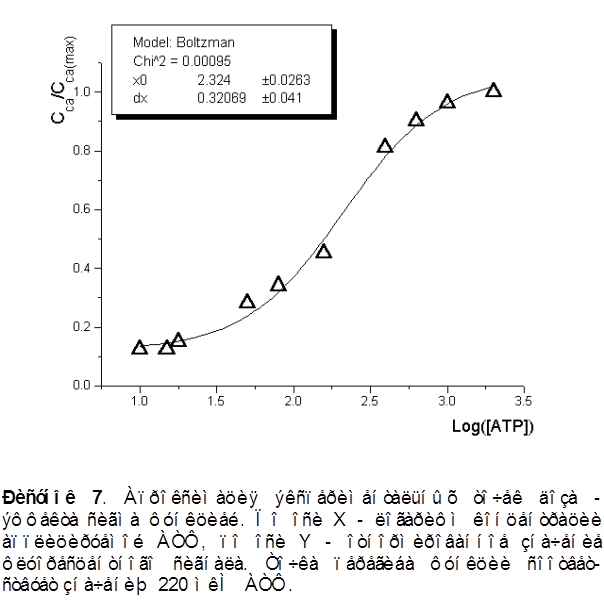
Наблюдалась доза - зависимость амплитуды ответа от концентрации АТФ в пределах от 10 до 2000 мкМ. На рисунке 7 представлена апроксимация экспериментальных точек сигма - функцией. Концентрация АТФ, при которой амплитуда ответа составляет половину максимальной, - 220 ± 20 мкМ.
|
|
Целью наших исследований было определение подтипов пуринорецепторов вовлеченных в генерацию [Ca2+]i транзиента. Известно, что АТФ активирует несколько типов пуринорецепторов, а именно Р2 пуринорецепторы подтипов Р2х и Р2у (Peter Illes et al, 1993, Burnstock and Kennedy 1985), а также Са2+ каналы плазмалеммы (Кришталь, 1983).
С целью обеспечения достоверности полученных данных, принимая во внимание тот факт, что работа проводилась на тонких срезах мозга, мы провели серию экспериментов с подавлением распостранения потенциалов действия с помощью тетродотоксина (TTX). При использовании TTX выяснилось, что амплитуда и форма [Ca2+]i транзиентов не изменяется или изменяется незначительно. Полученные результаты позволяют нам предполагать, что получаемые нами данные отображают процессы происходящие в отдельной наблюдаемой клетке. К сожалению мы не имели возможности проводить все эксперименты c тетродотоксином из-за дороговизны препарата. Для изучения характеристик рассматриваемых пуринорецепторов, принимающих участие в генерации [Ca2+]i транзиента, применялись следующие протоколы экспериментов.
1. Приложение АТФ в растворе не содержащем Ca2+. Удаление кальция из внеклеточного раствора незначительно влияло на кальциевый транзиент, вызванный аппликацией 10 мкМ АТФ, но в тоже время подавляло на 45% ± 7% кальциевый транзиент вызванный 100 мкМ АТФ. (Рис. 8).
|
|

1. Для исследования способности внутриклеточного Ca2+ депо к перезаполнению мы последовательно апплицировали АТФ несколько раз с интервалом в 60 секунд в обычном растворе и в растворе не содержащем Ca2+ . Как видно из рисунка 9 вторая аппликация АТФ вызывала Ca2+ транзиент меньшей (на 30% ± 4%) амплитуды по сравнению с первой (контрольной) аппликацией. Последующие аппликации АТФ разделенные 60 ти секундным интервалом не вызывали заметного уменьшения амплитуды [Ca2+]i транзинета по сравнению со второй аппликацией в стандартном растворе
|
|
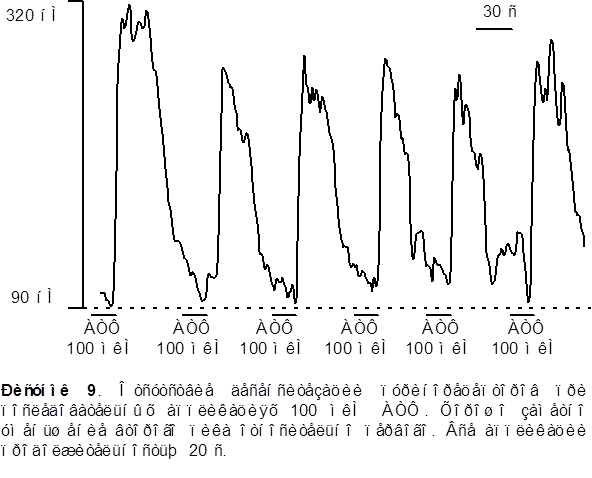
В бескальциевом растворе - наоборот, вторая аппликация АТФ вызывала значительное уменьшение Ca2+ транзиента на 40% ± 5% от контроля. Последующие аппликации АТФ в бескальциевом растворе приводили к последовательному уменьшению амплитуды [Ca2+]i транзиента и после двух - трех последовательных аппликаций сигнал исчезал вообще (рисунок 10). Такое уменьшение вероятно обусловлено истощением внутриклеточных депо.
|
|

1. Ингибирование захвата Ca2+ эндоплазматическим ретикулом тапсигаргином (спецефическим блокатором Ca2+ насоса эндоплазматического ретикулума) уменьшало [Ca2+]i транзиенты, вызванные приложением АТФ, на 63% ± 5% для концентрации АТФ 100 мкМ АТФ (рис. 11). Как видно из рисунка 11 приложение тапсигаргина не влияло на базальный уровень Ca2+ в клетке.
1. Приложение кадмия в концентрации 50 мкМ, верапамила в концентрации 100 мкМ и 50 мкМ никеля обратимо уменьшало амплитуду [Ca2+]i транзиента вызванного приложением 100 мкМ АТФ на 35%, 20%, и 15% сответственно. Данные представлены на рисунке 12.
|
|

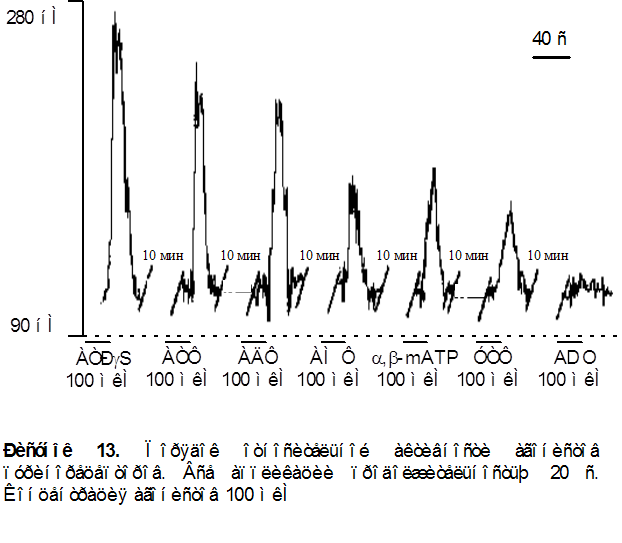
1. Приложение различных агонистов пуринорецепторов вызывало различные по амплитуде ответы. Порядок относительной активности лигандов для данного объекта был следующим ATPgS > ATФ = AДФ >> a,b-methylene ATP » AMФ > УТФ>> аденозин (ADO), данные преставлены на рисунке 13.
|
|
1. Сурамин, известный антагонист Р2 типа пуринорецепторов, уменьшал [Ca2+]i транзиенты, вызванные приложением 100 мкМ АТФ на 76% ± 7% и также не изменял концентрацию [Ca2+]i в покое. Данные представлены на рисунке 14.
|
|

5. ОБСУЖДЕНИЕ
При исследовании средних слоев неокортекса крыс мы обнаружили их способность отвечать на приложение АТФ. Таким образом мы можем сделать вывод о наличии в данном объекте пуринорецепторов. Ответ на АТФ является доза - зависимым с амплитудой половинного ответа в 220 нМ. Последовательные приложения АТФ, после второго приложения, не вызывали уменьшения амплитуды [Ca2+]in транзиентов. Из этого можно сделать вывод об отсутствии десенситизации данного типа рецепторов.
Исследуя источники повышения цитозольного кальция мы обнаружили, что АТФ активирует как ионотропные так и метаботропные рецепторы. Блокаторы потенциал - управляемых кальциевых каналов такие как кадмий, никель и верапамил уменьшали АТФ - индуцированные кальциевые транзиенты на 35% - 15%, что говорит об опосредованном АТФ активировании потенциал - управляемых каналов.
Для исследования активацию метаботропных рецепторов. Для этого мы прилагали АТФ в бескальциевом растворе, - амплитуда ответа при этом уменьшалась на 45% ± 7%. Этот результат говорит о том, что внутриклеточные депо принимают участие в генерации [Ca2+]in ответов, причем их вклад близок к половине. При повторных аппликациях АТФ в бескальциевом растворе Ca2+ ответ исчезал полностью после второй аппликации, т.е. внутриклеточные депо полностью истощаются. Исследуя путь высвобождения внутриклеточного кальция мы апплицировали тапсигаргин - спецефический блокатор АТФ - азы эндоплазматического ретикулума. [Ca2+]in транзиенты уменьшались на 63% ± 5% в присутствии тапсигаргина. Аппликация коффеина, агониста рианодиновых рецепторов, в клетках моторной коры 14 дневных крыс не вызывали повышения уровня [Ca2+]in. Следовательно выброс [Ca2+]in из внутриклеточного депо происходит по IP3 - чувствительному механизму.
В дальнейшем мы исследовали более подробно типы присутствующих пуринорецепторов. Построив ряд активности агонистов для Р1 и Р2 пуринорецепторов по амплитудам ответов, мы сделали заключение базирующееся на отсутствии ответа на аденозин, что в нашем объекте присутствуют только Р2 пуринорецепторы. Проводя дальнейшую субклассификацию Р2 типа рецепторов мы использовали сурамин - блокатор некоторых типов Р2х и Р2у рецепторов. Приложение сурамина уменьшало амплитуду [Ca2+]i транзинета вызванного приложением 100 мкМ АТФ на 76% ± 5%, что говорит о наличии этих типов рецепторов в исследуемом объекте.
6. ВЫВОДЫ
1. Приложение АТФ в различных концентрациях вызывает Ca2+ транзиенты в клетках моторной коры крыс.
2. Ответ на АТФ является доза - зависимым с амплитудой половинного ответа в 220 нМ.
3. АТФ активирует как ионотропный так и метаботропные пути повышения внутриклеточного кальция.
4. В генерации АТФ индуцированного повышения [Ca2+]in принимают участие некоторые типы потенциал - управляемых кальциевых каналов.
5. Высвобождение внутриклеточного кальция происходит из IP3 чувствительных депо.
6. В данном объекте присутсвуют только Р2 подтипы пуринорецепторов.
7. Сурамин - антагонист Р2Х2 и Р2Х5 и Р2У рецепторов уменьшает амплитуду [Ca2+]in транзиенты, что говорит присутствии некоторых из вышеперечисленных рецепторов.
7. СПИСОК ЛИТЕРАТУРЫ
1. A.Shmigol, A.Verkhratsky & G. Isenberg (1995): Calcium-induced calcium release in rat sensory neurones. Journal of Physiology (London), 489.3 627-636.
2. A.Shmigol, G. Isenberg, P.Kostyuk & A. Verkhratsky (1994): Calcium-induced Ca2+ release from internal stores in rat dorsal root ganglion neurones. In: European Journal of Neuroscience, Suppl. 7, Proceedings of the 16th Annual ENA Meeting, p. 146.
3. A.Shmigol, N.Svichar, P.Kostyuk & A.Verkharatsky. (1995): “Incremental” caffeine-induced calcium release in mouse sensory neurones. European Joutnal of Neuroscience, Supple № 8. p111. Proceedings of the 16th Annual ENA Meeting.
4. A.Shmigol, Yu.Usachev, N.Pronchuk, S.Kirischuk, P.Kostyuk & A.Verkhratsky (1994): Properties of the caffeine sensitive intracellular calcium stores in mammalian neurons. Neurophysiology /Neirophiziologia, v. 26 No. 2, p. 16 - 25.
5. A.Verkhratsky, A. Shmigol, S. Kirischuk, N. Pronchuk & P. Kostyuk (1994): Age-dependent changes in calcium currents and calcium homeostasis in mammalian neurons. Annals of the New York Academy of Sciences, v. 747, p365 - 381.
6. Abbracchio, M. P., Burnstock, G. (1994) Purinoceptors: are there families of P2x and P 2y purinoceptors? Pharmac. Ther. 64: 445-475
7. Anatoly Smigol, Platon Kostyuk, Alexey Verhratsky (1994) Role of caffeine-sensitive Ca2+ stores in Ca2+ signal termination in adult DRG neurones // NeuroReport v.5, 2073-2076.
8. Anatoly Smigol, Sergey Kirischuk, Platon Kostyuk, Alexey Verhratsky (1994) Different properties of caffeine-sensitive Ca2+ stores in peripherial and central mammalian neurones // Pflugers Arch v.426, 174-176.
9. Baker P. F., Blaustein M.P., Hodgkin A.L. and Steinhardt R. A. (1969) The influence of calcium on sodium efflux in squid axons. J. Physiol., Lond. 200, 431‑458.
10. Bean B.P. (1992) Pharmacology and electrophysiology of ATP‑activated ion channels. Trends Pharmacol. Sci. 13, 87 ‑ 90.
11. Belan P., Kostyuk P., Snitsarev V. and Tepikin A. (1993) Calcium clamp in isolated neurones of the snail Helix pomatia. J. Physiol., Lond. 462, 47 ‑ 58.
12. Bronner, F. (1990). Intracellular Ca2+ regulation.. New York: Wiley‑Liss.
13. Burk S. E., Lytton J. , MacLennan D. H. and Shull G. E. (1989). cDNA cloning, functional expressing, and mRNA tissue distribution of a third organellar Ca2+ pump. J. Biol. Chem. 164, 18561‑18568.
14. Burnstock, G. (1972) Purinergic nerves. Pharmacol. Rev. 24: 509-581
15. Burnstock, G. (1978) A basis for distinguishing two types of purinergic receptor. in: book
16. Burnstock, G. (1990) Co-transmission. Arch. Int. Pharmacodyn. 304: 7-33
17. Burnstock, G., Kennedy, C.(1985) Is there a basis for distinguishing two types of P2 purinoceptor? Gen.Pharmacol. 16: 433-440
18. Carafoli E. (1992) Calcium pump of the plasma membrane. Physiol. Rev. 71, 129 ‑ 153.
19. Chen, C.-C., Akopian, A.N. et al, (1995) A P2x purinoceptors expressed by a sybset of sensory neurones. Nature 377: 428 - 431
20. Кришталь О.А., Марченко С.М. (1983). Рецепторы АТФ в сенсорных нейронах млекопитающих. Докл. Акад. Наук УССР.
21. Gianini G., Clementi E., Ceci R., Marziali G., and Sorremtino V. (1992) Expression of a ryanodine receptor Ca2+ that is regulated by TGF‑b, Science, 257, 91 ‑ 94.
22. Ginetta Collo et al, (1996) Cloning of P2X5 andP2X6 receptors and the distribution and properties of an extended family of ATP-gated ion channels. The J. of Neurosci. 16(8): 2495-2507
23. Gordon, J. L. (1986) Extracellular ATP: effects, sources and fate. Biochem.J. 233: 309-319
24. Grynkiewicz, G., Poenie, M., and Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescent properties. J. Biol. Chem., 260, 3440-3450, 1985.
25. Heizmann C.W. and Hunziker W. (1991) Intracellular calcium‑binding proteins: more sights than insights. Trends Biochem. Sci. 16, 98 ‑ 103.
26. Heschler J. and Schultz G. (1993) G‑proteins involved in the calcium channel signalling system. Curr. Opin. Neurobiol. 3, 360‑367.
27. Hiderman, R. H., Martin, M., Zimmerman, J. K., Pivorun, E. B. (1991) Identification of a unique membrane receptor for adenosin 5¢,5¢¢¢- P1,P4-tetraphosphate. J. Biol. Chem. 266: 6915-6918
28. Hoyle, C. H. V. (1990) Pharmacological activity of adenine dinucleotides in the periphery: possible receptor classes and transmitter function. Gen. Pharmacol. 21: 827-831
29. Hymel L., Inui M., Fleischer S. and Schindler H. (1988). Purified ryanodine receptor of skeletal muscle sarcoplasmic reticulum forms Ca2+‑activated oligomeric Ca2+ channels in planar bilayers. Proc.Natl.Acad.Sci.U.S.A. 85, 441‑445.
30. Kirischuk S.I., Voitenko N.V., Kettenmann H.O. and Verkhratsky A.N. (1994) Mechanisms of cytoplasmic calcium signalling in cerebellar Bergman glial cells // Neurophysiology v.26, 417-419.
31. Kirischuk, V.Matiash, A.Kulik, N.Voitenko, P.Kostyuk, A.Verkhratsky (1996) Activation of P2-purino, a1-adreno and H1-histamine receptors triggers cytoplasmic calcium signalling in cerebellar Pupkinje neurons // Neuroscience v.73, 643-647
32. Kostyuk and A. Verhratsky (1994) Calcium stores in neurones and glia //Neuroscience v. 63, N.2, 381-404.
33. Kostyuk P. G. (1992). Calcium ions in nerve cell function. Oxford, New York, Tokyo: Oxford University Press.
34. Kuno M., Maeda N. and Mikoshiba K. (1994) IP3‑activated Ca2+‑permeable channels in the incide‑out patches of cultured cerebellar Purkinje cells. Biochem. Biophys. Res. Comm. 199, 1128 ‑ 1135.
35. Lуckhoff A. and Clapham D.E. (1992) Inositol 1,3,4,5‑tetrakisphosphate activates an endothelial Ca2+‑permeable channel. Nature 355, 356‑358.
36. Londos, C., Cooper, D. M. F., Wolff, J. (1980) Subclasses of external adenosine receptors. Proc. Natl. Acad. Sci. 77: 2551-2554
37. Lytton J., Westlin M. and Hanley M. R. (1991). Thapsigargin inhibits the sarcoplasmic or endoplasmic reticulum Ca‑ATPase family of calcium pums. Biol. Chem. 266, 17067‑17071.
38. Mackgrill J. J. and Lai F. A. (1994). Solubilization of the type 3 ryanodine receptor from rabbit brain. Biophys. J. 66, A147
39. McPherson P. S., Kim Y. K., Valdivia H., Knudson C. M., Takekura H., Franzini‑Armstrong C., Coronado R. and Campbell K. P. (1991). The brain ryanodine receptor: A caffeine‑sensitive calcium release channel. Neuron 7, 17‑25.
40. N.Voitenko, S.Kirischuk, A.Kulik, A.Verkhratsky (1995) Calcium signalling in granule neurones of the mouse cerebellar slices // Pflugers Archiv European Journal of Physiology, v.430, Supplement 4, R124.
41. Nicholls D.G. (1985) A role for the mitochondria in the protection of the cell against calcium overload. Prog. Brain Res. 63, 97‑106.
42. Pintor, J., Diaz-Rey, M. A., Torres, M., Miras-Portugal, M. T. (1992) Presence of diadenosine polyphosphates-Ap4A and Ap5A-in rat brain synaptic terminals. Ca2+-dependent release evoked by 4-aminopyridine and veratridine. Neurosci. Lett. 136: 141-144
43. Ribeiro, J. A., Sebastiao, A. M. (1986) Adenosine receptors and calcium: basis for proposing a third (A3) adenosine receptor. Prog. Neyrobiol. 26: 179-209
44. Rios E. and Pizarro C. (1991) Voltage‑sensor of excitation‑contraction coupling in skeletal muscle. Physiol. Rev. 76, 849 ‑ 908
45. Ross C. A., Danoff S. K., Schell M. J., Snyder S. H. and Ullrich A. (1992). Three additional inositol 1,4,5‑trisphosphate receptors: Molecular cloning and differential localization in brain and peripheral tissues. Proc. Natl. Acad. Sci. U.S.A. 89, 4265‑4269.
46. S.Kirischuk, N.Voitenko, P.Kostyuk, A.Verkhratsky (1995) Calcium signalling in granule neurones studied in cerebellar slices // Cell Calcium v.18, 464-476
47. S.Kirischuk, N.Voitenko, P.Kostyuk, A.Verkhratsky (1996) Age-associated Changes of Citoplasmic Calcium Homeostasis in Cerebellar Granule Neurones in situ: Investigation on Thin Cerebellar Slices. // Experimental Gerontology
48. S.Kirischuk, N.Voitenko, T.Moller, H.Kettenmann and A.Verkhratsky (1995) ATP-induced cytoplasmic calcium mobilization in bergman glial cells // J. Neuroscience v.15, 8234-8248.
49. Scheggerburger R., Zhou Z., Konnerth A. and Neher E. (1993). Fractional contribution of calcium to the cation current through glutamate receptor channels. Neuron 11, 133‑143.
50. Sergej Kirischuk and Alexej Verkhratsky (1996) [Ca2+]i recordings from neural cells in acutely isolated cerebellar slices employing differential loading of the membrane-permeant form of the calcium indicator fura-2 // Pflugers Arch. -Eur. J. Physiology v.431, 977-983
51. Sergej Kirischuk, Nana Voitenko, Platon Kostyuk, Alexej Verkhratsky (1996) Calcium signalling in granule neurones studied in cerebellar slices // Cell Calcium v.19, 59-71
52. Shmigol A., Kirischuk S., Kostyuk P. and Verkhratsky A. (1994). Different properties of caffeine‑sensitive Ca2+ stores in peripheral and central mammalian neurones. Pflьgers Arch. 426, 174‑176.
53. Shmigol, D.Eisner & A.Verkhratsky (1995): Cyclic ADP ribose enhances Ca2+-induced Ca2+ release in mouse sensory neurones. Journal of Physiology, London, v. 483P, p63P.
54. Shmigol, N. Svichar, P. Kostyuk & A. Verkhratsky (1996) Gradual caffeine-induced Ca2+ release in mice DRG neurones is controlled by cytoplasmic and intraluminal Ca2+. Neuroscience, 73 N 4, 1061-1067
55. Shmigol, P. Kostyuk & A. Verkhratsky (1995): Thapsigargin blocks plasmalemmal voltage-operated calcium channels in mouse DRG neurones. Journal of Physiology, London, v. 483P, p64P.
56. Soltoff, S. P., McMillian, M.K., Talamo, B.R., Cantley, L. C.(1993) Blockade of ATP binding site of P2 purinoceptors in rat parotid acinar cells by isothiocyanate compounds. Biochem. Pharmacol. 45: 1936-1940
57. Tatsumi H. and Katayama Y. (1993) Regulation of intracellular free calcium concentration in acutely dissociated neurones from rat nucleus basalis.J.Physiol., Lond.464,165‑181.
58. Tepikin A. V., Kostyuk P. G., Snitsarev V. A. and Belan P. V. (1992a). Extrusion of calcium from a single isolated neuron of the snail Helix pomatia. J. Membrane Biol. 123, 43‑37.
59. Thayer S.A. and Miller R.J. (1990) Regulation of the intracellular free calcium concentration in single rat dorsal root ganglion neurones in vitro. J.Physiol. (London), 425, 85 ‑ 115.
60. Ursula Windscheif, (1996) Purinoceptors: from history to recent progress. Review. J. Pharm. Pharmacol. 48: 993-1011
61. Van Calker, D., Muller, M., Hamprecht, B. (1979) Adenosine regulates via two different types of receptors, the accumulation of cyclic AMP in cultured brain cells. J. Neurochem. 33: 999-1005
62. Verkhratsky & A.Shmigol (1996) Calcium-induced calcium release in neurones. Cell Calcium, v.19, No 1, 1-14.
63. Voitenko N., Kirischuk S., Verkhratsky A. (1995) Mechanisms of cytoplasmic calcium signalling in cerebellar granule neurones in situ. // Експериментальна та клінічна фізіологія, збірник наукових праць до 100-річчя кафедри фізіології Львівського медичного університету, р.357.
64. Zhou Z. and Neher E. (1993). Calcium permeability of nicotininc acetylcholine receptor channels in bovine adrenal chromaffine cells. Pflugers Arch. 425, 511‑517.
65. Zhou Z. and Neher E. (1993). Mobile and immobile calcium buffers in bovine adrenal chromaffin cells. J.Physiol., Lond. 469, 245‑273.
0 комментариев