Общая характеристика
Одна из трудно разрешимых проблем, с которой встречались исследователи при описании системы коммуникации, основной на использовании гормонов, представлено на схеме (рис. 1). Во внеклеточной жидкости гормоны присутствуют в очень низкой концентрации – обычно в пределах 10(-15) - 10(-19) моль/л. Это на много ниже содержание других, структурно сходных соединений (стеролов, аминокислот, пептидов, белков) и иных веществ, каторые находятся в крови в концентрации 10(-5) – 10(-3) моль/л. Следовательно, клетки-мишени должны отличать данный гормон не только от других гормонов, присутствующих в малых количествах, но и от прочих соединений, присутствующих в 10(6)– 10(9) – кратном количестве. Столь высокую степень избирательности обеспечивают особые принадлежащие клетке молекулы узнавания, называемые рецепторы. Биологический эффект гормонов начинается с их связывания со специфическими рецепторами, а завершается, как правило диссоциацией гормона и рецептора (в соответствии с тем принципов, что надежная система контроля должна обладать средством прерывания действия агента).

Рис 1. Специфичность и избирательность рецепторов гормонов. Во внеклеточной жидкости содержится множество разнообразных соединений, но рецепторы узнают лишь очень немногие из них. Кроме того, рецепторы должны выбрать определенные молекулы из множества других, присутствующих в более высокой концентрации. На рисунке показано, что каждая клетка может нести либо один тип рецепторов, либо несколько
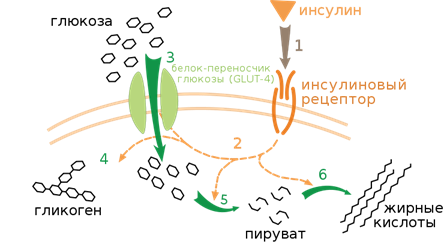
Механизм действия инсулинаРецепторы инсулина. Действие инсулина начинается с его связывания со специфическим гликопротеиновым рецептором на поверхности клетки-мишени. Различные эффекты этого гормона (рис 2.) могут проявляться либо через несколько секунд или минут (транспорт, фосфорилирование белков, активации и ингибирование ферментов, синтез РНК), либо через несколько часов (синтез белка и ДНК и клеточный рост).

Рис 2. Связь между рецептором инсулина и его действием. (Courtesy of C. R. Kahn.)
Инсулиновый рецептор подробно исследован с помощью биохимических методов и технологии рекомбинантных ДНК. Он представляет собой гетеродимер, состоящий из двух субъединиц (a и b) в конфигурации α2-β2, связанных между собой дисульфидными мостиками. Обе субъединицы содержат много гликозидных остатков. Удаление сиаловой кислоты и галактозы снижает как способность связывать инсулин, так и активность этого гормона. Каждая из гликопротеиновых субъединиц обладает особой структурой и определенной функцией. a-Субъединица (мол. масса 135000) целиком расположена вне клетки, связывание инсулина, вероятно, осуществляется с помощью богатого цистином домена. b-Субъединица (мол. масса 95000) – трансмембранный белок, выполняющий вторую важную функцию рецептора, т. е. преобразование сигнала. Цитоплазматическая часть b-субъединицы обладает тирозинкиназной активностью и содержит участок аутофорилирования. Считается, что и то и другое важно для преобразования сигнала и действия инсулина. Поразительное сходство между тремя рецепторами, выполняющими различные функции, проиллюстрировано на схеме (рис. 3). Действительно, последовательность некоторых участков b-субъединиц гомологичны таковым в рецепторе
 |
ФРЭ.
Рис 3. Схема строения рецепторов липопротеинов низкой плотности (ЛПНП), фактора роста эпидермиса (ФРЭ) и инсулина. В каждом из этих рецепторов аминоконцы находятся в той части молекулы, которая выступает из клетки. Рамками обозначены участки, богатые цистеином, которые, как считают, участвуют в связывании лиганда. В каждом рецепторе (~25 аминокислот) имеется короткий домен, пересекающий плазматическу мембрану (серая полоса), и внутриклеточный домен варьирующей длинны. Рецепторы ФРЭ и инсулина обладают тирозинкиназной активностью, локализованном в цитоплазматическом домене; кроме того, в этом домене находятся участки, в которых происходит аутофосфорилирование. Инсулиновый рецептор представляет собой гетеротетрамер, отдельные цепи (вертикальные полосы) которые связаны между собой дисульфидными мостиками.
Рецептор инсулина постоянно синтезируется и распадается; его период полужизни составляет 7 – 12 ч. Рецептор синтезируется в виде одноцепочечного пептида в шероховатом эндоплазматическом ретикулуме и быстро гликозируется в аппарате Гольджи. Предшественник человеческого рецептора инсулина состоит из 1382 аминокислот, его мол. масса составляет 190000, при расщеплении он образует зрелые a- и b-субъединицы. У человека ген инсулинового рецептора локализован в хромосоме 19.
Рецепторы инсулина обнаружены на поверхности большинства клеток млекопитающих. Их концентрация достигает 20000 на клетку, причем часто они выявляются и на таких клетках, которые не относятся к типичным мишеням инсулин. Спектр метаболических эффектов инсулина хорошо известен. Однако инсулин участвует и в таких процессах, как рост и репликация клеток, органогенез и дифференцировка у плода, а также в процессах заживления и регенерации тканей. Строение инсулинового рецептора, способность различных инсулинов связываются с рецепторами, и вызывать биологические реакции практически идентичны в клетках всех типов и всех видов. Так, свиной инсулин почти всегда в 10 – 20 раз эффективнее свиного проинсулина, который в свою очередь в 10 – 20 раз эффективнее инсулина морской свинки даже у самой морской свинки. Инсулиновый рецептор имеет, по-видимому, высоко консервативную структуру, еще более консервативную, чем структура самого инсулина.
При связывании инсулина с рецептором происходят следующие события: 1) изменяется конформация рецептора,
2) рецепторы связываются друг с другом, образуя микроагрегаты, пятна (patches) или нашлепки,
3) рецептор подвергается интернализации,
4) возникает какой то сигнал.
Значение конформационных изменений рецептора не известно, но интернализация, вероятно, служит средством регуляции количества и кругооборота рецепторов. В условиях высокого содержания инсулина в плазме, например при ожирении и акромегалии, число инсулиновых рецепторов снижается, и чувствительность тканей-мишений к инсулину уменьшается. Такая «снижающая» регуляция обусловлена потерей рецепторов в результате их интернализации, т. у. Процесса проникновения инсулин-рецепторных комплексов в клетку путем эндоцитоза с помощью покрытых клатрином пузырьков.
В центре внимания современных исследователей лежит тот факт, что инсулиновый рецептор сам является ферментом, чувствительным к инсулину, поскольку при связывании инсулина он подвергается аутофосфорилированию. Эта функция осуществляется b-субъединицей, которая, действует как протеинкиназа, переносит g-фосфат с АТР на остаток тирозина b-субъединице. Инсулин повышает Vmax этой ферментативной реакции, а в двухвалентные катионы, особенно Mn²_, снижают Км для АТР.
Фосфорилирование тирозина нетипично для клеток млекопитающих (на долю фосфотирозина приходится всего 0,03% фосфоаминокислот, содержащихся в нормальных клетках), и вполне возможно, что наличие у рецепторов ФРЭ, ТФР, ИФР-1 тирозинкиназной активности не случайно. Существует предложение, что тирозинкиназная активность – важный фактор в действии продуктов ряда вирусных анкогенов. Их связь с клеточными аналогами онкогенов, обладающими сходными свойствами при злокачественном и нормальном клеточном росте, расматривается выше. Изучение структуры этих компонентов выявило высокую степень гомологии между рецепторами и онкогенами, например между рецептором ФРЭ и erb-B, между рецептором ТФР и v-sis и между инсулиновым рецептором v-ros.
Участие тирозинкиназы в преобразовании инсулин-рецепторного сигнала не доказано, но оно могло бы заключатся в фосфорилировании специфического белка, инициирующего действие инсулина, в запуске каскада фосфорилирование-дефосфорилирование, в изменение некоторых свойств клеточной мембраны или образование какого-то связанного с мембраной продукта, например фосфолипида.
Фосфолирование-дефосфолирование белка.
Многие из метаболических эффектов инсулина, особенно те, которые возникают быстро, опосредованы его влиянием на реакции фосфолирование-дефосфолирование белка, что в свою очередь влияет на ферментативную активность данного белка. Перечень ферментов, активность которых регулируется таким путем, приведен в таблице (?). в некоторых случаях инсулин снижает внутриклеточное содержание сАМР (активируя сАМР-фосфодиэстеразу), что приводит к уменьшению активности сАМР-зависимой протеинкиназы. Такие эффекты характерны для гликогенсинтазы и фосфорилазы. В других случаях действия инсулина не зависит от сАМР и сводится к активации других протеинкиназ (например, в случае тирозинкиназы инсулинового рецептора), ингибированию третьих протеинкиназ или (что значительно чаще) к стимуляции фосфотаз фосфопротеинов. Дефосфолирование увеличивает активность ряда ключевых ферментов (таб.1). Такие ковалентные модификации обеспечивают почти мгновенные изменения активностей ферментов.
Таблица 1.Ферменты, степень фосфорилирования и активность каторых регулируется инсулином.
| Фермент | Изменения активности | Возможный механизм |
| Метаболизм сАМР Фосфодиэстераза низкая КМ | повышается | Фосфолирование |
| протеинкиназа (сАМР-зависимая) | понижается | » |
| Метаболизм гликргена Гликогенсинтаза | повышается | Дефосфолирование |
| Киназа фосфорилазы | понижается | » |
| Гликолиз и глюконеогенез Пируватдегидрогеназа | повышается | » |
| Пируваткиназа | » | » |
| 6-Фосфофрукто-2-киназа | » | » |
| Фруктоза-2,6- бисфосфатаза | понижается | » |
| Обмен липидов Ацетил-СоА – карбоксилаза | повышается | Фосфолирование |
| ГМГ-СоА-редуктаза (гидроксиметилглутарил -СоА-редуктаза) | » | Дефосфолирование |
| Триацилглицероллипаза | понижается | » |
| Другие процессы Тирозинкиназа(рецептор инсулина) | ? | Фосфолирование |
При недостаточности инсулина или устойчивости к его действию развивается сахарный диабет. Примерно у 90% больных диабетом наблюдается инсулин-независимый сахарный диабет II типа (ИНЗСД). Для таких больных характерны ожирение, повышенное содержание в плазме инсулина и снижение количества инсулиновых рецепторов.
У некоторых людей образуются антитела к рецепторам инсулина. Эти тела предотвращают связывание инсулина с рецептором, и в результате у таких лиц развивается синдром тяжелой инсулинорезистентности. О важной роли инсулина для органогенеза свидетельствуют редкие случаи карликовости. Этот синдром характеризуется низким весом при рождении, малой мышечной массой, малым количеством подкожного жира, очень мелкими чертами лица, инсулинорезистентностью со значительным повышением содержания биологически активного в плазме и ранней смертью. У некоторых таких больных либо совсем отсутствовали рецепторы инсулина, либо они были дефективными
Список литературы:
1. Гренер Д. К. Марри. Р. Биохимия человека.1998г.
2. Кохн К. Р. Молекулярный механизм действия инсулина. 1985г.
3. Andreone I. Insulin modulation of gene expression, In: Diabetes and Metabolism Reviews. 1985
4. Kono T. Action of insulin on glucose transport and cAMP phosphodiesterase in fat cells: Involvement of two distinct molecular mechanisms. 1983
![]()
![]()
![]()
![]()
Похожие работы
... , глюкагономы, соматостатиномы, гипертиреоз) – избыточная секреция соответствующих гормонов, обладающих чётко выраженным контринсулиновым действием. - К лекарствам и химическим веществам, индуцирующим сахарный диабет, относятся глюкокортикостероиды, тиреоидные гормоны, α- и β-адренергические агонисты, никотиновая кислота, тиазиды, дилантин, пентамидин. - Некоторые ...
... в крови антител к β-клеткам поджелудочной железы. Характер наследования в настоящее время не совсем ясен, сложность прогнозирования наследования связана с генетической гетерогенностью сахарного диабета, построение адекватной модели наследования требует дополнительных статистических и генетических исследований. Патогенез В патогенезе сахарного диабета выделяют два основных момента: 1) ...
... воздействию на эксплантный эндометрий при аденомиозе, дает высокий терапевтический эффект. Исчезновение узлов при небольших размерах, профилактика их формирования при внутреннем эндометриозе, уменьшение объема миомы матки на 1/3—1/4 первоначальной величины при значительных ее размерах путем перорального назначения неместрана в дозе 5—10 мг в неделю отмечено в 2/3 наблюдений в пределах 4—12 ...
... синдрома может быть врожденный или приобретенный дефект диафрагмы седла, а также повышение давления в супраселлярной цистерне. 3.1 Лабораторная диагностика Для оценки функционального состояния гипоталамо-гипофизарной системы используется определение в крови целого комплекса гормонов, обеспечивающего всю линию функциональной связи: гипоталамус → гипофиз → периферическая железа ...
0 комментариев