МИЧУРИНСКИЙ
ГОСУДАРСТВЕННЫЙ
ПЕДАГОГИЧЕСКИЙ
ИНСТИТУТ
Реферат по
физиологии
НА ТЕМУ:
физиология сенсорных систем
Выполнил:
5
курса
студент 53/6
группы
факультета
биологии
Болдырев
Е.А.
Преподаватель:
Микляева
М.А.
Мичуринск, 1999
ОглавлениеОглавление 2
Кожный анализатор 3
Значение кожного анализатора. 3
Периферический отдел кожного анализатора. 3
Пороги раздражения и пространственного различения. 5
Явления адаптации. 5
Проводящие пути кожного анализатора. 6
Корковый отдел кожного анализатора. 6
Взаимодействие различных видов кожной чувствительности. 8
Кожный анализатор как источник рефлекторных реакций. 10
Слуховой анализатор 11
Рецепция звуковых раздражении 11
Функция звукопроводящего аппарата уха. 12
Внутреннее ухо. 13
Резонансная теория слуха. 15
Проводящие пути слухового анализатора. 16
Корковый отдел слухового анализатора. 17
Факторы, определяющие чувствительность слухового анализатора. 18
Кожный анализатор Значение кожного анализатора.У животных, даже обладающих самой примитивной нервной системой, имеются рецепторы, расположенные на поверхности тела и чувствительные к внешним раздражениям. У кишечнополостных они мало специализированы и реагируют на действия различных раздражителей. В процессе эволюционного развития происходила дифференциация структуры функции этих рецепторов. Соответственно дифференцировался и центральный отдел анализатора. В результате первичный анализатор общей чувствительности поверхности тела дал начало ряду специализированных анализаторов: вкусовому, обонятельному, боковой линии (у рыб), вестибулярному, слуховому и зрительному. Функциональное обособление этих анализаторов не только не снизило значения рецепторной функции кожного покрова, но, наоборот, способствовало ее специализации в определенных направлениях. Так, у многих млекопитающих большое значение приобрела волосковая чувствительность (волоски на летательных перепонках летучих мышей, «усы» ночных хищников). У человека кожный анализатор играет существенную роль в познании внешнего мира. Через рецепторы кожи человек получает представление о плотности и упругости тел, их поверхности (гладкость, шершавость и пр.), температуре и т. д. У ребенка первые представления о форме предметов, об их величине и пространственном соотношении развиваются на основе совместной деятельности нескольких анализаторов, к числу которых наряду со зрительным, двигательным и другими относится и кожный. Не менее велико значение кожного анализатора как источника рефлекторных реакций, особенно оборонительных.
Периферический отдел кожного анализатора.Прилагая слабое электрическое раздражение к различным соседним точкам кожи, можно вызвать раздельные ощущения прикосновения, тепла, холода и боли (ощущение легкого укола). Тот же эффект дает раздражение кожи тонкой иглой или щетинкой. На основании этих наблюдений выделяют тактильные, тепловые, холодовые и болевые рецепторы кожи и соответственно четыре вида кожной чувствительности. Их раздельное существование подтверждается отсутствием определенных видов чувствительности в отдельных участках кожи и слизистых оболочек, а также частичной потерей чувствительности при воздействии на кожу некоторыми химическими веществами. Так, новокаин вызывает потерю чувствительности к холоду, а затем к боли и некоторое ее снижение к теплу, тогда как тактильная чувствительность не изменяется.
Каждый вид
кожной чувствительности
связан с определенными
структурными
особенностями
рецепторных
аппаратов (рис.
1). Тактильные
рецепторы,
воспринимающие
прикосновение
и давление,
имеют различное
строение. Одни
из них, оплетающие
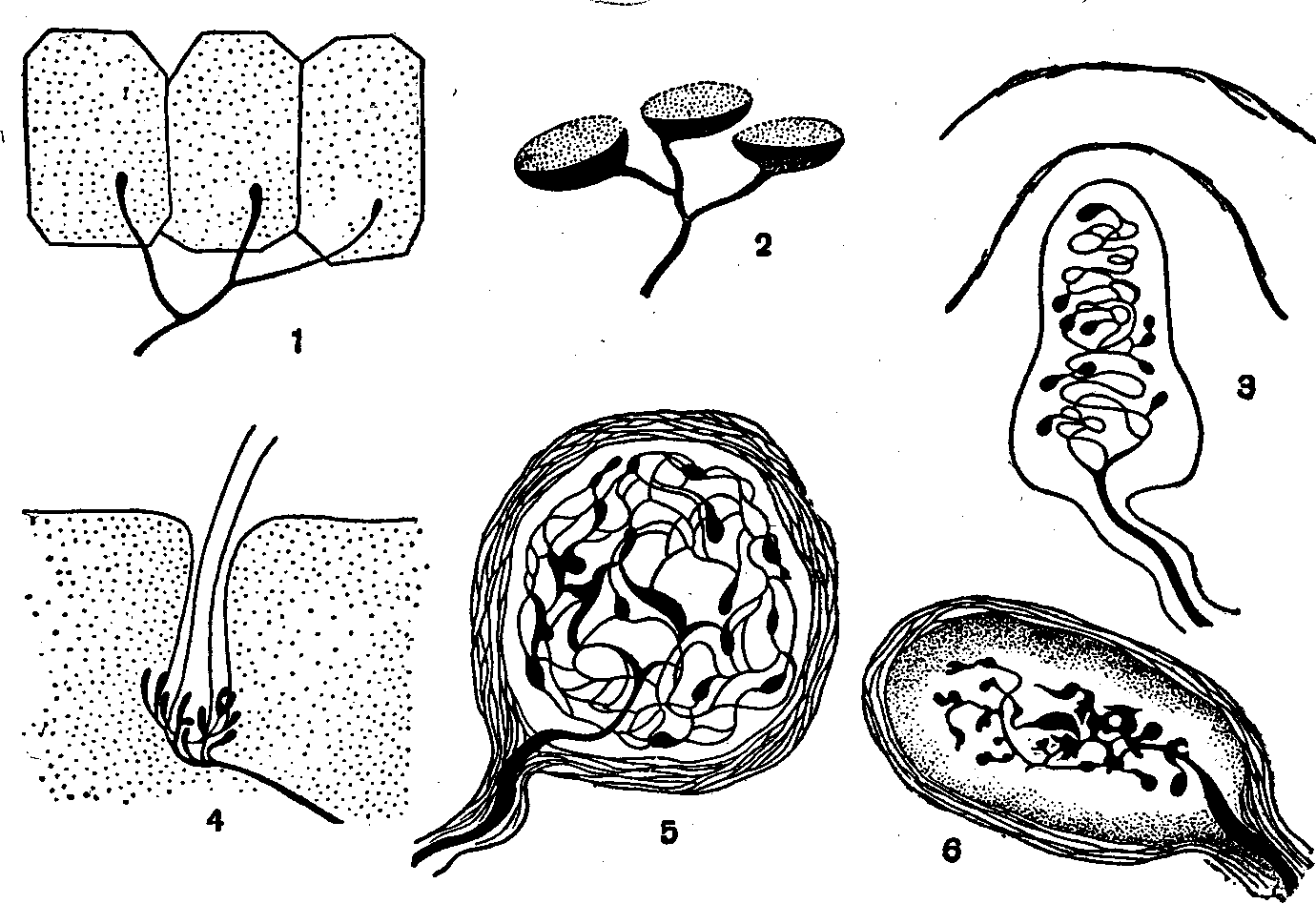
Рис. 1. Полусхематическое изображение некоторых рецепторных аппаратов кожи:
1 — свободные нервные окончания в эпидермисе; 2 — осязательные тельца Меркеля (главным образом в эпидермисе): 3 — осязательное тельце Мейсснера; 4 — нервное сплетение вокруг волосяной луковицы; 5 — чувствительное к холоду тельце (или колба) Краузе; 6 — фатерпачиниево тельце, чувствительное к давлению.
волосяную луковицу, обладают большой чувствительностью к малейшим колебаниям волоса. Другие обычно расположенные под самым эпидермисом в сосочках кожи, встречаются там, где отсутствуют волосы, а также в слизистой оболочке кончика языка. Особые рецепторы, которые находятся главным образом в подкожной клетчатке. возбуждаются не при легком прикосновении к коже, а при надавливании, -на. нее.
/ 1s'eцenmopы тепловые и холодовые (т. е. температурной чувствительности) расположены на различной глубине в собственно коже) и в верхних слоях подкожной клетчатки. Опытами с введением в кожу термоэлектрической иглы на различную глубину установлено, что холодовые рецепторы находятся ближе к поверхности, а тепловые в более глубоких слоях кожи.
Свободные нервные окончания в эпидермисе и в собственно коже, по-видимому, осуществляют болевую рецепцию. Однако достаточно сильное раздражение других рецепторов также может вызвать болевое ощущение.
Тщательное исследование кожи путем точечного раздражения отдельных ее пунктов выявило неодинаковое распределение различных видов чувствительности. В среднем, не считая волосистой части головы, приходится: болевых точек около 50 на 1 кв. см, а на всей поверхности кожи около миллиона; тактильных примерно вдвое меньше; холодовых в 4 раза меньше (12 на 1 кв. см); тепловых 1—2 на 1 кв. см или всего около 25 000. В коже различных областей тела количество и соотношение точек разных видов чувствительности сильно варьирует. На 1 кв. см кожи головы или ладонной стороны ногтевых фаланг кисти приходится более 100 тактильных точек, а на таком же участке кожи голени всего лишь 9—10. В то же время кожа лица содержит тепловых точек в 2—3 раза больше, а холодовых несколько меньше, чем кожа ногтевых фаланг.
Пороги раздражения и пространственного различения.Различное в отдельных областях кожи количество чувствительных точек, а также особенности структуры и глубины залегания соответствующих рецепторов обусловливают неодинаковые пороги адекватного
раздражения разных участков кожи. Так, порог тактильного раздражения для кожи кончика носа или ладонной стороны ногтевой фаланги пальцев руки раз в 10—15 меньше, чем для кожи живота и спины. Относительно редкое расположение холодовых и особенно тепловых точек приводит к тому, что температурное раздражение очень ограниченного участка кожи может совсем не вызвать соответствующего ощущения. В области голени, бедра и живота ощущение тепла иногда отсутствует даже в том случае, если площадь раздражаемого участка кожи превышает 1 кв. см.
Порог раздражения кожного анализатора зависит также от величины раздражаемой поверхности. Интенсивность температурного
•ощущения тем выше, чем большая поверхность кожи (например, руки) погружена в холодную или теплую воду.
Если одновременно прикасаться к двум соседним точкам кожи, то в зависимости от расстояния между ними можно получить ощущение либо одного, либо двух раздельных прикосновений. По наименьшему расстоянию между раздражаемыми точками кожи, при котором еще ощущается двойное прикосновение, можно судить о пороге пространственного различения тактильных раздражении (дискриминационная, 1Гли различительная, чувствительность). Для определения этого порога пользуются специальным прибором — эстезиометром. Он состоит из двух ножек с очень тонким, но. слегка притупленным концом; одна из них неподвижна, а другая передвигается вдоль разлинованной шкалы и может быть закреплена на любом расстоянии от первой. Исследования показали, что порог пространственного различения примерно равен: на кончике языка 1 мм, на ладонной стороне ногтевой фаланги пальцев руки 2 мм, на кончике носа 6—7 мм, на лбу 20—25 мм, на плече и предплечье 25—40 мм, на спине и груди 40—70 мм.
Явления адаптации.Кожные рецепторы проявляют резко выраженную адаптацию к непрерывно длящимся раздражениям. Надавливание на кожу или прикосновение к ней вызывает появление в соответствующих рецепторах и афферентных волокнах потенциалов действия, следующих друг за другом с большой частотой. Однако почти тотчас же потенциалы начинают становиться все более редкими, а через несколько секунд совсем исчезают. Соответственно 'прекращается и ощущение давления или прикосновения.
Относительную адаптацию к теплу и холоду легко обнаружить, 1 если одну руку опустить в воду, охлажденную до 15°, а другую — в воду, нагретую до 45°: при последующем переносе обеих рук в воду, имеющую температуру 30°, получается ясное ощущение, что одна рука находится в теплой воде, а другая — в холодной. Опыты показывают, что температурные ощущения зависят не только от абсолютной температуря раздражителя, но также и от состояния кожи, т. е. ее температуры и адаптации к теплу или холоду.
Проводящие пути кожного анализатора.Афферентные волокна, связанные с различными видами кожной чувствительности, отличаются друг от друга толщиной миэлиновой оболочки, а следовательно, и скоростью проведения импульсов. По волокнам безмякотным и обладающим тонкой миэлиновой оболочкой поступают импульсы, связанные с температурной, болевой, а отчасти и тактильной чувствительностью. Волокна второго нейрона проводящих путей перекрещиваются, и направляются к ядрам багров промежуточного мозга, где находятся тела третьего нейрона, проводящего импульсы к коре больших полушарий.
Через боковые ответвления (коллатерали) нервных волокон и через промежуточные (вставочные) нейроны импульсы, идущие от кожных рецепторов, могут переходить в спинном мозгу на эфферентные пути. Часть волокон направляется к бугоркам четверохолмия, при участии которых осуществляются такие рефлекторные реакции на раздражение кожи, как поворот головы, движение глаз, настораживание и т. п.
Корковый отдел кожного анализатора.У человека ядро коркового отдела кожного анализатора расположено в постцентральной области коры больших полушарий. Основная масса афферентных волокон направляется к полю 5, которое находится в глубине центральной борозды, а также к соседним полям 1 и 2. Уже в спинном мозгу пучки волокон, несущих импульсы от разных участков кожи, занимают строго определенное положение, а, дойдя до латерального ядра бугров промежуточного мозга, заканчиваются около различных клеточных групп, каждая из которых направляет свои аксоны к определенным участкам коры преимущественно противоположного полушария. В результате ядро коркового отдела кожного анализатора можно рассматривать как своеобразную проекцию кожи.
Исследование мозга людей, у которых при жизни отсутствовала чувствительность отдельных участков кожи, а также опыты с электрическим раздражением коры во время мозговых операций позволили установить корковую локализацию кожной чувствительности у человека. В верхнем участке постцентральной" извилины проецируется кожа нижних конечностей, а в среднем и нижнем участках — кожа верхних конечностей я головы. Величина корковой проекции различных областей кожи пропорциональна их богатству рецепторами, что совпадает и с функциональной значимостью отдельных участков кожного анализатора. Так, проекция кожи туловища занимает относительно малое Пространство; большой участок занимает проекция стопы, а еще больший — кисти. Особенно велики размеры проекции большого пальца и губ. Следует, однако, отметить, что участки коры, связанные с определенными областями кожи, в сильной степени перекрывают друг друга.
Повреждение Постцентральной извилины, и особенно поля 3, влечет за собой резкое расстройство анализа и синтеза кожных раздражении. Однако даже при полном разрушении постцентральной области коры (например, при удалении мозговой опухоли) происходит постепенное восстановление грубой чувствительности к давлению, теплу, холоду и боли. Это объясняется наличием в других участках коры рассеянных элементов кожного анализатора. Особенно широко рассеяны те элементы, раздражение которых вызывает болевые ощущения. Своеобразное нарушение наблюдается при поражениях верхней теменной области (поля 5 и 7): точно локализуя раздражаемый участок кожи, человек не может определить характер раздражения, а при ощупывании не различает знакомые предметы, т. е. не узнает их.
Анализ и синтез кожных раздражении. В опыте на спинномозговом препарате животного можно обнаружить элементарный пространственный анализ, проявляющийся в различных рефлекторных двигательных реакциях в зависимости от места раздражения. Наличие четырех основных видов кожной чувствительности, связанных с различными рецепторами, создает возможность качественного периферического анализа раздражении.
В известной мере анализ, а также и синтез может продолжаться и .в проводящих путях, особенно в соответствующих ядрах бугров промежуточного мозга. Здесь взаимодействие импульсов, идущих от разных участков кожи и от разных ее рецепторов, может определять как ответную реакцию, осуществляемую через подкорковые двигательные ядра, так и судьбу тех импульсов, которые направляются в кору больших полушарий. В буграх же происходит взаимодействие импульсов, относящихся к разным анализаторам, что, по-видимому, служит источником тех неясных, смутных ощущений общего состояния организма, которые принято называть низшими эмоциями.
Высший анализ и синтез раздражении, связанный с образованием положительных и отрицательных условных рефлексов, происходит в корковом отделе анализатора. Здесь возникают связи между определенными комбинациями очагов возбуждения в кожном анализаторе и теми очагами, которые появляются в коре под влиянием раздражении, падающих на другие анализаторы. Опыты на собаках показали, что можно выработать условные рефлексы на самые различные тактильные, температурные и болевые раздражения, сочетая их с действием тех или иных безусловных раздражителей. Очень сходные раздражения, как правило, легко могут быть отдифференцированы. Так, удается выработать дифференцировку на тактильные раздражители, приложенные к двум соседним участкам кожи, а также на различную силу или частоту раздражении.
Тонкий анализ и синтез раздражении приобретает особое значение в тех случаях, когда выбывают из строя другие анализаторы. Высокого совершенства аналитико-синтетическая деятельность кожного анализатора достигает у людей, потерявших зрение и слух. Примером может служить слепоглухонемая Ольга Скороходова, опубликовавшая в 1947 году книгу под названием «Как я воспринимаю окружающий мир». Малейшие колебания температуры, еле заметное движение воздуха, ничтожные детали ощупываемого предмета — все это становилось источником определенных условных связей, позволяющих легко и быстро ориентироваться в окружающих явлениях.
Взаимодействие различных видов кожной чувствительности.Обычно раздражения, воспринимаемые кожей, носят комплексный характер, ибо действуют на разные виды рецепторов, причем получается единое обобщенное ощущение, характер которого зависит от степени вовлечения в реакцию различных видов рецепторов. Приложение к коже холодного или теплого предмета в первый момент вызывает отчетливое ощущение прикосновения, к которому быстро присоединяется и становится доминирующим ощущение холода или тепла. Объясняется это тем, что импульсы от тактильных рецепторов быстрее достигают центральной нервной системы, ибо проходят по толстым мякотным волокнам, тогда как от тепловых и холодовых рецепторов импульсы идут по безмякотным или тонким мякотным волокнам. Следовательно, прежде всего очаг возбуждения возникает в тех корковых клетках, которые получают импульсы от тактильных рецепторов. Последующее появление второго очага возбуждения — в клетках, получающих импульсы от тепловых или холодовых рецепторов,— вызывает в силу отрицательной индукции понижение возбудимости клеток, воспринимающих тактильное раздражение.
Сила очагов возбуждения, возникающих в коре под влиянием импульсов от тактильных и болевых рецепторов, зависит от характера и интенсивности раздражения, причем более сильный (доминирующий) очаг вследствие отрицательной индукции способен понижать возбудимость слабого очага. Этим объясняется ослабление или отсутствие тактильных ощущений при сильном болевом раздражении, или, наоборот, уменьшение болевых ощущений, если потирать рукой ушибленное место, прикладывать к нему холодный или теплый предмет. Болевые ощущения могут совсем отсутствовать, когда одновременно сильно раздражаются тактильные и относительно слабо болевые рецепторы. Аналогичный корковый механизм лежит в основе подавления болевых ощущений путем интенсивного сжимания рук; в этом случае сильный очаг возбуждения возникает не только в кожном, но и в двигательном анализаторе.
Значение коркового взаимодействия импульсов, приходящих от различных рецепторов кожи, особенно отчетливо выявляется при некоторых заболеваниях центральной нервной системы. Поражение бугров промежуточного мозга может резко нарушить тактильную чувствительность, тогда как болевая и температурная сохраняются. Однако болевые и температурные ощущения изменяются: они становятся очень резкими и плохо локализуются. Например, укол в области плеча воспринимается как резкое болевое раздражение всей или большей части конечности. Это объясняется тем, что возбуждение, возникающее в коре под влиянием болевого раздражения, легко иррадиирует, чему способствует отсутствие отрицательной индукции со стороны корковых центров тактильной чувствительности.
Сходные явления отмечал английский невропатолог Хэд. Он перерезал у себя веточку кожного нерва, тотчас же сшил ее концы и затем в продолжение длительного времени наблюдал за восстановлением кожной чувствительности по мере того, как происходила регенерация нервных волокон. Прежде всего начала восстанавливаться болевая и грубая температурная чувствительность. В этот период легкий угол большого пальца иглой и даже простое прикосновение к коже вызывали резкое и мучительное болевое ощущение. Приложение теплового раздражителя (+45° и выше), а также холодового (+10° и ниже) вызывало столь же резкие и очень неприятные температурные ощущения. Все эти ощущения носили разлитой характер: определить место приложения раздражителя не удавалось. Значительно позднее стала восстанавливаться чувствительность тактильная и тонкая температурная. Вскоре болевые ощущения приобрели обычный, нормальный характер, и опять появилась способность точно локализовать наносимые раздражения.
На основании своих наблюдений Хэд пришел к выводу о существовании двух видов чувствительности: филогенетически древней, примитивной и позднее появившейся тонкой. Однако этот вывод требует дальнейших подтверждений, так как некоторые факты ему противоречат. Описанные Хэдом наблюдения следует рассматривать как лишнее подтверждение коркового взаимодействия импульсов, приходящих от различных рецепторов кожи, а также значения тактильной чувствительности для точной локализации наносимых на кожу раздражении.
Факторы, определяющие чувствительность кожного анализатора. Существенное влияние на чувствительность кожного анализатора оказывают температура кожи и состояние кровообращения в ней (например, сужение или расширение кожных сосудов). Известно, что при повышении температуры кожи ее чувствительность к тактильному и болевому раздражениям повышается, а при охлаждении понижается. Изменение температуры влияет и на порог пространственного различения.
Чувствительность к теплу и холоду, далеко не одинаковая у отдельных людей, в сильной степени зависит от адаптации кожи к этим раздражителям. Как правило, она особенно велика при температуре кожи 28—30°. Наблюдения над чувствительностью кожи руки показали, что при этой температуре разностный порог ощущения нередко может достигать 0,1°. Чувствительность понижается к тепловым раздражителям при адаптации- кожи к низкой температуре, а к холодовым — при адаптации к высокой температуре.
Изменения кожной чувствительности зависят и от состояния центрального отдела анализатора. Во-первых, центральная нервная система, реагируя на поступающие с периферии импульсы, оказывает рефлекторные влияния на кожу: изменяет ее функциональное состояние, а тем самым и чувствительность. Во-вторых, меняется возбудимость корковых клеток кожного анализатора; она повышается, если афферентные импульсы достаточно интенсивны и особенно при образовании жизненно важных условных связей. Этим объясняется повышение кожной чувствительности под влиянием профессиональных навыков, а также при нарушении функции других анализаторов, что имеет место у слепых и у слепоглухонемых. Слабые тактильные и температурные раздражения, в особенности длительно и часто повторяющиеся, наоборот, понижают возбудимость корковых клеток и приводят к развитию в них процесса торможения. Методом условных рефлексов показано, что такие раздражения легко вызывают иррадиацию торможения.
Изменение возбудимости коркового отдела анализатора может происходить и под влиянием сдвигов функционального состояния корковых отделов других анализаторов. Установлено, что порог тактильных раздражении, а также порог их пространственного различения в условиях освещения ниже (т. е. чувствительность выше), чем в темноте. Порог тактильных раздражении понижается и в том случае, если усиливается приток импульсов с рецепторов двигательного анализатора (например, при нанесении болевого раздражения путем внутримышечной инъекции солевого раствора). Повышение кожной чувствительности при раздражении центрального отдела зрительного и двигательного анализаторов объясняется иррадиацией возбуждения на корковые клетки кожного анализатора.
Порог раздражения может, наоборот, повышаться вследствие отрицательной индукции, возникающей под влиянием сильного очага возбуждения в корковом отделе другого анализатора. Так, значительное мышечное напряжение резко повышает порог болевых и тактильных раздражении, т. е. понижает чувствительность к ним. Аналогичное влияние, особенно на порог пространственного различения тактильных раздражении, оказывает утомление. Надо полагать, что и в этом снижении кожной чувствительности существенную роль играют корковые процессы.
Кожный анализатор как источник рефлекторных реакций.Рефлекторные реакции возникают при раздражении рецепторов любого анализатора. Так, вкусовые и запаховые раздражители вызывают рефлексы со стороны органов пищеварения; в ответ на звуковое или световое раздражение может появиться ориентировочный рефлекс и т. д. Не составляет исключения и кожный анализатор. Сосудистые, двигательные и другие рефлексы легко возникают в ответ на различные раздражения кожи. Особое значение приобретают рефлексы на болевое раздражение.
Сильное болевое ощущение возникает при действии на кожу любых раздражителей, если они достигают большой интенсивности и оказывают повреждающее действие. Иными словами, болевыми раздражителями могут оказаться самые разнообразные физические и химические агенты, будь то тепло или холод, механическое воздействие (например, давление или растяжение), химические вещества и т. д. Следовательно, для рецепции боли адекватна не природа раздражителя, а интенсивность его воздействия на кожу.
Если в результате образования соответствующих условных рефлексов звуковые, зрительные и другие раздражители могут быть сигналами предстоящего повреждения организма, то болевое раздражение сигнализирует уже наступившее повреждение. В ответ на такой сигнал возникает оборонительная безусловнорефлекторная реакция; она направлена на устранение раздражителя или на удаление от него.
Оборонительная реакция на повреждающее болевое раздражение кожи не ограничивается тем или иным ответным двигательным актом. Она проявляется в значительных изменениях функций различных органов. Еще в 70-х годах прошлого века Павлов обнаружил, что в условиях острого опыта болевое раздражение вызывает резкое торможение секреторной функции пищеварительных желез. В дальнейшем было установлено, что под влиянием боли наступают рефлекторные изменения кровообращения, повышается свертываемость крови и увеличивается содержание в ней адреналина и сахара, нарушается функция почек и т. д. Иногда при сильном и внезапном раздражении наблюдаются остановка сердца и другие сильные изменения жизненно важных функций, в результате чего наступает гибель организма.
Таким образом, рефлекс на болевое раздражение представляет собой целостную реакцию всего организма. Характер этой реакции зависит как от состояния самого организма, так и от интенсивности повреждающего действия. Чаще всего болевое раздражение повышает возбудимость нервной системы и вызывает такие координированные реакции различных органов, которые облегчают протекание защитных функций организма.
Слуховой анализатор Рецепция звуковых раздражении
Орган слуха. У большинства беспозвоночных нет специальных тонорецепторов, чувствительных только к звуковым колебаниям. Однако у насекомых описаны специфические слуховые органы; они могут быть расположены в различных местах тела и состоят из тонкой натянутой перепонки, отделяющей наружный воздух от слуховой полости. С внутренней стороны перепонки находятся слуховые рецепторные клетки. При помощи этих органов некоторые насекомые могут воспринимать звуки очень большой частоты — до 40 и даже до 90 тысяч колебаний в секунду.
У низших позвоночных периферический слуховой орган вместе с вестибулярным аппаратом дифференцируется из переднего конца органа боковой линии, рецепторы которого воспринимают колебания водной среды. Ослепленная щука при условии сохранения органа боковой линии схватывает проплывающую мимо рыбу и передвигается, не натыкаясь на встречные предметы, которые отражают колебания воды, производимые движениями щуки. Колебания большей частоты воспринимаются только развившимся из переднего конца органа боковой линии мешочком и его слепым выростом, получившим название лагены (lagena). У амфибий (и особенно у рептилий) ближе к основанию лагены появляется особый слуховой участок— натянутая перепонка, состоящая из параллельно расположенных соединительнотканных волоконец. У млекопитающих за счет разрастания этого участка слепой вырост резко удлиняется. Изгибаясь, он принимает форму раковины улитки с различным у разных животных числом витков. Отсюда и название этого органа — улитка.
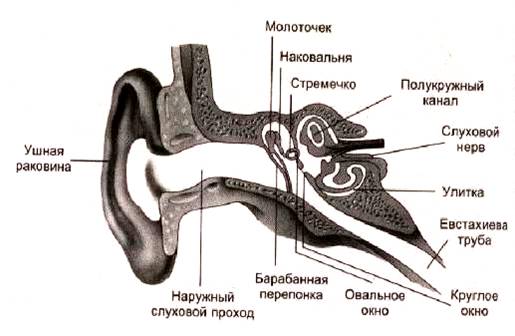
Ухо как периферический орган слухового анализатора состоит не только из рецепторного аппарата, скрытого в толще височной кости и образующего вместе с вестибулярным аппаратом так называемое внутреннее ухо. Существенное значение имеют те части уха, которые связаны с улавливанием звуков и их проведением к рецепторному аппарату.
Звукопроводящий аппарат всех наземных животных — это среднее ухо, или барабанная полость, которая образовалась за счет первой жаберной щели. Уже у рептилий в этой полости находится слуховая косточка, «облегчающая передачу звуковых колебаний. У млекопитающих имеются три сочлененные между собой косточки, способствующие увеличению силы звуковых колебаний. Звукоулавливающий аппарат, или наружное ухо, состоит из наружного слухового прохода и ушной раковины, которая впервые появляется у млекопитающих. У многих из них она подвижна, что позволяет направлять ее в сторону появления звуков и тем самым лучше их улавливать.
Функция звукопроводящего аппарата уха.Барабанная полость (рис. 1а) сообщается с наружным воздухом через особый канал — слуховую, или евстахиеву, трубу, наружное отверстие которой находится в стенке носоглотки. Обычно оно закрыто, но в момент глотания раскрывается. При резком изменении атмосферного давления, например при спуске в глубокую шахту, при подъеме или приземлении самолета, может возникнуть значительная разница между давлением наружного воздуха и давлением воздуха в барабанной полости, что вызывает неприятные ощущения, а иногда и повреждение барабанной перепонки. Раскрытие отверстия слуховой трубы способствует выравниванию давления, а потому при

Рис. 1. Полусхематическое изображение среднего уха:
/— наружный слуховой проход', 2— барабанная полость; 3 — слуховая труба; 4 — барабанная перепонка; 5 — молоточек; 6 — наковальня; 7 — стремя; 8 — окно преддверия (овальное); Я — окно улитки (круглое); 10— костная ткань.
Внутри барабанной полости находятся три слуховые косточки — молоточек, наковальня и стремя, соединенные между собой суставами. Среднее ухо отделено от наружного барабанной перепонкой, а от внутреннего — костной перегородкой с двумя отверстиями. Одно из них называется овальным окном или окном преддверия. К его краям при помощи эластичной кольцевой связки прикреплено основание стремени, Другое отверстие — круглое окно, или окно улитки,— затянуто тонкой соединительнотканной мембраной.Воздушные звуковые волны, попадая в слуховой проход, вызывают колебания барабанной перепонки, которые через систему слуховых косточек, а также через воздух, находящийся в среднем ухе, передаются перилимфе внутреннего уха. Сочлененные между собой слуховые косточки можно рассматривать как рычаг первого рода, длинное плечо которого соединено с барабанной перепонкой, а короткое укреплено в овальном окне. При передаче движения с длинного на короткое плечо происходит /уменьшение размаха (амплитуды) за счет увеличения развиваемой силы. Значительное увеличение силы звуковых колебаний происходит еще и потому, что поверхность основания стремени во много раз меньше поверхности барабанной перепонки. В целом сила звуковых колебаний увеличивается по крайней мере в 30—40 раз.
При мощных звуках вследствие сокращения мышц барабанной полости увеличивается напряжение барабанной перепонки и уменьшается подвижность основания стремени, что ведет к понижению силы передаваемых колебаний.
Полное удаление барабанной перепонки лишь снижает слух, но не ведет к его потере. Это объясняется тем, что существенную роль в передаче звуковых колебаний играет мембрана круглого окна, которая воспринимает колебания воздуха, находящегося в полости среднего уха.
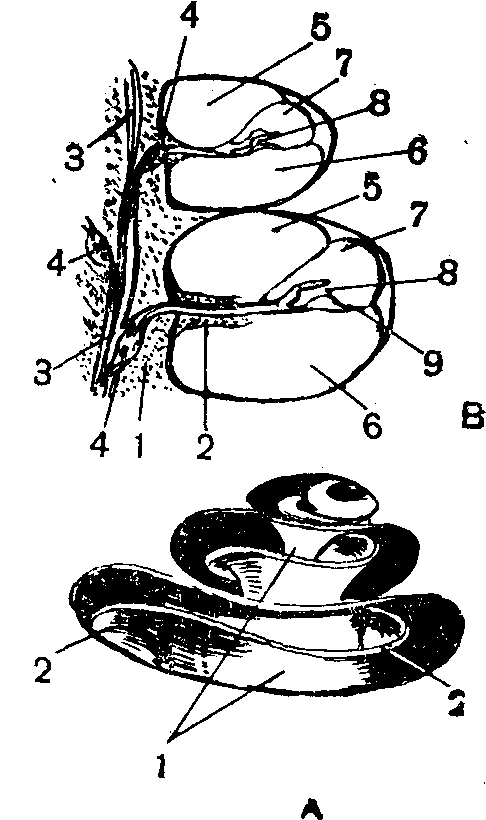
Рис. 3. Схематическое изображение улитки внутреннего уха:
А — костный канал улитки;
В — схема поперечного разреза части улитки; — костный стержень;2 — спиральная костная пластинка; 3 — волокна улиткового нерва;
4 — скопление тел первого нейрона слухового проводящего пути; 5 — лестница преддверия; S—лестница барабана; 7— улитковая часть перепончатого лабиринта;
5 — кортиев орган; S — основная пластинка.
Внутреннее ухо.Внутреннее ухо представляет собой сложную систему каналов, находящихся в пирамиде височной кости и получивших название костного лабиринта. Расположенные в нем улитка и вестибулярный аппарат образуют перепончатый лабиринт (рис. 2). Пространство между стенками костного и перепончатого лабиринтов заполнено жидкостью — перилимфой. К слуховому анализатору относится только передняя часть перепончатого лабиринта, которая расположена внутри костного канала улитки и вместе с ним образует два с половиной оборота вокруг костного стержня (рис. 3). От костного стержня внутрь канала отходит отросток в виде винтообразной спиральной пластинки, широкой у основания улитки и постепенно суживающейся к ее вершине. Эта пластинка не доходит до противоположной, наружной стенки канала. Между пластин-

Рис. 2. Общая схема костного и находящегося в нем перепончатого лабиринта:
/ — кость; 2 — полость среднего уха; 3 —стремя;
4 — окно преддверия; 5— окно улитки; 6 — улитка; 7 и 8 — отолитовый аппарат (7 — саккулус или круглый мешочек; 8 — утрикулус, или овальный мешочек); 9, 10 и 11 — полукружные каналы 12 — пространство между костным и перепончатым лабиринтами, заполненное перилимфой.
кой и наружной стенкой расположена улитковая часть перепончатого лабиринта, вследствие чего весь канал оказывается разделенным на два этажа, или прохода. Один из них сообщается с преддверием костного лабиринта и называется лестницей преддверия, другой начинается от окна улитки, граничащего с барабанной полостью, и называется лестницей барабана. Оба прохода сообщаются только в верхнем, узком конце улитки.
На поперечном разрезе улитковая часть перепончатого лабиринта имеет форму вытянутого треугольника. Его нижняя сторона, граничащая с лестницей барабана, образована основной пластинкой, которая состоит из погруженных в гомогенную массу тончайших эластических соединительнотканных волокон, натянутых между свободным краем спиральной костной пластинки и наружной стенкой канала улитки. Верхняя сторона треугольника граничит с лестницей преддверия, отходя под острым углом от верхней поверхности
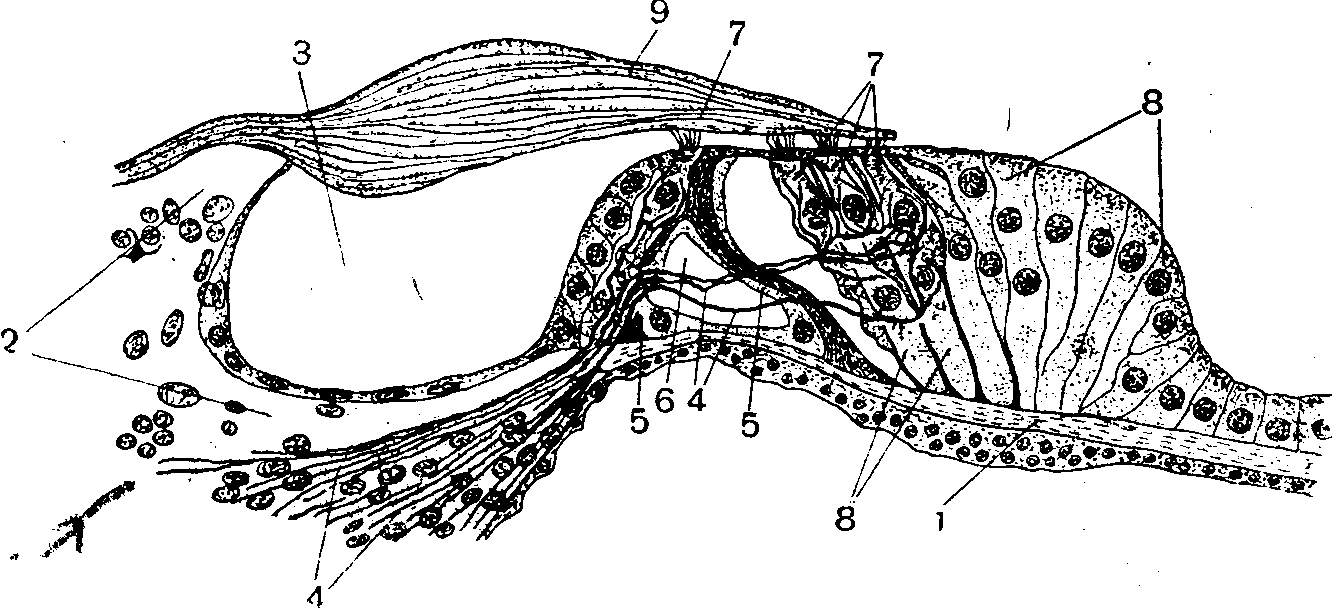
Рис. 4. Схема строения кортиева органа:
/ — основная пластинка; 2 — костная спиральная пластинка; 3 — спиральный канал;
4 — нервные волокна; S — столбовые клетки, образующие тоннель (6); 7 — слуховые, или волосковые, клетки; 8 — опорные клетки; 9 — покровная пластинка.
спиральной костной пластинки и направляясь, как и основная пластинка, к наружной стенке канала улитки. Третья, самая короткая сторона треугольника состоит из соединительной ткани, плотно сращенной с наружной стенкой костного канала. Функция кортиева органа. Рецепторный аппарат слухового анализатора, или спиральный кортиев орган, расположен внутри улитковой части перепончатого лабиринта на верхней поверхности основной пластинки (рис. 4). Вдоль внутренней части основной пластинки, на некотором расстоянии друг от друга, расположены два ряда столбовых клеток, которые, соприкасаясь своими верхними концами, отграничивают свободное треугольное пространство, или тоннель. По обе стороны от него находятся чувствительные к звуковым колебаниям см/ховые, или волосковые, клетки, каждая из которых на своей верхней свободной поверхности имеет 15—20 небольших тончайших волосков. Концы вслосков погружены в покровную пластинку, она укреплена на костной-спиральной пластинке и свободным концом покрывает кортиев орган. Волосковые клетки расположены кнутри от тоннеля в один ряд, а кнаружи—в три ряда. От основной пластинки они отделены опорными клетками.
К основаниям волосковых клеток подходят конечные разветвления волокон биполярных нервных клеток, тела которых расположены в центральном канале костного стержня улитки, где они образуют так называемый спиральный узел, гомологичный межпозвоночному узлу спинномозговых нервов. Каждая из трех с половиной тысяч внутренних волосковых клеток связана с одной, а иногда и с двумя отдельными нервными клетками. Наружные волосковые клетки, количество которых достигает 15—20 тысяч, могут быть соединены и с несколькими нервными клетками, но при этом каждое нервное волокно дает ответвления только к волоско-вым клеткам одного и того же ряда.
Перилимфа, окружающая перепончатый аппарат улитки, испытывает давление, которое и меняется соответственно частоте, силе и форме звуковых колебаний. Изменения давления вызывают колебания основной пластинки вместе с расположенными на ней клетками, волоски которых испытывают при этом изменения давления со стороны покровной пластинки. Это, по-видимому, и ведет к возникновению возбуждения в волосковых клетках, которое передается на конечные разветвления нервных волокон.
Резонансная теория слуха.Среди различных теорий, объясняющих механизм периферического анализа звуков, наиболее обоснованной следует считать резонансную теорию, предложенную Гельмгольцем в 1863 году. Если около открытого рояля воспроизвести музыкальным инструментом или голосом звук определенной высоты, то начнет резонировать, т. е. звучать в ответ, струна, настроенная на тот же самый тон. Изучая структурные особенности основной пластинки улитки, Гельмгольц пришел к выводу, что звуковые волны, приходящие из окружающей среды, вызывают колебания поперечных волокон пластинки по принципу резонанса.
Всего насчитывают в основной пластинке около 24 000 поперечных эластических волокон. Они различны по длине и степени натяжения: самые короткие и сильнее натянутые расположены у основания улитки; чем ближе к ее вершине, тем они длиннее и слабее натянуты. Согласно резонансной теории, различные участки основной пластинки реагируют колебанием своих волокон на звуки разной высоты. Такое представление подтвердилось опытами Л. А. Андреева. После выработки у собак условных рефлексов на чистые тоны различной высоты улитку одного уха он полностью удалял, а улитку другого подвергал частичному повреждению. В зависимости от того, какой участок кортиева органа второго уха был поврежден, наблюдалось исчезновение ранее выработанных положительных и отрицательных условных рефлексов на звуки определенной частоты колебаний. 'При разрушении кортиева органа ближе к основанию улитки исчезали условные рефлексы на высокие тоны. Чем ближе к верхушке локализовалось повреждение, тем ниже были тоны, утратившие значение условных раздражителей.
Проводящие пути слухового анализатора.Первый нейрон проводящих путей слухового анализатора — упомянутые выше биполярные клетки. Их аксоны образуют улитковый нерв,, волокна которого входят в продолговатый мозг и оканчиваются в ядрах, где расположены клетки второго нейрона проводящих путей. Аксоны клеток второго нейрона доходят до внутреннего коленчатого тела,
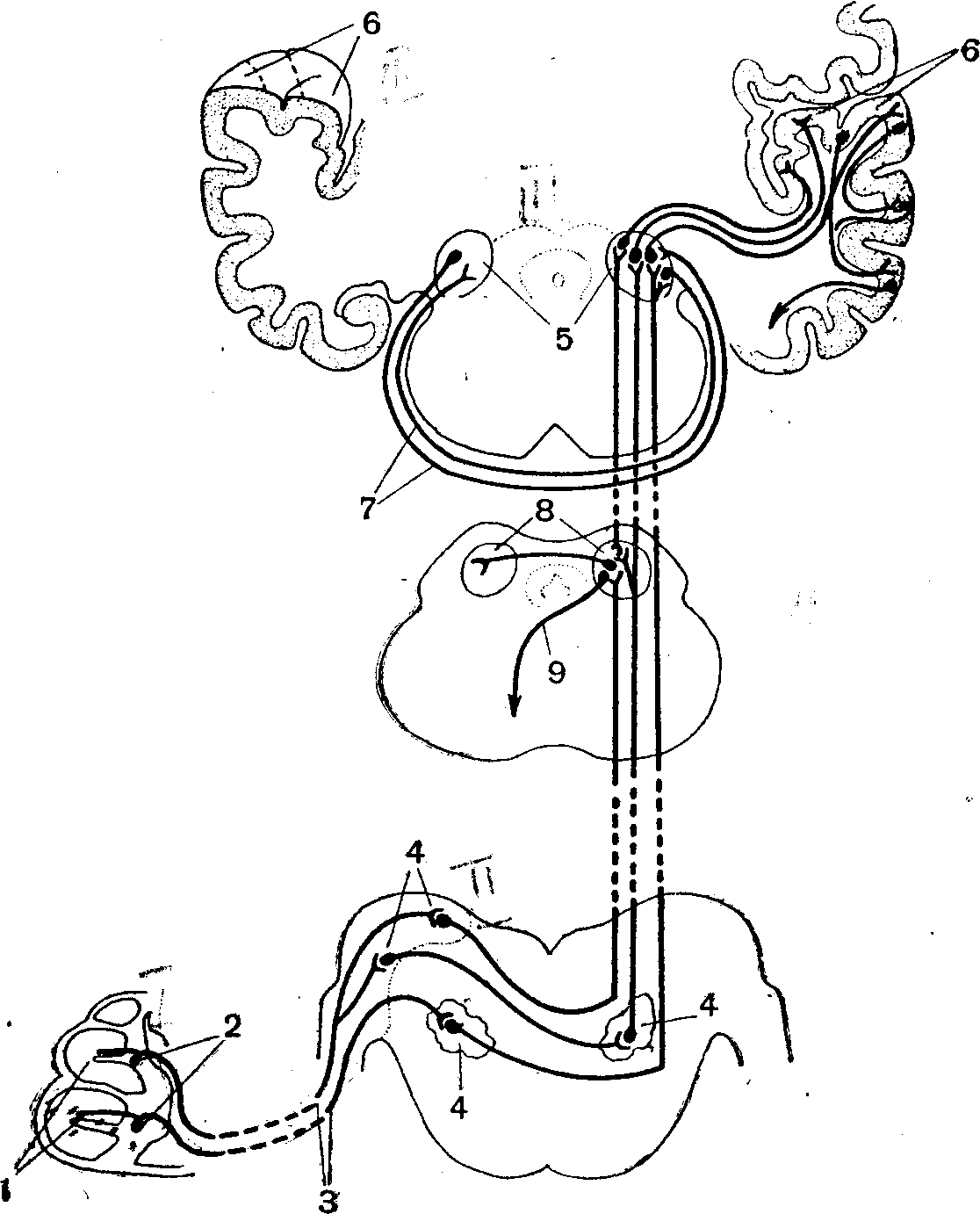
Рис. 5. Схема проводящих путей слухового анализатора:
1 — рецепторы кортиева органа; 2 — тела биполярных нейронов; 3 — улитковый нерв; 4 — ядра продолговатого мозга, где ' расположены тела второго нейрона проводящих путей; 5 — внутреннее коленчатое тело, где начинается третий нейрон основных проводящих путей; 6 •— верхняя поверхность височной доли коры больших полушарий (нижняя стенка поперечной щели), где оканчивается третий нейрон; 7 — нервные волокна, связывающие оба внутренних коленчатых тела; 8 — задние бугры четверохолмия; 9 — начало эфферентных путей, идущих от четверохолмия.
главным образом противоположной стороны. Здесь начинается третий нейрон, по которому импульсы достигают слуховой области коры больших полушарий (рис. 5).
Помимо основного, проводящего пути, связывающего периферический отдел слухового анализатора с его центральным, корковым отделом, существуют и другие пути, через которые могут осуществляться рефлекторные реакции на раздражение органа слуха у животного и после удаления больших полушарий. Особое значение имеют ориентировочные реакции на звук. Они осуществляются при участии четверохолмия, к задним и отчасти передним буграм которого идут коллатерали волокон, направляющихся к внутреннему коленчатому телу.
Корковый отдел слухового анализатора.У человека ядро коркового отдела слухового анализатора расположено ^в височной, области коры больших,, полушарий. В той части поверхности височной' области, которая представляет собой нижнюю стенку поперечной, или сильвиевой, щели, расположено поле 41. К нему, а возможно и к соседнему полк» 42, направляется основная масса волокон от внутреннего коленчатого тела. Наблюдения показали, что при двустороннем разрушении указанных полей наступает полная глухота. Однако в тех случаях, когда поражение ограничивается одним полушарием, может наступить небольшое и нередко лишь временное понижение слуха. Это объясняется тем, что проводящие пути слухового анализатора неполностью перекрещиваются. К тому же оба внутренних коленчатых тела связаны между собой промежуточными нейронами, через которые импульсы могут переходить с правой стороны на левую и обратно. В результате корковые клетки каждого полушария получают импульсы с обоих кортиевых органов.
От коркового отдела слухового анализатора идут эфферентные пути к нижележащим отделам мозга, и прежде всего к внутреннему коленчатому телу и к задним буграм четверохолмия. Через них осуществляются корковые двигательные рефлексы на звуковые раздражители. Путем раздражения слуховой области коры можно вызвать у животного ориентировочную реакцию настораживания (движения ушной раковины, поворот головы и т. п.). Анализ и синтез звуковых раздражении. Анализ звуковых раздражений начинается в периферическом отделе слухового анализатора, что обеспечивается особенностями строения улитки, и прежде всего основной пластинки, каждый участок которой колеблется в ответ на звуки только определенной высоты.
Высший анализ и синтез звуковых раздражении, основанный на образовании положительных и отрицательных условных связей, происходит в корковом отделе анализатора. Каждый звук, воспринимаемый кортиевым органом, приводит в состояние возбуждения определенные клеточные группы поля 41 и соседних с ним полей. Отсюда возбуждение распространяется в другие пункты коры больших полушарий, особенно в поля 22 и 37. Между различными клеточными группами, которые повторно приходил.i в состояние возбуждения пэд влиянием опргделеннэго звукового раздражения или комплекса последовательных звуковых раздражении, устанавливаются все более прочные условные связи. Они устанавливаются также между очагами возбуждения в слуховом анализаторе и теми очагами, которые одновременно возникают под влиянием раздражителей, действуюдих на другие анализаторы. Так образуются все новые и новыэ условные связи, обогащзюд ie анализ и синтез звуковых раздражении.
В основе анализа и синтеза звуковых речевых раздражении лежит установление условных связей между очагами возбуждения. которые возникают под влиянием непосредственных раздражителей, действуюдих на различные анализаторы, и теми очагами, которые вызываются звуковыми речевыми сигналами, обэзначаюдими эти раздражители. Так называемый слуховой центр речи, т. е. тот участок слухового анализатора, функция которого связана с речевым анализом и синтезом звуковых раздражении, иными словами, с пониманием слышимой речи, расположен в основном в левом полушарии и занимает задний конец поля и прилегающий участок поля.
Факторы, определяющие чувствительность слухового анализатора.Ухо человека особенно чувствительно к частоте звуковых и - колебаний от 1030 до 40ЭЭ в секунду. Чувствительность к более высоким и более низким звукам значительно падает, особенно с приближением к нижнему и верхнему пределам воспринимаемых частот. Так, для звуков, частота колебаний которых приближается к 20 или к 20 000 в секунду, порог повышается в 10 ООЭ раз, если определять силу звука по производимому им давлению. С возрастом чувствительность слухового анализатора, как правило, значительно понижается, но главным образом к звукам большой частоты, к низким же (до 1000 колебаний в секунду) остается почти неизменной вплоть до старческого возраста.
В условиях полной тишины чувствительность слуха повышается. Если же начинает звучать тон определенной высоты и неизменной интенсивности, то вследствие адаптации к нему ощущение громкости снижается сначала быстро, а потом все более медленно. Одновременно, хотя и в меньшей степени, понижается чувствительность к звукам, более или менее близким по частоте колебаний к звучащему тону. Однако обычно адаптация не распространяется на весь диапазон воспринимаемых звуков. По прекращении звучания вследствие адаптации к тишине уже через 10—15 секунд восстанавливается прежний уровень чувствительности.
Частично адаптация зависит от периферического отдела анализатора, а именно от изменения как усиливающей функции звукопроводящего аппарата, так и возбудимости волосковых клеток кортиева органа. Центральный отдел анализатора также принимает участие в явлениях адаптации, о чем свидетельствует хотя бы тот факт, что при действии звука только на одно ухо сдвиги чувствительности наблюдаются в обоих ушах. На чувствительность слухового анализатора, и в частности на процесс адаптации, оказывают влияние изменения корковой возбудимости, которые возникают в результате как иррадиации, так и взаимной индукции возбуждения и торможения при раздражении рецепторов других анализаторов. Изменяется чувствительность и при одновременном действии двух тонов разной высоты. В последнем случае слабый звук заглушается более сильным главным образом потому, что очаг возбуждения, возникающий в коре под влиянием сильного звука, понижает вследствие отрицательной индукции возбудимость других участков коркового отдела того же анализатора.
Длительное воздействие сильных звуков может вызвать запредельное торможение корковых клеток. В результате чувствительность слухового анализатора резко понижается. Такое состояние сохраняется некоторое время после того, как прекратилось раздра
Похожие работы

... сущности психологических явлений, выступающих в форме внутренних переживаний (ощущений, мыслей, чувств) и которые недоступны прямому наблюдению, происходит благодаря работе сенсорных систем. Сенсорные системы можно классифицировать на несколько групп. По характеру раздражителей: 1 – механические (тактильная, болевая, проприоцептивная, вестибулярная сенсорные системы, барорецептивный отдел ...
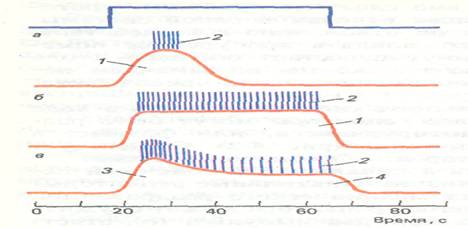
... четко выраженных компонента: начальный — фазный, последующий — тонический. Таким образом, организм, имея в своем составе все три типа рецепторов, получает информацию о начальных и конечных моментах воздействия раздражителя, а также о его силе и длительности. На основе деятельности тонических рецепторов может постоянно анализироваться и прослеживаться уровень сенсорного воздействия. Фазный тип ...

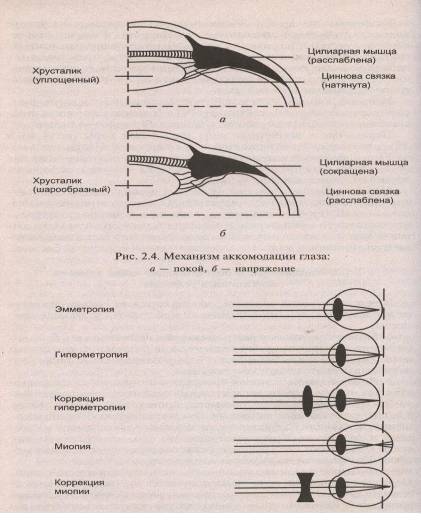
... устройствах, чтобы обеспечить адекватное понимание речи, речевая информация должна передаваться в речевом диапазоне частот. Структурно-функциональная характеристика Рецепторный (периферический) отдел слухового анализатора, превращающий энергию звуковых волн в энергию нервного возбуждения, представлен рецепторными волосковыми клетками кортиева органа {орган Корти), находящимися в улитке. ...
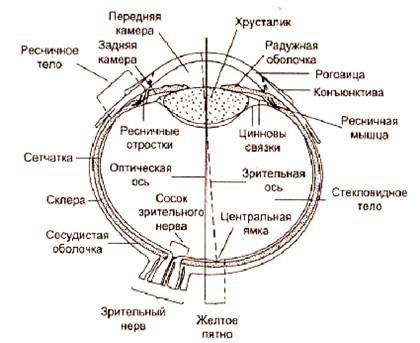
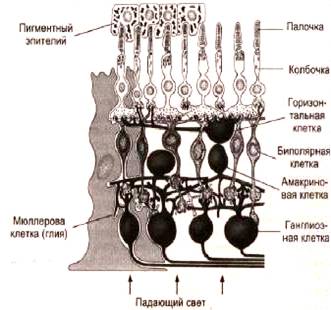
... основными, но нарушение любой из них отражается на качестве и количестве зрительной информации вплоть до полного прекращения ее поступления в ЦНС. ОПТИЧЕСКАЯ СИСТЕМА ГЛАЗА, СТРОЕНИЕ И ФИЗИОЛОГИЯ СЕТЧАТКИ К оптической системе глаза относятся: роговица, водянистая влага, радужка, зрачок, хрусталик и стекловидное тело Глазное яблоко, имеет шаровидную форму и помещается в костной ...
0 комментариев