План:
Введение. Исследование белков. Классификация белков. Состав и строение пептидная связь элементарный состав молекулярная масса аминокислоты строениеа) первичная структура
б) вторичная структура
в) третичная структура
· пространственная структура
г) четвертичная структура
· денатурация
Химические и физические свойства. Химический синтез белков. Значение белков. Вывод.Список использованной литературы.
Введение
Белки - высокомолекулярные азотистые органические вещества, построенные из аминокислот и играющие фундаментальную роль в структуре и жизнедеятельности организмов. Белки – основная и необходимая составная часть всех организмов. Именно Белки осуществляют обмен веществ и энергетические превращения, неразрывно связанные с активными биологическими функциями. Сухое вещество большинства органов и тканей человека и животных , а также большая часть микроорганизмов состоят главным образом из белков (40-50%), причем растительному миру свойственно отклонение от этой средней величины в сторону понижения, а животному – повышения. Микроорганизмы обычно богаче белком (некоторые же вирусы являются почти чистыми белками). Таким образом, в среднем можно принять, что 10% биомассы на Земле представлено белком, то есть его количество измеряется величиной порядка 1012- 1013тонн. Белковые вещества лежат в основе важнейших процессов жизнедеятельности. Так , например , процессы обмена веществ ( пищеварение, дыхание, выделение, и другие) обеспечиваются деятельностью ферментов , являющихся по своей природе белками. К белкам относятся и сократительные структуры, лежащие в основе движения, например сократительный белок мышц ( актомиозин), опорные ткани организма (коллаген костей, хрящей, сухожилий) , покровы организма ( кожа, волосы, ногти и т.п.) , состоящие главным образом из коллагенов, эластинов, кератинов, а также токсины, антигены и антитела, многие гормоны и другие биологически важные вещества. Роль белков в живом организме подчеркивается уже самим их названием «протеины» ( в переводе с греческого protos – первый, первичный) , предложенным в 1840 голландским химиком Г. Мульдером, который обнаружил , что в тканях животных и растений содержатся вещества, напоминающие по своим свойствам яичный белок. Постепенно было установлено, что белки представляют собой обширный класс разнообразных веществ, построенных по одинаковому плану. Отмечая первостепенное значение белков для процессов жизнедеятельности, Энгельс определил, что жизнь есть способ существования белковых тел, заключающийся в постоянном самообновлении химических составных частей этих тел.
В природе существует примерно 1010-1012 различных белков, обеспечивающих жизнедеятельность организмов всех степеней сложности от вирусов до человека, они обеспечивают жизнь более 2 млн. видам организмов. Белками являются ферменты, антитела, многие гормоны и другие биологические активные вещества. Необходимость постоянного обновления белков лежит в основе обмена веществ. Именно поэтому белки и явились тем исключительным материалом , который послужил основой возникновения жизни на Земле. Ни одно вещество из всех веществ биологического происхождения не имеет столь большого значения и не обладает столь многогранными функциями в жизни организма как белки.
Ф. Энгельс писал: „Повсюду, где мы встречаем жизнь, мы находим, что она связана с каким-либо белковым телом и повсюду, где мы встречаем какое-либо белковое тело, которое не находится в процессе разложения, мы без исключения встречаем и явления жизни“.
Исследование белков
Свое название белки получили от яичного белка, который с незапамятных времен использовался человеком как составная часть пищи. Согласно описаниям Плиния Старшего, уже в Древнем Риме яичный белок применялся и как лечебное средство. Однако подлинная история белковых веществ начинается тогда, когда появляются первые сведения о свойствах белков как химических соединений (свертываемость при нагревании, разложение кислотами и крепкими щелочами и т. п.). Среди белков животного происхождения, вслед за яичным белком, были охарактеризованы белки крови. Образование сгустков крови при ее свертывании описано еще основателем учения о кровообращении У. Гарвеем; позднее на этот факт обратил внимание и Р. Бойль. Среди растительных белков пальма первенства принадлежит нерастворимой в воде клейковине из пшеничной муки, которую впервые получил Я. Беккари. В своих работах, он отметил сходство клейковины с веществами животной природы.
Впервые термин белковый (albumineise) применительно ко всем жидкостям животного организма использовал французский физиолог Ф. Кене в 1747 г., и именно в таком толковании термин вошел в 1751 г. в «Энциклопедию» Д. Дидро и Ж. Д'Аламбера.
С этого периода исследования, связанные с получением белков, приобретают систематический характер. В 1759 г. А. Кессель-Майер, а несколько позднее И. Руэль описали выделение клейковины из различных растений и охарактеризовали ее свойства. В 1762 г. А. Халлер исследовал процесс образования и свертывания казеина, а в 1777 г. А. Тувенель, работавший тогда в Петербурге, называет творог белковой частью молока . Важнейший этап в изучении белков связан с работами французского химика А. Фуркруа, который рассматривал белки как индивидуальные вещества и доказал единую природу белковых веществ, выделенных из растительных и животных источников. Для трех главных белковых компонентов крови он предложил названия альбумин, желатин и фибрин. В 1780 г. Ф. Вассерберг относит к телам белковой природы хрусталик глаза.
К началу XIX столетия появляются первые работы по химическому изучению белков. Уже в 1803 г. Дж. Дальтон дает первые формулы белков - альбумина и желатина - как веществ, содержащих азот. В 1810 г. Ж. Гей-Люссак проводит химические анализы белков - фибрина крови, казеина и отмечает сходство их элементного состава. Решающее значение для понимания химической природы белков имело выделение при их гидролизе аминокислот. Вероятно, первым это сделал А. Браконно в 1820 г., когда, действуя на белки серной кислотой, при кипячении он получил «клеевой сахар», или глицин, при гидролизе фибрина из мяса - лейцин и при разложении шерсти - также лейцин и смесь других продуктов гидролиза. Первой открытой аминокислотой был, видимо, аспарагин, выделенный Л. Вокленом из сока спаржи Asparagus (1806). В это же время Ж. Пруст получил лейцин при разложении сыра и творога. Затем из продуктов гидролиза белка были выделены многие другие аминокислоты.
Первая концепция строения белков принадлежит голландскому химику Г. Мульдеру (1836). Основываясь на теории радикалов, он сформулировал понятие о минимальной структурной единице, входящей в состав всех белков. Эту единицу, которой приписывался состав 2C8H12N2 + 50, Мульдер назвал протеином (Рг), а свою концепцию - теорией протеина.Позднее состав протеина был уточнен – C40H62N10O12; дополнительно к протеинным единицам некоторые белки содержали серу и фосфор. Формула белков, предложенная Мульдером в 1838 г., выглядела так:
белок сыворотки крови 10Pr S2P
белок куриных яиц 10Pr SP
фибрин 10Pr SP
казеин 10Pr S
клейковина растений 10Pr S2
кристаллин (из хрусталика глаза) 15Рг
Работы Г. Мульдера способствовали широкому распространению взглядов о единстве всех белков, их фундаментальном значении в мире живой природы.
В ходе проверки «теории протеина» были резко расширены химические исследования белков, и в этом приняли участие выдающиеся химики того времени Ю. Либих и Ж. Дюма. Ю. Либих, поддерживавший в принципе идею протеиновой единицы, уточнил формулу протеина C48H72N12O14, Ж. Дюма предложил свой вариант C48H74 N 12О15 -, однако Г. Мульдер отстаивал правильность составленной им формулы. Его поддерживал И. Берцелиус, изложивший теорию протеина в качестве единственной теории строения белка в знаменитом учебнике химии (1840), что означало полное признание и торжество концепции Г. Мульдера.
Однако вскоре наступают трудные времена для теории протеина. В 1846 г. Н. Э. Лясковский, работавший в лаборатории Ю. Либиха, доказал неточность многих приведенных Г. Мульдером анализов. Свои сомнения в правильности теории публично высказал Ю. Либих, он планировал начать широкие исследования структуры белков и даже изучил продукты распада белковых веществ. Понимая весомость аргументов оппонентов, Г. Мульдер пытался корректировать формулу протеина (C36H50N8O10), но в конце концов уступил под натиском новых фактов и открытий. Теория протеина стала достоянием истории, однако ее значение непреходяще, ибо она стимулировала химические исследования белков, сделала белки одним из главных объектов бурно развивающейся химии природных веществ.
Открытие аминокислот в составе белков
| Аминокислота | Год | Источник | Кто впервые выделил |
| Глицин | 1820 | Желатина | А. Браконно |
| Лейцин | 1820 | Мышечные волокна | А, Браконно |
| 1839 | Фибрин шерсти | Г. Мульдер | |
| Тирозин | 1848 | Казеин | Ф. Бопп |
| Серии | 1865 | Шелк | Э. Крамер |
| Глутаминовая кислота | 1866 | Растительные белки | Г. Риттхаузен |
| Аспарагиновая кислота | 1868 | Конглутин, легумин (ростки спаржи) | Г. Риттхаузен |
| Фенилаланин | 1881 | Ростки люпина | Э. Шульце, И, Барбьери |
| Аланин | 1888 | Фиброин шелка | Т. Вейль |
| Лизин | 1859 | Казеин | Э. Дрексель |
| Аргинин | 1895 | Вещество рога | С. Гедин |
| Гистидин | 1896 | Стурин,гистоны | А. Кессель |
| Цистин | 1899 | Вещество рога | К. Мёрнер |
| Валин | 1901 | Казеин | Э. Фишер |
| Пролин | 1901 | Казеин | Э. Фишер |
| Гидроксипролин | 1902 | Желатина | Э. Фишер |
| Триптофан | 1902 | Казеин | Ф.Гопкинс, Д, Кол |
| Изолейцин | 1904 | Фибрин | Ф.Эрлих |
| Метионин | 1922 | Казеин | Д. Мёллер |
| Треонин | 1925 | Белки овса | С. Шрайвер и др. |
| Гидроксилизин | 1925 | Белки рыб | С. Шрайвер и др. |
Для формирования современных представлений о структуре белка существенное значение имели работы по расщеплению белковых веществ протеолитическими ферментами. Одним из первых их использует Г. Мейснер. В 1850 г. К. Леман предлагает называть пептонами продукты разложения белков пепсином. Изучая этот процесс, Ф. Хоппе-Зайлер и Ш. Вюрц в 70-х годах прошлого столетия пришли к важному выводу, что пептоны образуются в результате гидролиза белков ферментом. Они были весьма близки к правильному толкованию таких экспериментов с позиций структурной химии, но, к сожалению, последнего шага на пути к теории строения белка сделать не сумели. Очень близок к истине был и А. Я. Данилевский, который в своей работе "Исследование состава, физического и химического строения продуктов распадения белковых веществ и генетических отношений между различными их видами" справедливо утверждал, что белки построены из аминокислот и имеют полимерную природу.
Дальнейшие структурные исследования белка, а также основополагающие работы Т. Курциуса по синтезу пептидов привели в конце концов к формулированию пептидной гипотезы, согласно которой белки построены из аминокислот, соединенных пептидными связями -СО-NH-. В 1902 Э. Фишер создал метод анализа и разделения аминокислот, основанный на переводе их в сложные эфиры, которые можно было подвергать фракционной перегонке, не опасаясь разложения. С помощью этого метода провел качественное и количественное определение продуктов расщепления белков и открыл аминокислоты валин, пролин и гидроксипролин. Позднее из аминокислот он получил продукты их конденсации, названные полипептидами. Последовательно синтезировал ди-, три- и т.д. пептиды, всего около 125. Один из них, состоящий из 18 аминокислот, долгое время оставался наиболее сложным из всех синтезированных органических соединений с известной структурой. Фишер установил механизм соединения аминокислот в линейные цепочки через образование пептидной связи (и ввел этот термин), разработал методы синтеза D- и L-аминокислот. Пептидная теория получила полное подтверждение в дальнейших исследованиях. Изучение строения белков было поставлено на прочную научную основу.
В 1934 г. Лайнус Полинг совместно с А.E. Мирски сформулировал теорию строения и функции белка. В 1936 г. он положил начало изучению атомной и молекулярной структуры белков и аминокислот (мономеров, из которых состоят белки) с применением рентгеновской кристаллографии.В 1942 г. Полингу и его коллегам, получив первые искусственные антитела, удалось изменить химическую структуру некоторых содержащихся в крови белков, известных как глобулины.В 1951 г. П. и Р.Б. Кори опубликовали первое законченное описание молекулярной структуры белков. Это был результат исследований, длившихся долгих 14 лет. Применяя методы рентгеновской кристаллографии для анализа белков в волосах, шерсти, мускулах, ногтях и других биологических тканях, они обнаружили, что цепи аминокислот в белке закручены одна вокруг другой таким образом, что образуют спираль. Это описание трехмерной структуры белков ознаменовало крупный прогресс в биохимии.
Классификация белков.
Из-за относительно больших размеров белковых молекул , сложности их строения и отсутствия достаточно точных данных о структуре большинства белков еще нет рациональной химической классификации белков. Существующая классификация в значительной мере условна и построена главным образом на основании физико-химических свойств белков, источников их получения , биологической активности и других, нередко случайных, признаков. Так, по физико-химическим свойствам белки делят на фибриллярные и глобулярные , на гидрофильные(растворимые) и гидрофобные (нерастворимые) и т.п. По источнику получения белки подразделяют на животные, растительные и бактериальные; на белки мышечные, нервной ткани, кровяной сыворотки и т.п.; по биологической активности – на белки-ферменты, белки-гормоны, структурные белки, сократительные белки, антитела и т.д. Следует, однако, иметь в виду, что из-за несовершенства самой классификации, а также вследствие исключительного многообразия белков многие из отдельных белков не могут быть отнесены ни к одной из описываемых здесь групп.
Все белки принято делить на простые белки ,или протеины, и сложные белки , или протеиды (комплексы белков с небелковыми соединениями).Простые белки являются полимерами только аминокислот; сложные, помимо остатков аминокислот, содержат также небелковые, так называемые простетические группы.
Протеины представляют собой простые белки, состоящие только из остатков аминокислот. Они широко распространены в животном и растительном мире.
Гистоны
Имеют сравнительно низкую молекулярную массу (12-13 тыс.), с преобладанием щелочных свойств. Локализованы в основном в ядрах клеток. Растворимы в слабых кислотах, осаждаются аммиаком и спиртом. Имеют только третичную структуру. В естественных условиях прочно связаны с ДНК и входят в состав нуклеопротеидов. Основная функция — регуляция передачи генетической информации с ДНК и РНК (возможна блокировка передачи).
Протамины
Самая низкая молекулярная масса (до 12 тыс.). Проявляет выраженные основные свойства. Хорошо растворимы в воде и слабых кислотах. Содержатся в половых клетках и составляют основную массу белка хроматина. Как и гистоны образуют комплекс с ДНК, функция - придают ДНК химическую устойчивость.
Глютелины
Растительные белки, содержащиеся в клейковине семян злаковых и некоторых других, в зеленых частях растений. Нерастворимые в воде, растворах солей и этанола, но хорошо растворимы в слабых растворах щелочей. Содержат все незаменимые аминокислоты, являются полноценными продуктами питания.
Проламины
Растительные белки. Содержатся в клейковине злаковых растений. Растворимы только в 70%-м спирте (это объясняется высоким содержанием пролина и неполярных аминокислот).
ПротеиноидыБелки опорных тканей (кость, хрящ, связки, сухожилия, ногти, волосы). Нерастворимые или трудно растворимые в воде, солевых и водно-спиртовых смесях белки с высоким содержанием серы. К протеиноидам относятся кератин, коллаген, фиброин.
АльбуминыНевысокой молекулярной массой (15-17 тыс.). Характерны кислые свойства. Растворимы в воде, и слабых солевых растворах. Осаждаются нейтральными солями при 100%-м насыщении. Участвуют в поддержании осмотического давления крови, транспортируют с кровью различные вещества. Содержатся в сыворотке крови, молоке, яичном белке.
ГлобулиныМолекулярная масса до 100 тыс.. В воде нерастворимы, но растворимы в слабых солевых растворах и осаждаются в менее концентрированных растворах (уже при 50%-м насыщении). Содержатся в семенах растений, особенно в бобовых и масленичных; в плазме крови и в некоторых других биологических жидкостях. Выполняющие функцию иммунной защиты, обеспечивают устойчивость организма к вирусным инфекционным заболеваниям.
Сложные белки делят на ряд классов в зависимости от характера простетической группы.
Фосфопротеины
Имеют в качестве небелкового компонента фосфорную кислоту. Представителями данных белков являются казеиноген молока, вителлин (белок желтков яиц). Такая локализация фосфопротеидов свидетельствует о важном их значении для развивающегося организма. У взрослых форм эти белки присутствуют в костной и нервной тканях.
ЛипопротеиныСложные белки, простетическая группа которых образована липидами. По строению это небольшого размера (150-200 нм) сферические частицы, наружная оболочка которых образована белками (что позволяет им передвигаться по крови), а внутренняя часть — липидами и их производными. Основная функция липопротеинов — транспорт по крови липидов. В зависимости от количества белка и липидов, липопротеиды подразделяются на хиломикроны, липопротеиды низкой плотности (ЛПНП) и высокой плотности (ЛПВП), которые иногда обозначаются как a- и b-липопротеиды.
Металлопротеины
Содержат катионы одного или нескольких металлов. Наиболее часто это — железо, медь, цинк, молибден, реже марганец, никель. Белковый компонент связан с металлом координационной связью.
Гликопротеины
Простетическая группа представлена углеводами и их производными. Исходя из химического строения углеводного компонента, выделяют 2 группы:
Истинные — в качестве углеводного компонента наиболее часто встречаются моносахариды. Протеогликаны — построены из очень большого числа повторяющихся единиц, имеющих дисахаридный характер (гиалуроновая кислота, гипарин, хондроитин, каротинсульфаты).
Функции: структурно-механическую (имеются в коже, хряще, сухожилиях); каталитическую (ферменты); защитную; участие в регуляции клеточного деления.
ХромопротеиныВыполняют ряд функций: участие в процессе фотосинтеза и окислительно-восстановительных реакциях, транспорт С и СО2. Являются сложными белками, простетическая группа которых представлена окрашенными соединениями.
Нуклеопротеины
Роль протеистической группы выполняет ДНК или РНК. Белковая часть представлена в основном гистонами и протаминами. Такие комплексы ДНК с протаминами обнаружены в сперматозоидах, а с гистонами — в соматических клетках, где молекула ДНК “намотана” вокруг молекул белка-гистона. Нуклепротеинами по своей природе являются вне клетки вирусы — это комплексы вирусной нуклеиновой кислоты и белковой оболочки — капсида.
Состав и строение
Пептидная связь
Белки представляют собой нерегулярные полимеры, построенные из остатков a-аминокислот, общую формулу которых в водном растворе при значениях pH близких к нейтральным можно записать как NH3+CHRCOO– . Остатки аминокислот в белках соединены амидной связью между a-амино- и a-карбоксильными группами. Связь между двумя a-аминокислотными остатками обычно называется пептидной связью, а полимеры, построенные из остатков a-аминокислот, соединенных пептидными связями, называют полипептидами. Белок как биологически значимая структура может представлять собой как один полипептид, так и несколько полипептидов, образующих в результате нековалентных взаимодействий единый комплекс.
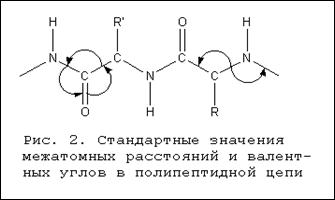 Все входящие в пептидную связь атомы располагаются в одной плоскости (планарная конфигурация).
Все входящие в пептидную связь атомы располагаются в одной плоскости (планарная конфигурация).
Расстояние между атомами С и N (в -СО-NH-связи) равно 0,1325 нм, то есть меньше нормального расстояния между a-углеродным атомом и атомом N той же цепи, выражаемого величиной 0,146 нм. Вместе с тем оно превышает расстояние между атомами С и N, соединенными двойной связью (0,127 нм). Таким образом, связь С и N в -СО-NH -группировке может рассматриваться как промежуточная между простой и двойной вследствие сопряжения π-электронов карбонильной группы со свободными электронами атома азота. Это определенным образом сказывается на свойствах полипептидов и белков: по месту пептидных связей легко осуществляется таутомерная перегруппировка, приводящая к образованию енольной формы пептидной связи, отличающейся повышенной реакционной способностью.
Элементный состав белков
Белки содержат в среднем около 1 6% азота, 50-55% углерода , 21-23% кислорода , 15-17% азота , 6-7% водорода , 0,3-2,5% серы . В составе отдельных белков обнаружены также фосфор, йод, железо, медь и некоторые другие макро- и микроэлементы, в различных, часто очень малых количествах.
Содержание основных химических элементов в белках может различаться, за исключением азота, концентрация которого характеризуется наибольшим постоянством.
Для изучения аминокислотного состава белков используется главным образом метод гидролиза, то есть нагревание белка с 6-10 моль/ литр соляной кислотой при температуре 100-110 0С. получают смесь a-аминокислот, из которых можно выделить индивидуальные аминокислоты. Для количественного анализа этой смеси в настоящее время применяют ионообменную и бумажную хроматографию. Сконструированы специальные автоматические анализаторы аминокислот.
Разработаны также ферментативные методы ступенчатого расщепления белка. Некоторые ферменты расщепляют макромолекулу белка специфически – только в местах нахождения определенной аминокислоты. Так получают продукты ступенчатого расщепления - пептоны и пептиды, последующим анализом которых устанавливают их аминокислотный остаток.
В результате гидролиза различных белков выделено не более 30 a-аминокислот. Двадцать из них встречаются чаще других.
При образовании молекулы белка или полипептида a-аминокислоты могут соединяться в различной последовательности . Возможно огромное число различных комбинаций, например из 20 a-аминокислот можно образовать больше 1018 комбинаций. Существование различного типа полипептидов практически неограничено.
Последовательность соединения аминокислот в том или ином белке устанавливают путем ступенчатого расщепления или рентгеноструктурным анализом.
Для идентификации белков и полипептидов используют специфические реакции на белки. Например :
а) ксантопротеиновая реакция ( появление желтого окрашивания при взаимодействии с концентрированной азотной кислотой, которое в присутствии аммиака становиться оранжевым ; реакция связана с нитрованием остатков фенилаланина и тирозина);
б) биуретовая реакция на пептидные связи – действие разбавленного сульфата меди (II) на слабощелочной раствор белка сопровождающийся появлением фиолетово-синей окраски раствора ,что обусловлено комплексообразованием между медью и полипептидами.
в) реакция Миллона (образование желто-коричневого окрашивания при взаимодействии с Hg(NO3)2 + HNO3 + HNO2;
Молекулярная масса
Белки являются высокомолекулярными соединениями. Это полимеры, состоящие из сотен и тысяч аминокислотных остатков — мономеров. Соответственно и молекулярная масса белков находится в пределах 10000-1000000. Так, в составе рибонуклеазы (фермента, расщепляющего РНК) содержится 124 аминокислотных остатка и ее молекулярная масса составляет примерно 14000. Миоглобин (белок мышц), состоящий из 153 аминокислотных остатков, имеет молекулярную массу 17000, а гемоглобин – 64500 (574 аминокислотных остатка). Молекулярные массы других белков более высокие: g-глобулин (образует антитела) состоит из 1250 аминокислот и имеет молекулярную массу около 150000, а молекулярная масса белка вируса гриппа – 320 000 000.
Аминокислоты
В настоящее время в различных объектах живой природы обнаружено до 200 различных аминокислот. В организме человека их, например, около 60. Однако в состав белков входят только 20 аминокислот, называемых иногда природными.
Аминокислоты — органические кислоты, у которых атом водорода a-углеродного атома замещен на аминогруппу –NH2. Следовательно, по химической природе это a-аминокислоты с общей формулой:
COOH
![]()
H–C*–NH2
![]()
R
Из формулы видно, что в состав всех аминокислот входят следующие общие группировки: –C–, –NH2, –COOH. Боковые же цепи (радикалы –R) аминокислот различаются. Природа радикалов разнообразна: от атома водорода до циклических соединений. Именно радикалы определяют структурные и функциональные особенности аминокислот.
Все аминокислоты, кроме простейшей аминоуксусной кислоты — глицина (NH3+CH2COO-)имеют хиральный атом - C*- и могут существовать в виде двух энантиомеров (оптических изомеров): L-изомер и D-изомер.
В состав всех изученных в настоящее время белков входят только аминокислоты L-ряда, у которых, если рассматривать хиральный атом со стороны атома H, группы NH3+, COO- и радикал -R расположены по часовой стрелке. Необходимость при построении биологически значимой полимерной молекулы строить ее из строго определенного энантиомера очевидна — из рацемической смеси двух энантиомеров получилась бы невообразимо сложная смесь диастереоизомеров. Вопрос, почему жизнь на Земле основана на белках, построенных именно из L-, а не D-a-аминокислот, до сих пор остается интригующей загадкой. Следует отметить, что D-аминокислоты достаточно широко распространены в живой природе и, более того, входят в состав биологически значимых олигопептидов.
Структура
При изучении состава белков было установлено, что все они построены по единому принципу и имеют четыре уровня организации: первичную, вторичную, третичную, а отдельные из них и четвертичную структуры.
Первичная структура
Представляет собой линейную цепь аминокислот (полипептид), расположенных в определенной последовательности с четким генетически обусловленным порядком чередования и соединенных между собой пептидными связями.
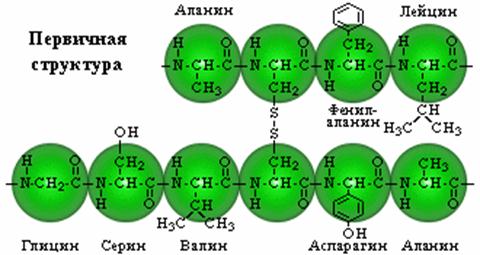

Пептидная связь образуется за счет a-карбоксильной группы одной аминокислоты и a-аминной группы другой
К настоящему времени установлены последовательности аминокислот для нескольких тысяч различных белков. Запись структуры белков в виде развернутых структурных формул громоздка и не наглядна. Поэтому используется сокращенная форма записи — трехбуквенная или однобуквенная.
При записи аминокислотной последовательности в полипептидных или олигопептидных цепях с помощью сокращенной символики предполагается, если это особо не оговорено, что a-аминогруппа находится слева, а a-карбоксильная группа — справа. Соответствующие участки полипептидной цепи называют N-концом (аминным концом) и С-концом (карбоксильным концом), а аминокислотные остатки — соответственно N-концевым и С-концевым остатками.
Вторичная структура
Вторичной структурой называют конформацию, которую образует полипептидная цепь. Для высокомолекулярных белков характерна структура спирали.
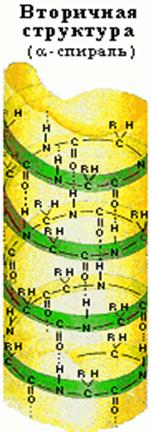 Впервые такая структура на основе рентгеноструктурного анализа была обнаружена при изучении главного белка волос и шерсти -a- кератина (Л. Полинг). Ее назвали a-структурой или a-спиралью. Обычно в природных продуктах встречаются белки со строением правой спирали, хотя известна и структура левой спирали.
Впервые такая структура на основе рентгеноструктурного анализа была обнаружена при изучении главного белка волос и шерсти -a- кератина (Л. Полинг). Ее назвали a-структурой или a-спиралью. Обычно в природных продуктах встречаются белки со строением правой спирали, хотя известна и структура левой спирали.
Спиральные структуры белка.
Для полипептидных цепей известно несколько различных типов спиралей. Если при наблюдении вдоль оси спирали она удаляется от наблюдателя по часовой стрелке, то спираль считается правой (правозакрученной), а если удаляется против часовой стрелки — левой (левозакрученной). Наиболее распространена правая a-спираль (предложена Л. Полингом и Р. Кори). Идеальная a-спираль имеет шаг 0,54 нм и число однотипных атомов на один виток спирали 3,6. строение спирали стабилизируется внутримолекулярными водородными связями.
В природных белках существуют лишь правозакрученные a-спиральные конформации полипептидных цепей, что сопряжено с наличием в белковых телах аминокислот только L-ряда (за исключением особых случаев).
При растяжении a-кератина образуется вещество с другими свойствами - b-кератин. При растяжении спираль макромолекулы белка превращается в другую структуру , напоминающую линейную. Отдельные полипептидные цепи здесь связаны межмолекулярными водородными связями. Эта структура называется b-структурой ( структура складчатого листа, складчатого слоя)
Складчатые структуры белка.
Одним из распространенных примеров складчатой периодической структуры белка являются так называемые b-складки, состоящие из двух фрагментов, каждый из которых представлен полипептидом.
b-складки также стабилизируются водородными связями между атомом водорода аминной группы одного фрагмента и атомом кислорода карбоксильной группы другого фрагмента. При этом фрагменты могут иметь как параллельную, так и антипараллельную ориентацию относительно друг друга.
Для того чтобы два участка полипептидной цепи располагались в ориентации, благоприятствующей образованию b-складок, между ними должен существовать участок, имеющий структуру, резко отличающийся от периодической.
Возникновение a- и b-структур в белковой молекуле является следствием того, что аминокислоты и в составе полипептидных цепей сохраняют присущую им способность к образованию водородных связей. Таким образом, крайне важное свойство аминокислот — соединяться друг с другом водородными связями в процессе образования кристаллических препаратов — реализуется в виде a-спиральной конформации или b-структуры в белковой молекуле. Следовательно, возникновение указанных структур допустимо рассматривать как процесс кристаллизации участков полипептидной цепи в пределах одной и той же белковой молекулы.
Третичная структура Сведения о чередовании аминокислотных остатков в полипептидной цепи (первичная структура) и наличие в белковой молекуле спирализованных, слоистых и неупорядоченных ее фрагментов (вторичная структура) еще не дают полного представления ни об объеме, ни о форме, ни тем более о взаимном расположении участков полипептидной цепи по отношению друг к другу. Эти особенности строения белка выясняют при изучении его третичной структуры, под которой понимают — общее расположение в пространстве составляющих молекул одной или нескольких полипептидных цепей, соединенных ковалентными связями. То есть третичная конфигурация — реальная трехмерная конфигурация, которую принимает в пространстве закрученная спираль, которая в свою очередь свернута спиралью. У такой структуры в пространстве имеются выступы и впадины с обращенными наружу функциональными группами.
Сведения о чередовании аминокислотных остатков в полипептидной цепи (первичная структура) и наличие в белковой молекуле спирализованных, слоистых и неупорядоченных ее фрагментов (вторичная структура) еще не дают полного представления ни об объеме, ни о форме, ни тем более о взаимном расположении участков полипептидной цепи по отношению друг к другу. Эти особенности строения белка выясняют при изучении его третичной структуры, под которой понимают — общее расположение в пространстве составляющих молекул одной или нескольких полипептидных цепей, соединенных ковалентными связями. То есть третичная конфигурация — реальная трехмерная конфигурация, которую принимает в пространстве закрученная спираль, которая в свою очередь свернута спиралью. У такой структуры в пространстве имеются выступы и впадины с обращенными наружу функциональными группами.
Полное представление о третичной структуре дают координаты всех атомов белка. Благодаря огромным успехом рентгеноструктурного анализа такие данные, за исключением координат атомов водорода получены для значительного числа белков. Это огромные массивы информации, хранящиеся в специальных банках данных на машиночитаемых носителях, и их обработка немыслима без применения быстродействующих компьютеров. Полученные на компьютерах координаты атомов дают полную информацию о геометрии полипептидной цепи, что позволяет выявить спиральную структуру, b-складки или нерегулярные фрагменты.
Третичная структура формируется в результате нековалентных взаимодействий (электростатические, ионные, силы Ван-дер-Ваальса и др.) боковых радикалов, обрамляющих a-спирали и b-складки, и непериодических фрагментов полипептидной цепи. Среди связей, удерживающих третичную структуру, следует отметить:
а) дисульфидный мостик (–S–S–) между двумя остатками цистеина;
б) сложноэфирный мостик (между карбоксильной группой и гидроксильной группой);
в) солевой мостик (между карбоксильной группой и аминогруппой);
г) водородные связи между группами -СО - и -NH-;
Третичной структурой объясняется специфичность белковой молекулы, ее биологическая активность.
Первые пространственные модели молекул белка — миоглобина и гемоглобина — построили в конце 50-х гг. XX в. английские биохимики Джон Ко-удери Кендрю (родился в 1917 г.) и Макс Фердинанд Перуц (родился в 1914 г.). При этом они использовали данные экспериментов с рентгеновскими лучами. За исследования в области строения белков Кендрю и Перуц в 1962 г. были удостоены Нобелевской премии. А в конце столетия была определена третичная структура уже нескольких тысяч белков.
Четвертичная структура
У большинства белков пространственная организация заканчивается третичной структурой, но для некоторых белков с молекулярной массой больше 50-100 тысяч, построенных из несколько полипептидных цепей характерна четвертичная.
Сущность такой структуры в объединении несколько полимерных цепей были в единый комплекс. Такой комплекс также рассматривается как белок, состоящий из нескольких субъединиц. Белки, состоящие из нескольких субъединиц, широко распространены в природе (гемоглобин, вирус табачной мозаики, фосфорилаза, РНК-полимераза). Субъединицы принято обозначать греческими буквами (так у гемоглобина имеется по две a и b субъединицы). Наличие нескольких субъединиц важно в функциональном отношении — оно увеличивает степень насыщения кислородом.
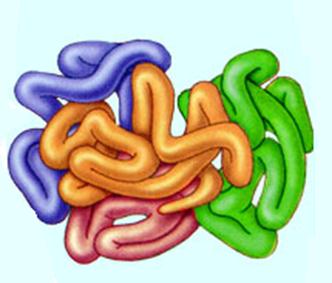 Четвертичная структура ( клубок белков)
Четвертичная структура ( клубок белков)
Четвертичная структура стабилизируется в основном силами слабых воздействий:
а) водородная; б) гидрофобная; в) ионные; г) ковалентные (дисульфидные, пептидные).
Денатурация белков
Денатурация белка — разрушение сил (связей), стабилизирующих четвертичную, третичную и вторичную структуры, приводящее к дезориентации конфигурации белковой молекулы и сопровождаемое изменением растворимости, вязкости, химической активности, характера рассеивания рентгеновских лучей, снижением или полной потерей биологической функции.
Различают физические (температура, давление, механическое воздействие, ультразвуковое и ионизирующее излучения) и химические (тяжелые металлы, кислоты, щелочи, органические растворители, алкалоиды) факторы, вызывающие денатурацию.
Обратным процессом является ренатурация, то есть восстановление физико-химических и биологических свойств белка. Иногда для этого достаточно удалить денатурирующий объект. Ренатурация невозможна если затронута первичная структура.
Химические и физические свойства
Несмотря на внешнее несходство, различные представители белков обладают некоторыми общими свойствами.
Так, поскольку все белки являются коллоидными частицами (размер молекул лежит в пределах 1 мкм до 1 нм), в воде они образуют коллоидные растворы. Эти растворы характеризуются высокой вязкостью, способностью рассеивать лучи видимого света, не проходят сквозь полупроницаемые мембраны.
Вязкость раствора зависит от молекулярной массы и концентрации растворенного вещества. Чем выше молекулярная масса, тем раствор более вязкий. Белки как высокомолекулярные соединения образуют вязкие растворы. Например, раствор яичного белка в воде.
Коллоидные частицы не проходят через полупроницаемые мембраны (целлофан, коллоидную пленку), так как их поры меньше коллоидных частиц. Непроницаемыми для белка являются все биологические мембраны. Это свойство белковых растворов широко используется в медицине и химии для очистки белковых препаратов от посторонних примесей. Такой процесс разделения называется диализом. Явление диализа лежит в основе действия аппарата “искусственная почка”, который широко используется в медицине для лечения острой почечной недостаточности.
Белки способны к набуханию, характеризуются оптической активностью и подвижностью в электрическом поле, некоторые растворимы в воде. Белки имеют изоэлектрическую точку.
Важнейшим свойством белков является их способность проявлять как кислые, так и основные свойства, то есть выступать в роли амфотерных электролитов. Это обеспечивается за счет различных диссоциирующих группировок, входящих в состав радикалов аминокислот. Например, кислотные свойства белку придают карбоксильные группы аспарагиновой и глутаминовой аминокислот, а щелочные — радикалы аргинина, лизина и гистидина. Чем больше дикарбоновых аминокислот содержится в белке, тем сильнее проявляются его кислотные свойства и наоборот.
Эти же группировки имеют и электрические заряды, формирующие общий заряд белковой молекулы. В белках, где преобладают аспарагиновая и глутаминовая аминокислоты, заряд белка будет отрицательным, избыток основных аминокислот придает положительный заряд белковой молекуле. Вследствие этого в электрическом поле белки будут передвигаться к катоду или аноду в зависимости от величины их общего заряда. Так, в щелочной среде (рН 7–14) белок отдает протон и заряжается отрицательно (движение к аноду), тогда как в кислой среде (рН 1–7)
подавляется диссоциация кислотных групп и белок становится катионом (движение к катоду):
![]()
![]()
![]() NH3+ кислая ср. NH3+
щелочная ср. NH2
NH3+ кислая ср. NH3+
щелочная ср. NH2
![]()
![]()
![]()
![]()
![]() R
R R
R
R R
COOH COO – COO –
Катион Амфион Анион
Таким образом, фактором, определяющим поведение белка как катиона или аниона, является реакция среды, которая определяется концентрацией водородных ионов и выражается величиной рН. Однако при определенных значениях рН число положительных и отрицательных зарядов уравнивается и молекула становится электронейтральной, то есть она не будет перемещаться в электрическом поле. Такое значение рН среды определяется как изоэлектрическая точка белков. При этом белок находится в наименее устойчивом состоянии и при незначительных изменениях рН в кислую или щелочную сторону легко выпадает в осадок. Для большинства природных белков изоэлектрическая точка находится в слабокислой среде (рН 4,8–5,4), что свидетельствует о преобладании в их составе дикарбоновых аминокислот.
Свойство амфотерности лежит в основе буферных свойств белков и их участии в регуляции рН крови. Величина рН крови человека отличается постоянством и находится в пределах 7,36–7,4 , несмотря на различные вещества кислого или основного характера, регулярно поступающие с пищей или образующиеся в обменных процессах, следовательно, существуют специальные механизмы регуляции кислотно-щелочного равновесия внутренней среды организма.
Белки активно вступают в химические реакции. Это свойство связано с тем, что аминокислоты, входящие в состав белков, содержат разные функциональные группы, способные реагировать с другими веществами. Важно, что такие взаимодействия происходят и внутри белковой молекулы, в результате чего образуется пептидная, водородная, дисульфидная и другие виды связей. К радикалам аминокислот, а, следовательно, и белков, могут присоединяться различные соединения и ионы.
Белки обладают большим сродством к воде, то есть они гидрофильны. Это значит, что молекулы белка, как заряженные частицы, притягивают к себе диполи воды, которые располагаются вокруг белковой молекулы и образуют водную или гидратную оболочку. Эта оболочка предохраняет молекулы белка от склеивания и выпадения в осадок. Величина гидратной оболочки зависит от структуры белка. Например, альбумины более легко связываются с молекулами воды и имеют относительно большую водную оболочку, тогда как глобулины, фибриноген присоединяют воду хуже, и гидратная оболочка и них меньше. Таким образом, устойчивость водного раствора белка определяется двумя факторами: наличием заряда белковой молекулы и находящейся вокруг нее водной оболочки. При удалении этих факторов белок выпадает в осадок. Данный процесс может быть обратимым и необратимым.
Обратимое осаждение белков (высаливание) предполагает выпадение белка в осадок под действием определенных веществ, после удаления которых он вновь возвращается в свое исходное (нативное) состояние. Для высаливания белков используют соли щелочных и щелочноземельных металлов (наиболее часто в практике используют сульфат натрия и аммония). Эти соли удаляют водную оболочку (вызывают обезвоживание) и снимают заряд. Между величиной водной оболочки белковых молекул и концентрацией солей существует прямая зависимость: чем меньше гидратная оболочка, тем меньше требуется солей. Так, глобулины, имеющие крупные и тяжелые молекулы и небольшую водную оболочку, выпадают в осадок при неполном насыщении раствора солями, а альбумины как более мелкие молекулы, окруженные большой водной оболочкой — при полном насыщении.
Необратимое осаждение связано с глубокими внутримолекулярными изменениями структуры белка, что приводит в потере ими нативных свойств — денатурации , которая влечет потерю растворимости, биологической активности и т.д. Необратимое осаждение можно вызвать кипячением, действием концентрированными растворами некоторых из минеральных и органических кислот, солями тяжелых металлов . Примером естественно вызванной денатурации служит расщепление белков в желудке, где имеется сильнокислая среда (рН 0,5–1,5), под действием протеолитических ферментов. Денатурация белков положена в основу лечения отравления тяжелыми металлами, когда больному вводят per os (“через рот”) молоко или сырые яйца с тем, чтобы металлы адсорбировались на поверхности денатурирующего белка и не действовали на белки слизистой оболочки желудка и кишечника, а также не всасывались в кровь.
Гидролиз белка достигается при помощи кипячения белка с сильными минеральными кислотами (кислотный гидролиз) или основаниями (щелочной гидролиз). Схема следующая:
О H О Н О О
![]()
![]() NH2 — СН—С—N—СH—С—N—СН—С— ·· + nH2O ·· + NH2—СН—С—ОН +
NH2 — СН—С—N—СH—С—N—СН—С— ·· + nH2O ·· + NH2—СН—С—ОН +
R1 R2 R3 R1
O O
+ NH2—СН—С—ОН + NH2—СН—С—ОН + ··
R2 R3
Химический синтез
Химический синтез белков имеет большое практическое и теоретическое значение. В практическом отношении важны белковые гормоны — инсулин и вазопрессин, в настоящее время получаемые синтетическим путем. Умение производить искусственным путем необходимые белки откроет огромные ресурсы для использования в медицине, технике и т.д.
Традиционные методы синтеза регулярных полимеров позволяют получить сополимеры, состоящие из двух (или более) сходных типов мономеров со статистическим распределением их по цепи, в том числе белков. В частности, возможно получение гомополимеров или статистических сополимеров, состоящих из аминокислотных остатков, связанных пептидными связями (полиаминокислот).
В качестве примера можно привести процесс получения полиаминокислот, основанный на конденсации N-карбоксиангидридов аминокислот, образуемых из соответствующих аминокислот обработкой фосгеном:
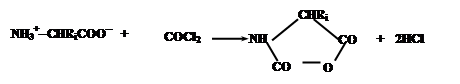
Эти соединения содержат электрофильную ангидридную группу, которая может атаковать алифатическую аминогруппу аминокислоты, используемой в качестве затравки, с выделением СО2 и одновременном освобождением новой аминогруппы из атакующей молекулы N-карбоксиангидрида, таким образом, открывая возможность поликонденсации:
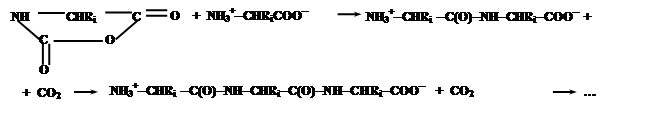
Нетрудно заметить, что каждая стадия поликонденсации (с учетом реакции образования N-карбоксиангидридов аминокислот) сопровождается превращением молекулы COCl2 в CO2 и 2HCl, что термодинамически выгодно и является источником свободной энергии для образования пептидной связи.
При синтезе нерегулярных полипептидов базируются также на активации карбоксильных групп. Большинство из них базируется на использовании N,N-дициклогексилкарбодиимида (ДЦК). Он способен в присутствии RCOO- и амина NH2R’ осуществить активацию карбоксильных групп:
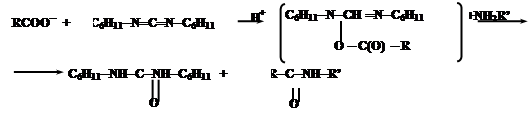
Промежуточным соединением является O-ацил-N,N’-дициклогексилмочевину (ДЦМ):
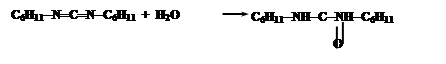
Значение белков
Функции белков чрезвычайно многообразны. Каждый данный белок как вещество с определенным химическим строением выполняет одну узкоспециализированную функцию и лишь в нескольких отдельных случаях — несколько взаимосвязанных. Например, гормон мозгового слоя надпочечников адреналин, поступая в кровь, повышает потребление кислорода и артериальное давление, содержание сахара в крови, стимулирует обмен веществ, а также является медиатором нервной системы у холоднокровных животных.




![]() Белки
Белки
 | |||
 | |||
Схема практического значения белков
.
Каталитическая (ферментативная) функция
Многочисленные биохимические реакции в живых организмах протекают в мягких условиях при температурах, близких к 40°С, и значениях рН близких к нейтральным. В этих условиях скорости протекания большинства реакций ничтожно малы, поэтому для их приемлемого осуществления необходимы специальные биологические катализаторы — ферменты. Даже такая простая реакция, как дегидратация угольной кислоты:
![]()
![]() CO2 + H2O HCO3-+ H+
CO2 + H2O HCO3-+ H+
катализируется ферментом карбоангидразой. Вообще все реакции, за исключением реакции фотолиза воды 2H2O®4H+ + 4e- + O2, в живых организмах катализируются ферментами (реакции синтеза, осуществляются при помощи ферментов синтетаз, реакции гидролиза — при помощи гидролаз, окисление — при помощи оксидаз, восстановление с присоединением — при помощи гидрогеназ и т.д.). Как правило, ферменты — это либо белки, либо комплексы белков с каким-либо кофактором — ионом металла или специальной органической молекулой. Ферменты обладают высокой, иногда уникальной, избирательностью действия. Например, ферменты, катализирующие присоединение a-аминокислот к соответствующим т-РНК в процессе биосинтеза белка, катализируют присоединение только L-аминокислот и не катализируют присоединение D-аминокислот.
Транспортная функция белков
Внутрь клетки должны поступать многочисленные вещества, обеспечивающие ее строительным материалом и энергией. В то же время все биологические мембраны построены по единому принципу — двойной слой липидов, в который погружены различные белки, причем гидрофильные участки макромолекул сосредоточены на поверхности мембран, а гидрофобные “хвосты” — в толще мембраны. Данная структура непроницаема для таких важных компонентов, как сахара, аминокислоты, ионы щелочных металлов. Их проникновение внутрь клетки осуществляется с помощью специальных транспортных белков, вмонтированных в мембрану клеток. Например, у бактерий имеется специальный белок, обеспечивающий перенос через наружную мембрану молочного сахара — лактозы. Лактоза по международной номенклатуре обозначается b-галаткозид, поэтому транспортный белок называют b-галактозидпермеазой.
Важным примером транспорта веществ через биологические мембраны против градиента концентрации является К/ Na-ый насос. В ходе его работы происходит перенос трех положительных ионов Na+ из клетки на каждые два положительных иона K+ в клетку. Эта работа сопровождается накоплением электрической разности потенциалов на мембране клетки. При этом расщепляется АТФ, давая энергию. Молекулярная основа натрий-калиевого насоса была открыта недавно, это оказался фермент, расщепляющий АТФ — калий-натрийзависимая АТФ-аза.
У многоклеточных организмов существует система транспорта веществ от одних органов к другим. В первую очередь это гемоглобин. Кроме того, в плазме крови постоянно находится транспортный белок — сывороточный альбумин. Этот белок обладает уникальной способностью образовывать прочный комплексы с жирными кислотами, образующимися при переваривании жиров, с некоторыми гидрофобными аминокислотами со стероидными гормонами, а также со многими лекарственными препаратами, такими, как аспирин, сульфаниламиды, некоторые пенициллины.
Рецепторная функция
Большое значение, в особенности для функционирования многоклеточных организмов, имеют белки-рецепторы, вмонтированные в плазматическую мембрану клеток и служащие для восприятия и преобразования различных сигналов, поступающих в клетку, как от окружающей среды, так и от других клеток. В качестве наиболее исследованных можно привести рецепторы ацетилхолина, находящиеся на мембране клеток в ряде межнейронных контактов, в том числе в коре головного мозга, и у нервно-мышечных соединений. Эти белки специфично взаимодействуют с ацетилхолином CH3C(O) – OCH2CH2N+(CH3)3 и отвечает на это передачей сигнала внутрь клетки. После получения и преобразования сигнала нейромедиатор должен быть удален, чтобы клетка подготовилась к восприятию следующего сигнала. Для этого служит специальный фермент — ацетилхолинэстераза, катализирующая гидролиз ацетилхолина до ацетата и холина.
Многие гормоны не проникают внутрь клеток-мишеней, а связываются со специфическими рецепторами на поверхности этих клеток. Такое связывание является сигналом, запускающим в клетке физиологические процессы.
Защитная функция
Иммунная система обладает способностью отвечать на появление чужеродных частиц выработкой огромного числа лимфоцитов, способных специфически повреждать именно эти частицы, которыми могут быть чужеродные клетки, например патогенные бактерии, раковые клетки, надмолекулярные частицы, такие как вирусы, макромолекулы, включая чужеродные белки. Одна из групп лимфоцитов — В-лимфоциты, вырабатывает особые белки, выделяемые в кровеносную систему, которые узнают чужеродные частицы, образуя при этом высокоспецифичный комплекс на этой стадии уничтожения. Эти белки называются иммуноглобулины. Чужеродные вещества, вызывающие иммунный ответ называют антигенами, а соответствующие к ним иммуноглобулины — антителами.
Антитела построены из четырех полипептидных цепей, связанных между собой дисульфидными мостиками.
Структурная функции
Наряду с белками, выполняющими тонкие высокоспециализированные функции, существуют белки, имеющие в основном структурное значение. Они обеспечивают механическую прочность и другие механические свойства отдельных тканей живых организмов. В первую очередь это коллаген — основной белковый компонент внеклеточного матрикса соединительной ткани.
В эластичных тканях — коже, стенках кровеносных сосудов, легких - помимо коллагена внеклеточный матрикс содержит белок эластин, способный довольно в широких пределах растягиваться и возвращаться в исходное состояние.
Еще один пример структурного белка — фиброин шелка, выделяемый гусеницами шелкопряда в период формирования куколки и являющийся основным компонентом шелковых нитей.
Двигательные белки
Мышечное сокращение является процессом, в ходе которого происходит превращение химической энергии, запасенной в виде макроэргических пирофосфатных связей в молекулах АТФ, в механическую работу. Непосредственными участниками процесса сокращения являются два белка — актин и миозин.
Антибиотики
Большую и чрезвычайно важную в практическом отношении группу природных органических соединений составляют антибиотики — вещества микробного происхождения, выделяемые специальными видами микроорганизмов и подавляющие рост других, конкурирующих микроорганизмов. Открытие и применение антибиотиков произвело в 40-ые гг. революцию в лечении инфекционных заболеваний, вызываемых бактериями. Следует отметить, что на вирусы в большинстве случаев антибиотики не действуют и применение их в качестве противовирусных препаратов неэффективно.
Токсины
Ряд живых организмов в качестве защиты от потенциальных врагов вырабатывают сильно ядовитые вещества — токсины. Многие из них являются белками, однако, встречаются среди них и сложные низкомолекулярные органические молекулы. В качестве примера такого вещества можно привести ядовитое начало бледной поганки — a-аманитин.
Вывод:
В данной работе при помощи различных схем и таблиц были рассмотрены химические и физические свойства белков, классификация белков, состав и строение белков, были рассмотрены разнообразные функции белков, а также их значение.
Доказано, что белки - обязательная составная часть всех живых клеток, играют исключительно важную роль в живой природе, являются главным, наиболее ценным и незаменимым компонентом питания. Это связанно с той огромной ролью, которую они играют в процессах развития и жизни человека. Белки являются основой структурных элементов и тканей, поддерживают обмен веществ и энергии, участвуют в процессах роста и размножения, обеспечивают механизмы движений, развитие иммунных реакций, необходимы для функционирования всех органов и систем организма.
"Жизнь - это форма существования белка"
 Список использованной литературы:
Список использованной литературы:
· «ХИМИЯ—справочник для абитуриентов и студентов». Издательство acT-Фолио, Москва, 2000 год.
· Большая медицинская энциклопедия.
· «Энциклопедия для детей. Химия». Аванта+, Москва, 2000 год.
· Албертс Б., Брей Д., и др. Молекулярная биология клетки Москва, 1994.
· Биотехнология. Производство белковых веществ. В.А. Быков, М.Н. Манаков . Москва «Высшая школа» 1987г.
· Артеменко А.И. Органическая химия: учеб. для строит. спец. вузов. —М.: Высшая школа, 2000.
· Березин Б.Д., Березин Д.Б. Курс современной органической химии. Учебное пособие для вузов. —М.: Высшая школа, 1999.
· Кнорре Д.Г., Мызина С.Д. Биологическая химия. —М.: Высшая школа, 1998.
· Общая органическая химия. Под ред. Д. Бартона, У.Д. Оллиса. Нуклеиновые кислоты, аминокислоты, петиды, белки. —М.: Химия, 1986.
· Филлпович Ю.Б. Основы биохимии: уч. для студ. хим. и биол. спец. пед. инст. М.:Высшая школа, 1985.
· Шамин А.Н. История химии белка. —Москва : «Наука», 1977.
· Якубке Х.-Д., Ешкайт Х. Аминокислоты, пептиды, белки. Москва: «Мир», 1985.
Похожие работы

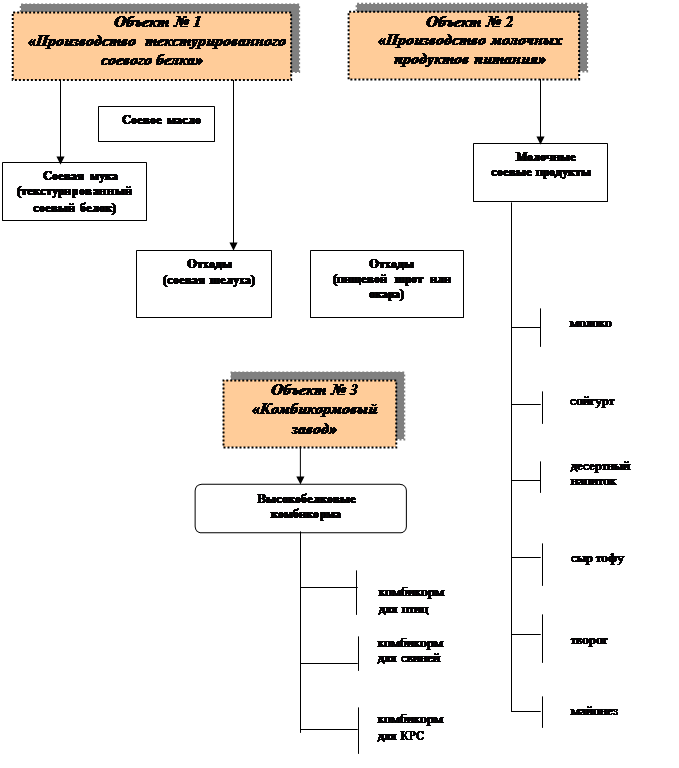
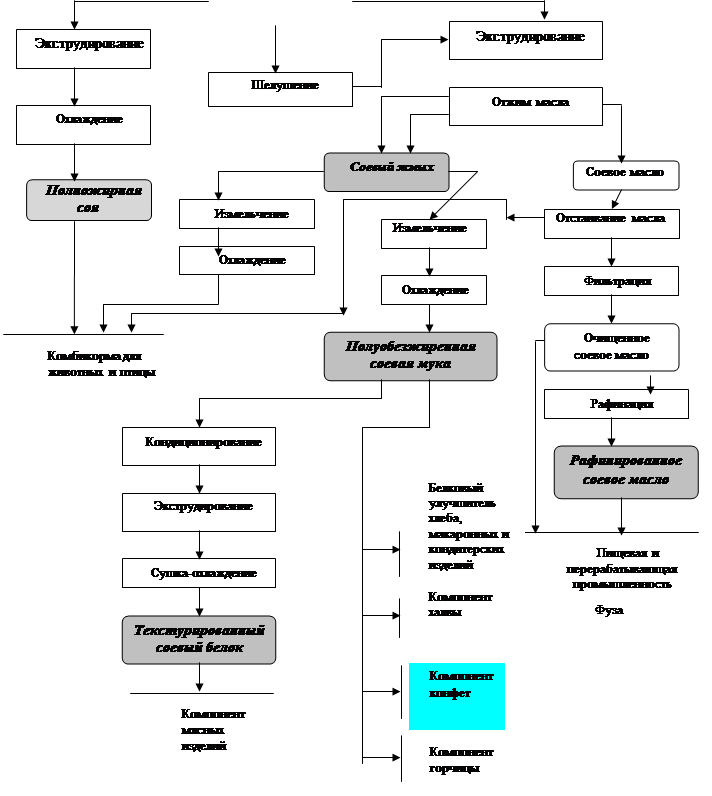
... и текстураты соевых белков являются наиболее распространенными производным сои с широко применяются в пищевой промышленности и производстве специализированных питательных смесей. Финансовая модель Оценка эффективности инвестиций в проект, являющийся предметом настоящего бизнес-плана, а также способности заемщика обслужить инвестиционный кредит, осуществлена с помощью финансовой модели. Периодом ...
... на 1/3 сильно пострадали от больших пожаров, то есть из 90 тыс. га лесной площади. 30 тыс. га пройдено верховыми пожарами. Хвойные насаждения сменены на лиственные. Место обитания белки обыкновенной на территории Унгутского лесничества в основном в насаждениях, прилегающих к реке Мана в 1 группе леса, т.к. смешанные кедрачи с примесью разных деревьев остались не тронутыми рубкой и уцелели от ...
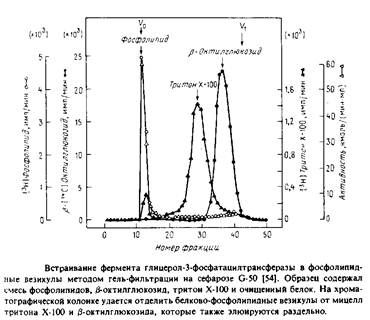
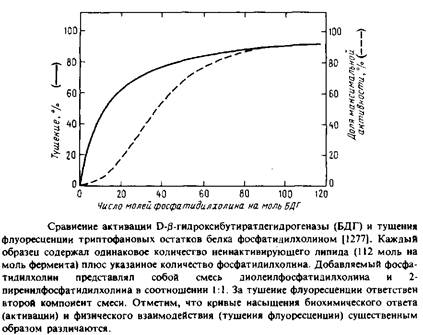
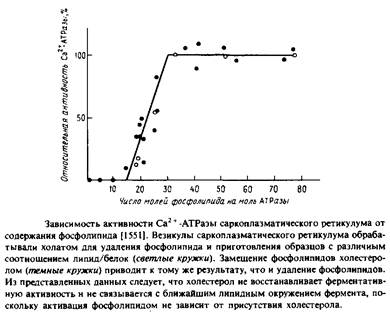
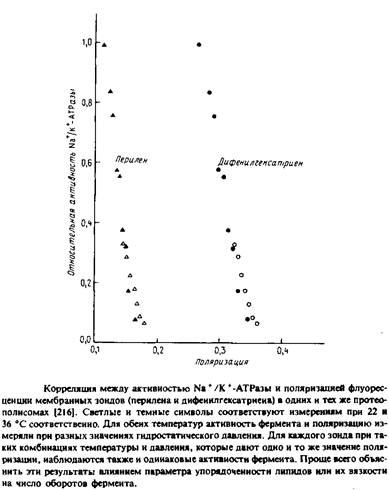
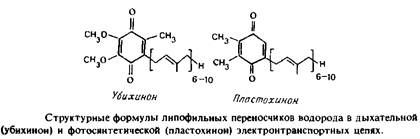
... . Все четыре системы являются основными белковыми компонентами мембран, в которых они локализованы, и могут служить иллюстрацией различных уровней организации мультиферментных комплексов в мембранной энзимологии. Первые две системы катализируют анаболические и катаболические реакции, протекающие в присутствии молекулярного кислорода и обычно липофильных мембраносвязанных субстратов. Терминальные ...
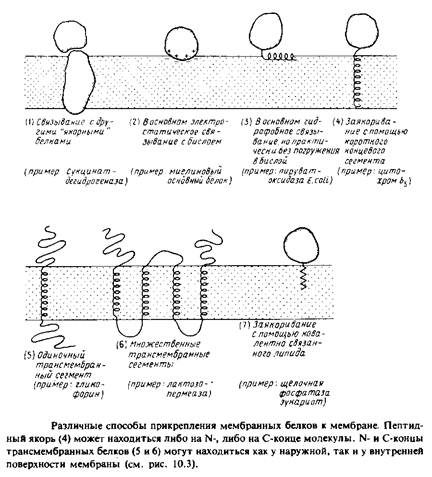

... и D2O, но анализ результатов в этом случае гораздо сложнее, хотя принципиально не отличается от предыдущего случая. Альтернативный способ определения молекулярной массы нативной формы мембранного белка состоит в равновесном ультрацентрифугировании. Распределение вещества в состоянии равновесия таково, что наклон графика зависимости логарифма концентрации от г2 равен где г — расстояние от ...
0 комментариев