МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ
РЕСПУБЛИКИ КАЗАХСТАН
КАФЕДРА БИОЛОГИИ
КОНТРОЛЬНАЯ РАБОТА
«Физиология растений»
Выполнила ст-ка гр.ЗБХ-41
КОТ Т.Р.
номер з/к 0009322
Проверил доцент
Пономарева Т.М.
г.Павлодар, 2003г.
План
1. Теория дыхания Палладина.
2. Перекисная теория Баха.
3. Место физиологии растений в биологии.
4. Схема связи физиологии растений с другими биологическими науками.
5. Пути влияния недостатков воды на рост побегов.
Теория дыхания В. И. Палладина. На основании многочисленных опытов В. И. Палладии еще в 1916 г. дал следующую схему процесса дыхания. По Палладину, окисление сахара при дыхании идет не за счет непосредственного окисления его кислородом воздуха, а через цепь сложных превращений.
Под влиянием окислительно-восстановительных ферментов особые вещества, которые Палладии назвал дыхательными пигментами, связывают водород воды, а кислород воды производит окисление сахара. Этот первый этап дыхания Палладии изобразил следующей схемой:
С6Н12О6 + 12R + 6И2О = 12RH2 + 6СО2
Символом R Палладии обозначил дыхательный пигмент, который является акцептором водорода. Присоединяя водород, дыхательный пигмент переходит в бесцветное соединение — дыхательный хромоген. Затем уже кислород воздуха производит окисление дыхательного хромогена в пигмент:
12RH2+602 = 12H20+12R
Таким образом, по теории Палладина, кислород воды производит окисление сахара, а кислород воздуха окисляет акцептор (дыхательный хромоген).
Выдвинутое В. И. Палладиным положение, что кислород воздуха не окисляет непосредственно дыхательный субстрат, а окисляет лишь водород воды, было экспериментально подтверждено А. Л. Курсановым и Б. Б. Вартапетяном (1955). Применяя тяжелокислородную воду (Н2О18) и меченый кислород (О218), им удалось показать, что поглощенный при дыхании молекулярный кислород непосредственно не присоединяется к углероду субстрата дыхания, а идет на биосинтез воды в тканях растения. Если растению давалась вода, меченая по кислороду (Н2О18), то выделившаяся углекислота содержала О18, а в случае использования О28 углекислота не содержала меченого кислорода.
А. И. Опарин (1921) обнаружил роль хлорогеновой кислоты как дыхательного фермента.
Значение работ В. И. Палладина состоит в том, что он: 1) выдвинул положение о дыхании как о сложном окислительно-восстановительном процессе; 2) отметил роль воды в дыхании; 3) установил, что углекислота, выделяющаяся при дыхании, анаэробного происхождения.
В настоящее время мы знаем, что перенос водорода дыхательного материала к окисляющему его кислороду очень сложен. Встрече водорода с кислородом предшествуют процессы перемещения водорода, передача электронов и активация кислорода.
Теория медленного окисления А. Н. Баха. Теория медленного окисления была сформулирована крупным 'русским ученым А. Н. Бахом (1896).
Как известно, кислород воздуха не является активным окислителем, так как обе валентности кислорода взаимно насыщают друг друга: О=О. Для того чтобы кислород мог быть активным окислителем, он должен быть сперва активизирован, т. е. должна получиться молекула с подобного рода связями: — О — О — .
В основе теории Баха лежит понятие о пероксидах — перекисеобразных соединениях, образующихся присоединением кислорода к самоокисляющимся веществам А:
В этом случае кислород может частично или полностью отщепляться в виде атомов с ненасыщенными валентностями и поэтому легко окисляет соединения, непосредственно не окисляющиеся.
По Баху, окислительные ферменты оксидазы состоят из самоокисляющихся веществ, которые, присоединяя кислород воздуха, образуют перекисеобразные соединения. Сами по себе они не окисляют полифенолы, но под действием ферментов пероксидаз отщепляют от себя активный атомарный кислород.
Пероксидазы, как известно, могут разлагать перекись водорода Н2О2 на воду и активный атомарный кислород:
2Н2О = 2НО + 2О '
Выделенный кислород уже окисляет соответственное вещество. Таким образом, пероксидазы, используя частично уже активированный (перекисный) кислород, осуществляют окисление дыхательного материала.
О том, что активация водорода также имеет большое значение, говорят опыты с таким акцептором водорода, каким является краситель метиленовый синий. Восстановленный метиленовый синий переходит в лейкосоединение, т. е. становится бесцветным. В бесцветном метиленовом синем имеется на два атома водорода больше, чем у нормального соединения. Можно сделать такой опыт. Взять семена гороха, плотно набить ими небольшую стеклянную банку, налить туда водный раствор метиленового синего и затем закрыть крышку. Раствор метиленового синего быстро обесцветится. Если затем вынуть семена из банки, то на воздухе они начинают синеть, так как происходит окисление метиленового синего кислородом воздуха. Иными словами, в анаэробных условиях семена восстановили краситель метиленовый синий за счет водорода воды. Сейчас можно считать доказанным наличие процесса активации водорода и кислорода при дыхании!
Место физиологии растений в системе биологических дисциплин.
Физиология растений относится к биологическим, теоретическим наукам, является отраслью экспериментальной ботаники, которая в XIX в. выделилась в самостоятельную науку. В разное время на базе физиологии растений сформировались вирусология (1902 г.), агрохимия (1910 г.), химия гербицидов и стимуляторов роста (1925 г.), микробиология (1930 г.), биохимия (1930 г.). Физиология растений тесно связана с биохимией, биофизикой, микробиологией, цитологией, генетикой, молекулярной биологией, химией, физикой, использует современные методы химии, физики, математики, кибернетики. Успешное развитие биохимии способствует изучению обмена веществ и энергии растений на субклеточном и молекулярном уровнях. Трудно установить границы между отдельными биологическими науками, науками о жизни. Однако прежде всего физиология растений обеспечивает необходимую интеграцию всех биологических значений на уровне целого растения и ценоза, в этом ее особая роль в системе биологических наук.
Физиология растений как фундаментальная основа агрономических наук. К. А. Тимирязев неоднократно указывал, что физиология растений является теоретической основой рационального земледелия. Изучая основные закономерности жизнедеятельности растений, раскрывая зависимость функций растений от условий внешней среды, физиология растений является фундаментальной основой всех агрономических наук (земледелие, растениеводство, овощеводство и др.), создает теоретическую основу агротехнических систем, направленных на повышение урожайности и качества продукции сельскохозяйственных культур. Фундаментальную основу современного научного земледелия и агротехники сельскохозяйственных культур составляют результаты исследований и рекомендации в области:
теории фотосинтетической продуктивности посевов, разработки методов повышения использования растениями солнечной энергии, позволяющих довести использование ФАР до 3—5 % (вместо 0,5—1,5 %);
разработки физиологических основ и способов применения минеральных удобрений под сельскохозяйственные культуры, позволяющие более эффективно использовать минеральные удобрения без вредных экологических последствий;
раскрытия механизмов и повышения уровня биологической фиксаций азота атмосферы бобовыми растениями;
выяснения составляющих водного баланса растений и разработки приемов более продуктивного использования осадков, оросительной воды, внедрения капельного и импульсного орошения, автоматизированных оросительных систем;
раскрытия природы механизмов устойчивости растений к неблагоприятным факторам внешней среды, приемов, позволяющих растению в экстремальных условиях не только выжить, но и обеспечить достаточно высокую продуктивность (приемы повышения морозоустойчивости, холодостойкости, солеустойчивости и др.);
физиологии иммунитета растений, механизмов и условий, повышающих устойчивость сельскохозяйственных растений к болезням и вредителям;
познания регуляторных систем и механизмов, обеспечивающих упорядоченность и регуляцию физиологических процессов, способность растений к адаптации в широком диапазоне меняющихся условий среды;
использования фитогормонов и синтетических регуляторов роста, позволяющих направленно влиять на ход формирования урожая и его качество в технологиях сельскохозяйственных культур;
использования специальных методов и технических средств диагностики функционального состояния растений в полевых условиях, в селекционном процессе, в закрытом грунте для успешного «диалога» с ним с целью оптимизации условий выращивания, борьбы с болезнями и вредителями, оценки засухо-, -морозо-, солеустойчивости (показатели газообмена, биоэлектрические потенциалы, градиенты температур, скорость водного тока и др.);
теоретических физиологических и биохимических основ хранения урожая, снижения его потерь с использованием инертных газов, полупроницаемых мембран, консервантов и др.;
изучения потребности оптимальных режимов и способов облучения отдельных видов и сортов растений в сооружениях защищенного грунта в овощеводстве, в селекционном процессе;
изучения процессов и механизмов распределения ассимилянтов в онтогенезе растений в целях направленного формирования урожаев.
Как фундаментальная область знаний физиология растений служит также теоретической основой биотехнологии и биоинженерии растений.

















Способность растений переносить недостаточное влагообеспечение является комплексным свойством. Она определяется возможностью растений отсрочить опасное уменьшение оводненности протоплазмы (избегание высыхания) и способностью протоплазмы переносить обезвоживание без повреждения (выносливостью). Избегание высыхания достигается благодаря морфологической, анатомической приспособленности растений к сохранению оптимальной оводненности тканей при сухости воздуха и почвы.
Протоплазма большинства растений чрезвычайно чувствительна к обезвоживанию. Клетки надземных органов мезофитов, к которым относится большинство сельскохозяйственных культур, погибают, если их несколько часов выдерживать в воздухе с относительной влажностью 92—96 %, что соответствует депрессии водного потенциала 5,5—11 МПа. Корни еще более чувствительны. Водный дефицит приводит к прогрессирующему обезвоживанию протоплазмы, что вызывает нарушение физиологических функций, их прекращение и повреждение протоплазматических структур. Снижение содержания воды в клетке ниже оптимального уровня, вызывающее нарушения метаболизма, называется водным стрессом.
Одним из показателей водного стресса является депрессия водного потенциала. Для типичной клетки листа мезофитов с водным потенциалом примерно 2 МПа установлены три степени водного стресса: мягкий стресс — снижение водного потенциала не более чем на 1 МПа; умеренный (средний) стресс — снижение водного потенциала на 1,2—1,5 МПа; суровый стресс — снижение водного потенциала более чем на 1,5 МПа. Мягкий стресс соответствует небольшой потере тургора, тогда как средний связан с подвяданием листьев, а суровый стресс — с завяданием растений.
Наибольшей чувствительностью к водному стрессу характеризуются ростовые процессы. Это объясняется, во-первых, тем, что в основе растяжения клеток лежат тургорные явления, а утрата тургора во время роста клеток ведет к мелкоклеточности, во-вторых, рост тесным образом связан с нуклеиновым и белковым обменом. Сложность этих процессов приводит к их большой уязвимости при всякого рода неблагоприятных воздействиях, в том числе и водном стрессе.
Влияние водного дефицита на метаболические процессы в значительной мере зависит от длительности его действия. При устойчивом завядании растений увеличивается скорость распада РНК, белков и одновременно возрастает количество небелковых азотсодержащих соединений. Влияние водного дефицита на углеводный обмен листа выражается вначале в снижении моно- и дисахаридов из-за снижения интенсивности фотосинтеза. Затем количество моносахаридов может возрастать в результате гидролиза полисахаридов. При длительном водном дефиците наблюдается уменьшение количества всех форм Сахаров. Детоксикация избытка образующегося при протеолизе аммиака происходит с участием органических кислот, количество которых возрастает в тканях при водном дефиците. Процессы восстановления идут успешно, если не повреждены при недостатке воды генетические системы клеток. Защита ДНК состоит в частичном выведении молекул из активного состояния с помощью ядерных белков и, возможно, с участием специальных стрессовых белков. Поэтому изменения количества ДНК обнаруживаются лишь при сильной длительной засухе.
Отрицательное действие водного стресса во время вегетации заключается в подавлении ростовых процессов как наиболее сложных и требующих координации между отдельными физиологическими функциями. Вследствие подавления роста сокращается площадь листьев, т. е. ассимилирующая поверхность, что и является основной причиной снижения урожаев при засухе.
Следует отметить, что рост листьев более чувствителен к водному дефициту, чем рост корней.
На протяжении многих лет считалось доказанным, что в пересыхающей почве вода доступна растениям до тех пор, пока содержание влаги в ней не достигнет коэффициента устойчивого завядания, когда в почве остается недоступная растению вода. Согласно этой точке зрения физиологические процессы, рост и развитие растений на почве, подвергающейся иссушению, протекают нормально до достижения коэффициента завядания. Однако накоплено много данных, показывающих, что на обмен веществ, а следовательно, на рост и развитие растений влияет даже слабый водный дефицит.
Растения, перенесшие только однократную сильную кратковременную засуху, так и не возвращаются к нормальному обмену веществ. Внутренний водный баланс растения зависит от комплекса факторов, связанных а) с самим растением (засухоустойчивость, глубина проникновения и ветвление корней, фаза развития); б) с количеством растений на данной площади; в) , с климатическими факторами (потери воды на испарение и транспирацию, температура и влажность воздуха, туман, ветер и свет, количество осадков и т. д.); г) с почвенными факторами (количество воды в почве, осмотическое давление почвенного раствора, структура и влагоемкость почвы и др.).
Дефицит влаги в растениях действует на такие процессы, как поглощение воды, корневое давление, прорастание семян, устричные движения, транспирация, фотосинтез, дыхание, ферментативная активность растений, рост и развитие, соотношение минеральных веществ и др. Изменяя обмен веществ, недостаток воды влияет на продуктивность, вкус плодов, плотность древесины, длину и прочность волокна у хлопчатника и т. д. Степень оводненности, необходимая для начала прорастания семян, сильно варьирует у разных видов. Между скоростью прорастания семян и скоростью поглощения воды существует определенная корреляция. На скорость прорастания влияют также свойства семян и водоудерживающая способность почвы.
Влияние водного дефицита на метаболические процессы в значительной мере зависит от длительности его действия. При устойчивом завядании растений увеличивается скорость распада РНК, белков и одновременно возрастает количество небелковых азотсодержащих соединений и отток их в стебель и колос (у злаков). В результате в условиях засухи содержание белков в листьях относительно уменьшается, а в семенах - увеличивается.
Влияние водного дефицита на углеводный обмен выражается вначале в снижении содержания моно- и дисахаридов в фотосинтезирующих листьях из-за снижения интенсивности фотосинтеза, затем количество моносахаридов может возрастать как следствие гидролиза полисахаридов листьев нижних ярусов. При длительном водном дефиците наблюдается уменьшение количества всех форм Сахаров.
Длительный водный дефицит снижает интенсивность фотосинтеза и, как следствие, уменьшает образование АТР в процессе фотосинтетического фосфорилирования. Под влиянием почвенной и атмосферной засухи тормозится также отток продуктов фотосинтеза из листьев в другие органы.
В условиях оптимального водоснабжения наблюдается положительная корреляция между интенсивностью дыхания и количеством фосфорилированных продуктов. Водный дефицит по-разному сказывается на дыхании листьев разного возраста: в молодых листьях содержание фосфорилированных продуктов резко падает, как и интенсивность дыхания, а у листьев, закончивших рост, эта разница четко не проявляется. При дефиците воды снижается дыхательный коэффициент.
Необходимо отметить, что в условиях водного дефицита верхние листья, в которых за счет некоторого усиления гидролитических процессов увеличивается содержание осмотически активных веществ, оттягивают воду от нижних листьев и дольше сохраняют ненарушенными синтетические процессы, а нижние листья в этих условиях засыхают раньше верхних.
При длительном недостатке воды в тканях фаза растяжения заканчивается быстрее, что приводит к укорочению стебля и корня, к уменьшению размеров листьев, к их мелкоклеточности и т.д.
Похожие работы

... , в которых выращивают растения и проводят опыты в условиях определенного состава воздуха, нужной температуры и освещения. Применяя эти методы, физиологи исследуют растения на молекулярном, субклеточном, клеточном и организменном (интактное растение) уровнях. Сейчас в биологических исследованиях широко применяют электронные микроскопы просвечивающего типа с разрешающей способностью 0,15—0,5 нм, в ...
... или транспирация представляют совокупность химических и физических процессов. Чтобы понять механизм физиологического процесса, необходимо выделить его физические и химические компоненты. В связи с этим физиология растений все более нуждается в усовершенствованных методах биохимии. Биохимический подход оказался очень плодотворным в исследованиях таких сложных процессов, как фотосинтез и дыхание. ...
... насекомое тем временем усердно собирает сладкий нектар, предложенный цветком в награду за труды. Цветки правильной формы, то есть имеющие насколько осей симметрии, называются актиноморфными. В классе двудольных растений наблюдается богатое разнообразие цветковых форм. 3. Описание некоторых семейств класса двудольных Семейство лютиковых объединяет около 1500 видов трав и кустарников, многие ...
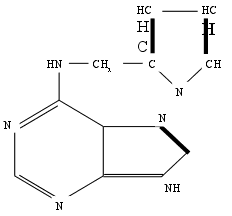
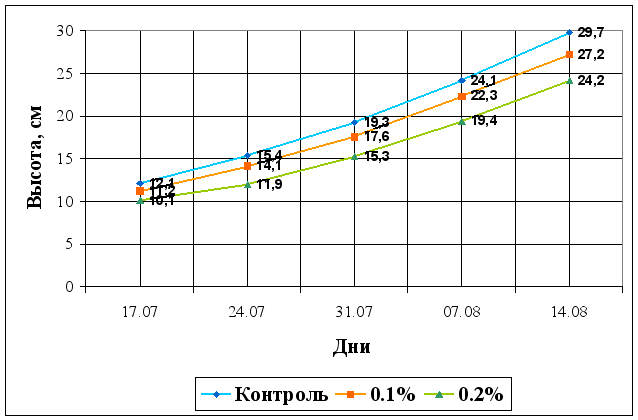
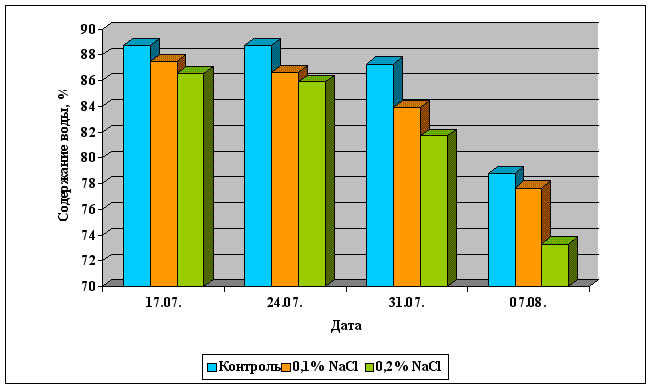
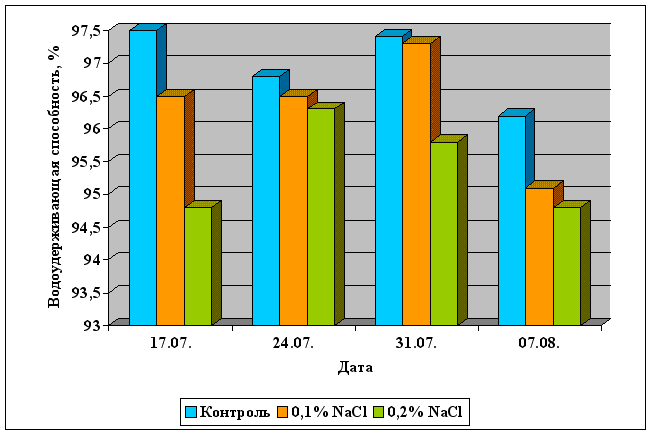
... Областного Университета в период с июня по сентябрь 2003 года. Объектом изучения являлись растения кукурузы сорта Россо. Изучение влияния 6-БАП на рост и биометрические показатели растений кукурузы при разном уровне засоления проводилось в условиях вегетационного опыта (почвенные культуры). Семена кукурузы проращивались при температуре 20оС в термостате в течение 3 дней, а затем высаживались в ...
0 комментариев