Пермская Государственная Медицинская Академия Кафедра биологической химииОбмен веществ и энергии в живых организмах. Выполнила: студентка I курса стомат. факультета 102 группы Бояршинова Анна Андреевна Проверил преподаватель: Поносов Виктор Леонидович Пермь, 2001г.
|
Обмен веществ.
В живых организмах любой процесс сопровождается передачей энергии. Энергию определяют как способность совершать работу. Специальный раздел физики, который изучает свойства и превращения энергии в различных системах, называется термодинамикой. Под термодинамической системой понимают совокупность объектов, условно выделенных из окружающего пространства.
Обмен веществ и энергии - это совокупность физических, химических и физиологических процессов превращения веществ и энергии в живых организмах, а также обмен веществами и энергией между организмом и окружающей средой. Обмен веществ у живых организмов заключается в поступлении из внешней среды различных веществ, в превращении и использовании их в процессах жизнедеятельности и в выделении образующихся продуктов распада в окружающую среду.
Все происходящие в организме преобразования вещества и энергии объединены общим названием - метаболизм (обмен веществ). На клеточном уровне эти преобразования осуществляются через сложные последовательности реакций, называемые путями метаболизма, и могут включать тысячи разнообразных реакций. Эти реакции протекают не хаотически, а в строго определенной последовательности и регулируются множеством генетических и химических механизмов. Метаболизм можно разделить на два взаимосвязанных, но разнонаправленных процесса: анаболизм (ассимиляция) и катаболизм (диссимиляция).
Анаболизм - это совокупность процессов биосинтеза органических веществ (компонентов клетки и других структур органов и тканей). Он обеспечивает рост, развитие, обновление биологических структур, а также накопление энергии (синтез макроэргов). Анаболизм заключается в химической модификации и перестройке поступающих с пищей молекул в другие более сложные биологические молекулы. Например, включение аминокислот в синтезируемые клеткой белки в соответствии с инструкцией, содержащейся в генетическом материале данной клетки.
Катаболизм - это совокупность процессов расщепления сложных молекул до более простых веществ с использованием части из них в качестве субстратов для биосинтеза и расщеплением другой части до конечных продуктов метаболизма с образованием энергии. К конечным продуктам метаболизма относятся вода (у человека примерно 350 мл в день), двуокись углерода (около 230 мл/мин), окись углерода (0,007 мл/мин), мочевина (около 30 г/день), а также другие вещества, содержащие азот (примерно б г/день).
Катаболизм обеспечивает извлечение химической энергии из содержащихся в пище молекул и использование этой энергии на обеспечение необходимых функций. Например, образование свободных аминокислот в результате расщепления поступающих с пищей белков и последующее окисление этих аминокислот в клетке с образованием СО2, и Н2О, что сопровождается высвобождением энергии.
Процессы анаболизма и катаболизма находятся в организме в состоянии динамического равновесия. Преобладание анаболических процессов над катаболическими приводит к росту, накоплению массы тканей, а преобладание катаболических процессов ведет к частичному разрушению тканевых структур. Состояние равновесного или неравновесного соотношения анаболизма и катаболизма зависит от возраста (в детском возрасте преобладает анаболизм, у взрослых обычно наблюдается равновесие, в старческом возрасте преобладает катаболизм), состояния здоровья, выполняемой организмом физической или психоэмоциональной нагрузки.
Цикл Кребса.
Цикл трикарбоновых кислот впервые был открыт английским биохимиком Кребсом. Он первым постулировал значение данного цикла, который источником которого является гликолитическое превращение углеводов. В дальнейшем было показоно, что цикл Кребса является тем центром, где сходятся практически все метаболические пути. Таким образом, цикл Кребса – общий конечный путь окисления ацетильных групп, в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль «клеточного топлива»- углеводов, жирных кислот и аминокислот.
Образовавшийся в результате гликолиза в цикле Эмбдена-Меергофа ацетил-КоА (продукт окислительного декарбоксилирования пирувата) окисляется до воды и углекислого газа в цикле Кребса (лимоннокислый цикл). Этот процесс осуществляется последовательными ферментативными реакциями, в результате которых высвобождается энергия (схема 6). Полный распад одной молекулы глюкозы дает 38 молекул АТФ, причем 24 из них образуются в цикле Кребса. Ферменты этого цикла локализуются в матриксе митохондрий (в стенке внутренней мембраны). Поступивший в цикл Кребса ацетил-КоА является конечным продуктом катаболизма не только углеводов, но также липидов и таких аминокислот, как фенилаланин, тирозин, лейцин и изолейцин.
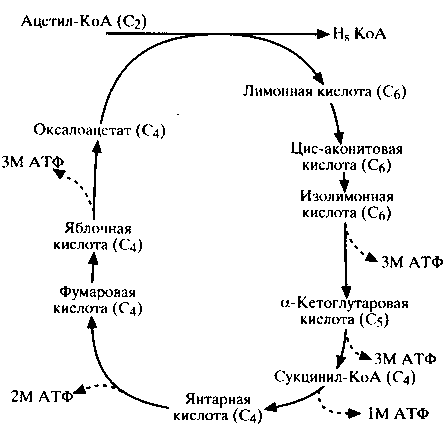
Схема 6. Цикл Кребса (лимоннокислый цикл).
Кроме того, существует прямой путь окисления глюкозы – гексозомонофосфатный (пентозный) цикл, который преобладает в эритроцитах половых железах, коре надпочечников, печени. Хотя окисление в гексозомонофосфатном цикле составляет всего 2% от обмена углеводов (при сахарном диабете может увеличиваться до 6%), для организма значение этого цикла очень велико. Особенность этого процесса – образование пентоз, накопление NADPH (2)-кофермента дегидрогеназ, участвующих в синтезе нуклеиновых кислот, холестерина, жирных кислот, активировании фолиевой кислоты и образовании АТФ. Гексозомонофосфатный цикл обеспечивает также процессы гидроксилирования, необходимые для синтеза биогенных аминов (катехоламины, серотонин) и стероидных гормонов коры надпочечников. Последовательная цепь реакций пентозного цикла (схема 7) приводит к образованию рибулозо-5-фосфата, который идет на построение нуклеотидов или серией обратных реакций преобразуется в гексозофосфаты с использованием их в гликолитическом цикле.
БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕКатаболизм органических веществ в тканях сопровождается потреблением кислорода и выделением СО2. Этот процесс называют тканевым дыханием. Кислород в этом процессе используется как акцептор водорода от окисляемых (дегидрируемых) веществ (субстратов), в результате чего синтезируется вода. Процесс окисления можно представить следующим уравнением: SH2 + 1/2 O2 à S + H2O. Окисляемые различные органические вещества (S - субстраты), представляют собой метаболиты катаболизма, их дегидрирование является экзоэргическим процессом. Энергия, освобождающаяся в ходе реакций окисления, либо полностью рассеивается в виде тепла, либо частично тратится на фосфорилирование ADP с образованием АТР. Организм превращает около 40% энергии, выделяющейся при окислении, в энергию макроэргических связей АТР. Большинство организмов в биосфере использует этот способ или очень сходный с ним (в качестве терминального акцептора водорода может быть не кислород, а другое соединение) как основной источник энергии, необходимый для синтеза внутриклеточной АТР. Таким путем клетка превращает химическую энергию питательных веществ, поступивших извне, в утилизируемую метаболическую энергию. Реакция дегидрирования и способ превращения выделившейся энергии путем синтеза АТР - это энергетически сопряженные реакции. Целиком весь сопряженный процесс называется окислительным фосфорилированием ADP:

Окислительное фосфорилирование ADP
Цепь транспорта электронов - ЦТЭ
Указанное выше уравнение для окислительно-восстановительной реакции представляет собой обобщенную форму, так как изображает процесс окисления субстратов как прямое дегидрирование, причем кислород выступает в роли непосредственного акцептора водорода. На самом деле кислород участвует в транспорте электронов иным образом. Существуют промежуточные переносчики при транспорте электронов от исходного донора электронов SH2 к терминальному акцептору - О2. Полный процесс представляет собой цепь последовательных окислительно-восстановительных реакций, в ходе которых происходит взаимодействие между переносчиками. Каждый промежуточный переносчик вначале выступает в роли акцептора электронов и протонов и из окисленного состояния переходит в восстановленную форму. Затем он передает электрон следующему переносчику и снова возвращается в окисленное состояние. На последней стадии переносчик передает электроны кислороду, который затем восстанавливается до воды. Совокупность последовательных окислительно-восстановительных реакций называется цепью переноса (транспорта) электронов, или дыхательной цепью:
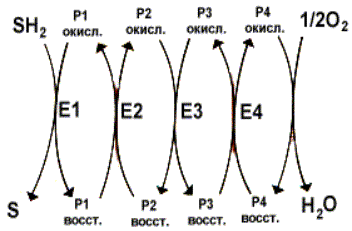
Перенос электронов и протонов с участием промежуточных переносчиков. SH2 - исходный донор протонов и электронов; P - промежуточные переносчики; E1, E2, E3, E4 - ферменты окислительно-восстановительных реакций
Промежуточными переносчиками в дыхательной цепи у высших организмов являются коферменты: NAD+ (никотинамид-адениндинуклеотид), FAD и FMN (флавинадениндинуклеотид и флавинмононуклеотид), кофермент Q (CoQ), семейство гемсодержащих белков - цитохромов (обозначаемых как цитохромы b, С1, С, А, А3) и белки, содержащие негеминовое железо. Все участники этой цепи разделены на четыре окислительно-восстановительные системы, связанные убихиноном (CoQ) и цитохромом С. Процесс начинается с переноса протонов и электронов от окисляемого субстрата на коферменты NAD+ или FAD. Это определяется тем, является ли дегидрогеназа, катализирующая первую стадию, NAD - зависимой или FAD - зависимой. Если процесс начинается с NAD+ , то следующим переносчиком будет FMN.
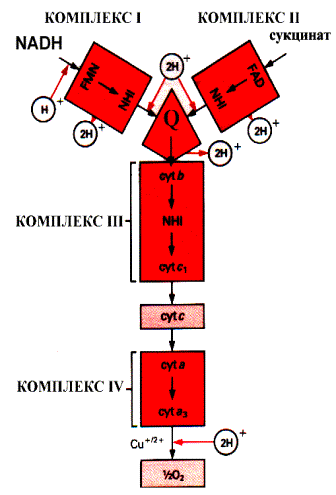
Последовательность промежуточных переносчиков протонов и электронов в дыхательной цепи
Тип участвующей дегидрогеназы зависит от природы субстрата. Но каким бы ни был исходный субстрат, электроны и протоны от флавинов переносятся к коферменту Q, а дальше пути электронов и протонов расходятся. Электроны с помощью системы цитохромов достигают кислорода, который затем, присоединяя протоны, превращается в воду. Чтобы разобраться в системе транспорта электронов, необходимо познакомиться с отдельными ее участниками. NAD - зависимая дегидрогеназа катализирует реакции окисления непосредственно субстрата (первичная дегидрогеназа). NAD+ является коферментом и выполняет роль акцептора водорода:
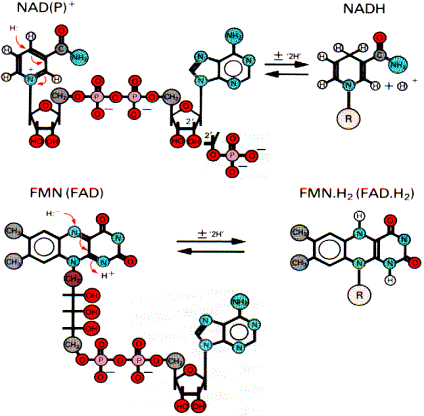
Коферменты дегидрогеназ
Символ 2Н+ означает два электрона и два протона, обычно переносимые в виде гидрид иона. В этом случае вместо терминов «донор электронов» и «акцептор электронов» иногда используют термины «донор или акцептор водорода». FAD - зависимая дегидрогеназа также выполняет функцию первичной дегидрогеназы. Коферментом является FAD, который является акцептором водорода от субстрата. NADH - дегидрогеназа катализирует окисление NADH и восстановление убихинона (CoQ). Переносчиком водорода является кофермент - FMN (комплекс 1). В процессе реакции водород сначала присоединяется к FMN, соединенному с ферментом, а затем передается на убихинон. Флавиновые коферменты (FAD и FMN) прочно связаны с ферментом как простетические группы, поэтому ферменты, в состав которых они входят, называются флавопротеины. Флавинмононуклеотид (FMN), или рибофлавин фосфат, неразрывно связан с белковой частью фермента. Строго говоря, FMN не является нуклеотидом, так как флавиновая часть связана с рибитолом, а не с рибозой.
Убихинон (кофермент Q) - производное изопрена:
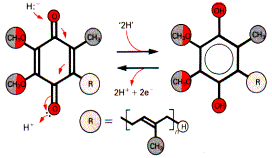
Название «убихинон» возникло из-за его повсеместной распространенности в природе. Кофермент Q действует как переносчик электронов на цитохромы.
Цитохромы - это гемопротеины - белки, содержащие в качестве прочно связанной простетической группы гем:
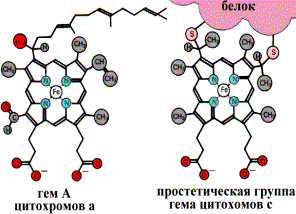
Простетическая группа гема в структуре цитохромов
Атом железа в геме может менять валентность, присоединяя или отдавая электроны:

В дыхательной цепи цитохромы служат переносчиками электронов и располагаются соответственно величине окислительно-восстановительного потенциала следующим образом: B, С1, С, а, а3. Гемовые группы цитохромов связаны с белковой частью донорно-акцепторными связями между ионом железа и соответствующими аминокислотными остатками:
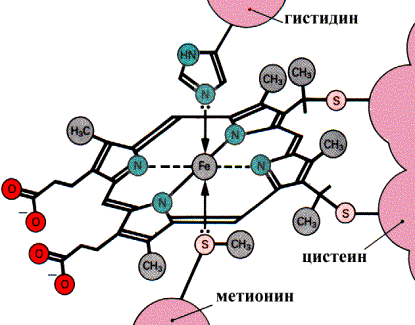
Связывание гема с белковой частью цитохрома С
В цитохромах С и С1 дополнительные ковалентные связи формируются между тиогруппами цистеина и боковыми винильными группами гема. QН2-дегидрогеназа (комплекс III) представляет собой комплекс цитохромов b и С1. Этот фермент катализирует окисление восстановленного кофермента Q и перенос электронов на цитохром С. Электроны последовательно переносятся атомами железа цитохромов b и С1, а затем поступают на цитохром С. Протоны после окисления QH2 освобождаются в раствор.
Цитохромоксидаза включает комплекс цитохромов а и а3 (комплекс IV). Цитохромоксидаза кроме гема содержит ионы меди, которые способны менять валентность и таким способом участвовать в переносе электронов:
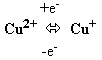
Цитохромоксидаза переносит электроны с цитохрома С на кислород. В переносе электронов участвуют сначала ионы железа цитохромов а и а3, а затем ион меди цитохрома а3. Молекула кислорода связывается с железом в геме цитохрома а3. Следовательно, переход электронов на кислород с иона меди цитохрома а3, происходит на молекуле фермента. Каждый из атомов молекулы кислорода присоединяет по два электрона и протона, образуя при этом молекулу воды.
Белки, содержащие негеминовое железо. Некоторое количество атомов железа в митохондриях связано не в геме цитохромов, а образует комплексы с другими белками. Эти белки называют также железосерными, так как атомы железа связаны с атомами серы цистеиновых остатков. Белки, содержащие негеминовое железо, участвуют в переносе электронов на нескольких стадиях, однако, не совсем ясны их локализация и механизм действия.
Окислительное фосфорилирование
Энергия, образующаяся при прохождении потока электронов по дыхательной цепи, используется для сопряженного фосфорилирования ADP. Эти два процесса взаимозависимы: окисление не может протекать в отсутствии ADP. Соотношение окисления и фосфорилирования определяется коэффициентом P/O (количество моль фосфорилированного ADP на 1/2 моль кислорода) коэффициент Р/О называется коэффициентом окислительного фосфорилирования и зависит от точки вхождения восстановительных эквивалентов в цепь транспорта электронов. Например Р/О=3, для субстратов, окисляемых NAD - зависимой дегидрогеназой , так как в дыхательной цепи есть три участка, где перенос электронов сопряжен с синтезом АТР. Не все субстраты передают электроны и протоны на NAD, некоторые окисляются FAD - зависимыми дегидрогеназами, которые переносят протоны и электроны сразу на убихинон, минуя первый комплекс. В этом случае Р/О=2. В действительности коэффициент фосфорилирования всегда меньше теоретической величины, потому что часть энергии, высвобождающейся при транспорте электронов, расходуется не на синтез АТР, а для переноса веществ через митохондриальную мембрану.
В сутки человек потребляет в среднем 27 моль кислорода. Основное его количество (примерно 25 моль) используется в митохондриях в дыхательной цепи. Следовательно, ежесуточно синтезируется 125 моль ATP или 62 кг (при расчете использовали коэффициент Р/О=2,5, то есть среднее значение коэффициента фосфорилирования). Масса всей АТР, содержащейся в организме, составляет примерно 20-30 г. Следовательно, можно сделать вывод, что каждая молекула АТР за сутки 2500 раз проходит процесс гидролиза и синтеза, что и характеризует интенсивность обмена АТР.
Сопряжение работы дыхательной цепи с процессом синтеза АТР
Существование такого сопряжения доказывается тем, что можно ингибировать образование АТР, не нарушая процесса транспорта электронов. Это достигается добавлением химических веществ, названных разобщителями. После удаления разобщителей синтез АТР восстанавливается. Изучение механизма сопряжении дает ответ на основные вопросы:
каким образом транспорт электронов служит источником энергии? как эта энергия передается в реакцию ADP + Pi a АТР?Существует несколько гипотез, объясняющих механизм сопряжения. Одной из них является хемоосмотическая теория. Цепь транспорта электронов функционирует как протонная (Н+)помпа, осуществляя перенос протонов из матрикса через внутреннюю мембрану в межмембранное пространство. Эндоэргический процесс выброса протонов из матрикса возможен за счет экзоэргических окислительно-восстановительных реакций дыхательной цепи. Перенос протонов приводит к возникновению разности концентрации Н+ с двух сторон митохондриальной мембраны: более высокая концентрация будет снаружи и более низкая - внутри. Митохондрия в результате переходит в «энергизованное» состояние, так как возникает градиент концентрации Н+ и одновременно разность электрических потенциалов со знаком плюс на наружной поверхности.
Электрохимический потенциал способен совершать «полезную» работу, он заставляет протоны двигаться в обратном направлении, но мембрана непроницаема для них кроме отдельных участков, называемых протонными каналами. Обратный перенос протонов в матрикс является экзоэргическим процессом, высвобождающаяся при этом энергия используется на фосфорилирование ADP. Эту реакцию катализирует фермент Н+-АТР-синтетаза, располагающаяся в области протонных каналов на внутренней поверхности внутренней мембраны.
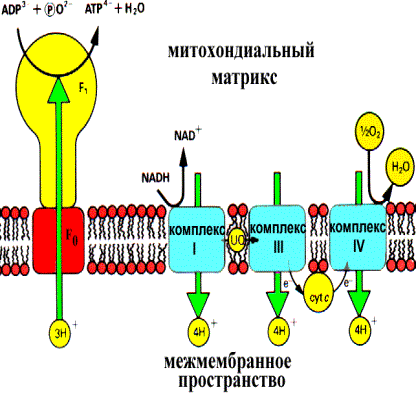
С опряжение цепи транспорта электронов и фосфорилирования ADP посредством протонного градиента
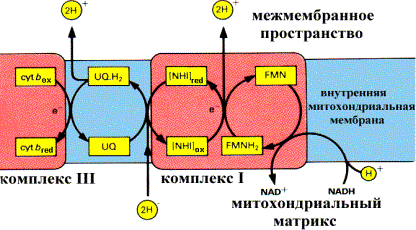
С труктура компонентов комплекса I, обеспечивающего функционирование «протонной помпы» при окислении NADH
Разобщение дыхания и фосфорилирования
Убедительные экспериментальные доказательства в пользу описанного механизма сопряжения дыхания и фосфорилирования были получены с помощью ионофоров. Молекулы этих веществ, как правило, липофильны и способны переносить ионы через мембрану. Например, 2,4-динитрофенол (протонофор) легко диффундирует через мембрану, в ионизированной и неионизированной форме, перенося протоны в сторону их меньшей концентрации в обход протонных каналов. Таким образом, 2,4-динитрофенол уничтожает электрохимический потенциал, и синтез АТР становится невозможным, хотя окисление субстратов при этом происходит. Энергия дыхательной цепи в этом случае полностью рассеивается в виде теплоты. Этим объясняется пирогенное действие разобщителей. Разобщающим действием обладают гормон щитовидной железы - тироксин, а также некоторые антибиотики, такие как валиномицин и грамицидин.
Дыхательный контроль
Скорость дыхания митохондрий может контролироваться концентрацией ADP. Это объясняется тем, что окисление и фосфорилирование жестко сопряжены. Энергия, необходимая клетке для совершения работы, поставляется за счет гидролиза АТР. Концентрация ADP при этом увеличивается; в результате создаются условия для ускорения дыхания, что и ведет к восполнению запасов АТР.
Ингибиторы цепи транспорта электронов и окислительного фосфорилирования
Ингибиторы, блокирующие дыхательную цепь, действуют в определенных местах, препятствуя работе дыхательных ферментов (KCN, барбитураты, ротенон). Существуют также вещества, ингибирующие окислительное фосфорилирование.
Похожие работы
... . Динамика химических превращений, происходящих в клетках, изучается биологической химией. Задачей физиологии является определение общих затрат веществ и энергии организмом и того, как они должны восполняться с помощью полноценного питания. Энергетический обмен служит показателем общего состояния и физиологической активности организма. Единица измерения энергии, обычно применяемая в биологии и ...
... веществ, осмотической и электрической работы. Основным источником энергии для осуществления в организме процессов жизнедеятельности является биологическое окисление питательных веществ. На это окисление расходуется кислород. Обмен веществ и энергии при различных уровнях функциональной активности организма Основной обмен. Под основным обменом (ОО) понимают минимальный уровень энергозатрат, ...
... повышен, при гипофункции этой железы он понижен. Основной обмен понижен у лиц, истощенных в результате продолжительного голодания или инфекционных болезней. Обмен веществ и энергии при деятельности организма. Наиболее резкое влияние на обмен веществ организма оказывает мышечная работа. Размер энерготрат при этом может повыситься в несколько раз. Либих считал, что мышечная работа совершается за ...
... организации жизни является биосферный, на котором изучают закономерности, характерные для всего живого, круговорот веществ и превращение энергии на Земле. Основы цитологии Химическая организация клетки Большинство живых организмов имеют клеточное строение. Клетка является структурной и функциональной единицей живого. Для нее характерны все признаки и функции живых организмов: обмен ...
0 комментариев