Уральская Государственная Медицинская Академия.
Кафедра биоорганической
и биологической химии
Курсовая работа по теме:
Переваривание и всасывание липидов.
Исполнитель: студентка педиатрического
факультета 223 группы
Комова М. О.
Руководитель: доцент, к.м.н. Трубачев С. Д.
Рецензент:
Екатеринбург 2002.
Содержание.I. Введение……………………………………………………………….3
II. Определение класса липидов, их классификация и биологическое
значение каждого класса…………………………………………...3
III. Принципы нормирования и возрастные нормы липидов в
питании……………………………………………………….……..5
IV. Этапы обмена липидов в организме…………………………………6
V. Липипротеиды………………………………………………………...7
1. Строение и химический состав………………………………72. Классификация ЛП……………………………………………9
3. Роль липопротеинов…………………………………………12 4. Наследственная недостаточность липопротеидов…………12VI. Переваривание и всасывание липидов…………………………….12
1. Желчь…………………………………………………………12 · Значение……………………………………………..12· Последствия нарушения секреции………………...14
· Химический состав…………………………………15 · Гуморальная регуляция секреции…………………16 2.ПАВ желудочно-кишечного тракта и механизмэмульгирования, значение………………………………..18
3. Расщепление липидов……………………………………….19 · ТГ…………………………………………………….19 · ФЛ……………………………………………………22 · ХС…………………………………………………….23 4. Химический состав и строение мицелл, механизмывсасывания липидов……………………………………..23
5. Механизм ресинтеза липидов в энтероцитах, значение…..26 6. Образование и обмен ХМ, значение ……………………….30 VII. Нарушения переваривания и всасывания липидов……………….34 1. Стеаторея……………………………………………………..34 2. Хиломикронемия…………………………………………….35 VIII. Заключение………………………………………………………….36IX. Приложение…………………………………………………………37
X. Список литературы…………………………………………………40
Введение. Уже при кратком знакомстве с молекулярными основами жизни мы сталкиваемся с липидами. Назовем их основные биологические свойства: Главные компоненты биологических мембран; Запасной, изолирующий и защищающий органы материал; Наиболее калорийная часть пищи; Важная составная часть диеты человека и животных; Переносчики ряда витаминов; Регуляторы транспорта витаминов и солей; Иммуномодуляторы; Регуляторы активности некоторых ферментов; Эндогормоны; Передатчики биологических сигналов.Этот список увеличивается по мере изучения липидов. В обеспечении названных и других функций участвуют липиды различной структуры в разных количествах: тонны триглицеридов служат китам как запас энергии и защита тела от внешних воздействий, а как эндогормоны или передатчики биологических сигналов действуют липиды других классов в микро- и нанограммовых дозах. Поэтому для понимания сути многих биологических процессов нужно иметь представления о переваривании и всасывании липидов, об их транспорте и синтезе в организме.
Определение класса липидов, их классификация и
биологическое значение .
В учебнике по общей химии под редакцией Ю. И. Полянского сказано: “Липиды представляют собой органические вещества, нерастворимые в воде, но растворимые в бензоле, эфире, ацетоне.” Сходные определения липидов чаще всего встречаются и в одном из лучших руководств по биохимии. Они имеют два существенных недостатка: во – первых, вместо четкой химической характеристики класса говорят о физических свойствах липидов, во – вторых, содержат фактические ошибки. Так, далеко не все липиды растворимы в перечисляемых органических растворителях. Н. Грин с соавторами, с одной стороны критикуют подобные определения, но с другой – не доводят дело до конца: “ Можно все же сказать, что настоящие липиды – это сложные эфиры жирных кислот и какого – либо спирта”. Как мы увидим, помимо сложных эфиров спиртов есть много других липидов. Неправильные определения влекут за собой запутанные, неверные классификации . В число липидов часто включают стерины, жирорастворимые витамины и другие соединения. Мы будем относить к липидам вещества с четко выраженной химической структурой, тесно связанные биохимически: липиды – это жирные кислоты и их производные. Что такое жирные кислоты? Из органической химии известно, что это алифатические монокарбоновые кислоты R – СООН. Как и для других классов природных соединений, определение наполнится глубоким содержанием после знакомства с главными представителями липидов [1, 1997]. Липиды разделяются на две группы по принципу гидролитического расщепления. Первая – липиды, не подвергающиеся гидролизу. К ним можно отнести некоторые углеводороды , например, сквален и картиноиды, высшие спирты, включая стерины, и высшие аминоспирты, высшие альдегиды, кетоны и хиноны ( витамины группы К, убихинон и т.д. ) , жирные кислоты (ЖК) и простогландины (ПГ). Во вторую группу включены липиды, гидролиз которых приводит к “освобождению” двух и более индивидуальных соединений. В эту группу входят в основном вещества, содержащие сложноэфирную и / или амидную связи, а также связь типа простого эфира, ацеталя или полуацеталя. Это – воски, эфиры стеринов, в том числе холестерина (ХС) и многоатомных спиртов (например, глицериды, фосфолипиды (ФЛ), включая сфиегомиелины ), гликолипиды, серусодержащие липиды и липиды, имеющие в своем составе аминокислоты.Если оставить в стороне ряд соединений, которые по отдельным признакам подходят к определению “липиды” или являются их предшественниками (например , жирные кислоты, сквален и др.) или производными (например, ПГ), то можно использовать следующую классификацию липидов, основанную на их структурных особенностях:
n глицериды;
n воски;
n ФЛ: глицерофосфолипиды , сфингомиелины;
n гликолипиды (гликосфинголипиды) : цереброзиды и ганглиозиды ;
n другие сложные липиды ( сульфолипиды и аминолипиды);
n стерины и их эфиры с ЖК.
Биологическое значение.
Воска: У позвоночных воски, секретируемые кожными железами, выполняют функцию защитного покрытия, смазающего и смягчающего кожу и предохраняющего ее от воды. Восковым секретом покрыты даже волосы. Перья птиц , особенно водоплавающих, и шкура животных имеют восковое покрытие, которое придает водоотталкивающие свойства. Воск овечьей шерсти, называемый линолином, в качестве спиртовой компоненты содержит ланостерин – один из конечных продуктов биосинтеза холестерина. Ланолин широко используется в медицине и косметике как основа для приготовления различных мазей и кремов.
Цереброзиды обнаруживаются главным образом в миелиновых оболочках и в мембранах нервных клеток мозга.
Ганглиозиды: Они найдены в сером веществе головного мозга. Локализованы в плазматических мембранах нервных клеток, где на их долю приходится около 6 % мембранных липидов. В меньшем количестве они обнаружены в мембранах клеток других тканей. Показано участие ганглиозидов в формировании защитного слоя клеток – гликокаликса и в осуществлении ими рецепторной функции.
ФЛ обнаружены в составе тканей и клеток всех живых существ, как в свободном виде, так и в виде белково – липидных комплексов (липопротеидов и протеолипидов) . Особенно много ФЛ содержится в оболочках и мембранах клеток и клеточных органелл (ядра, митохондрий и микросомах), где они образуют структурную основу мембраны – фосфолипидный бислой. Наиболее богаты ФЛ ткани мозга и нервов (до 30 % в пересчете на сухую массу ткани), печень(до 16 %), почки(до11%), сердце(до 10 %), скелетные мышцы (около 3 %). В плазме крови человека содержится 2,8 – 4,4 ммоль / л ФЛ.
Всюду, где содержатся ФЛ им сопутствует холестерин. Поэтому эти липиды иногда называют комплементарными.
Стерины и их эфиры с жирными кислотами: Наиболее важным представителем этого класса соединений является ХС. Каждая клетка в организме млекопитающих содержит ХС входя в состав мембранных клеток , НЭХС вместе с ФЛ и белками обеспечивает избирательную проницаемость клеточной мембраны и оказывает регулирующее влияние на состояние мембраны и на активность связанных с ней энзимов.
ХС является источником образования в организме млекопитающих желчных кислот, а также стероидных гормонов: тестостерона, эстрадиола, прогестерона, кортизоном, альдестерона. ХС, а точнее продукты его окисления 7-дегидрохолестерин, в результате воздействия УФ-лучей на кожу превращается в ней в витамин D3. Таким образом физиологическая функция ХС многообразна [5,1999].
Глицериды. ТГ составляют основную массу резервных липидов человеческого организма. Они выполняют резервную функцию, причем это преимущественно энергетический резерв организма. У человека массой 70 кг на долю резервных липидов приходится примерно 11 кг. Учитывая калорический коэффициент для липидов, равный 9,3 ккал/г, общий запас энергии в резервных ТГ составляет величину порядка 100000 ккал. Функция резервных ТГ как запаса пластического материала не столь очевидна, но все же продукты расщепления ТГ могут использоваться для биосинтезов, например, входящий в их состав глицерол может быть использаван для синтеза глюкозы или некоторых аминокислот.
Являясь одним из основных компонентов жировой ткани, ТГ участвуют в защите внутренних органов человека от механических повреждений. Кроме того , входя в большом количестве в состав подкожной жировой клетчатки, они участвуют втерморегуляции, образуя теплоизолирующую прослойку [6, 1999].
Принципы нормирования и возрастные нормы в питании.
Липиды – основные пищевые вещества, покрывающие 35% энергозатрат в организме человека [10, 2001]. Пищевой рацион должен содержать липиды из расчета 1,5 г на 1 кг массы тела, что составляет для 70-килограммового человека около 100 г липидов в сутки [6, 1999].
При нормировании количества пищи необходимо учитывать:
n вес (масса) тела;
n возраст;
n образ жизни;
n состояние организма [10, 2001].
Важно, чтобы соблюдалось оптимальное соотношение животных и растительных жиров. Оно должно составлять 70:30 .Потребность организм в растительных маслах, где в основном содержатся незаменимые жирные кислоты, равняется 25 – 30 граммам в сутки [9, 1994].
Педиатрическая диетология , или диетология развития, в течение уже нескольких десятков лет используют в качестве надежного ориентира в основании уровней потребления и рекомендации , факты, относящиеся к химическому составу и использованию грудным ребенком нутриентов “золотого стандарта” пищевого обеспечения.
Физиологические потребности в нутриентах расчитывают, исходя из результатов анализа реальных величин потребления в представительных выборках детей грудного возраста, а также на основе использования различных клинических, общепедиатрических и биохимических критериев степени обеспеченности при разных уровнях потребления. Во многих странах существуют государственные рекомендации по нормированию потреблений. Термин “ рекомендуемая норма потребления ’’ содержит в себе и некоторую гарантирующую избыточность или “ резерв надежности” рекомендации ориентированный на гетерогенность популяции. Естественно, что рекомендуемые нормы изменяются по мере накопления объективных подтверждений их необходимости. Решающее значение при этом имеют клинические и эпидемиологические данные об особенностях развития и здоровья детей при том или ином уровне поступления нутриента. Ниже приведены примеры нормирования питания на уровне ВОЗ [7,1999].
| 0–2мес | 3–5мес | 6–11мес. | 1–3г. | 3–7л. | 7–10л. | 11 – 13л. | 14-17л. | |
| Жиры, всего, г | ¾ | ¾ | ¾ | 53 | 68 | 79 | 93(м)¤85(д) | 100 ¤ 90 |
| В том числе Растительные, г | ¾ | ¾ | ¾ | 5 - 10 | 11 | 16 | 19(м)¤17(д) | 20 ¤ 18 |
| Жиры, г¤кг | 6,5 | 6,0 | 5,5 | ¾ | ¾ | ¾ | ¾ | ¾ |
Этапы обмена липидов в организме.
Липиды, поступающие с пищей, крайне гетерогенны по своему происхождению. В желудочно кишечном тракте они в значительной мере расщепляются до составляющих мономеров: высших жирных кислот, глицерола, аминоспиртов и др. Эти продукты расщепления всасываются в кишечную стенку и из них в клетках кишечного эпителия синтезируются липиды, свойственные человеку. Эти видоспецифические липиды далее поступают в лимфатическую и кровеносную системы и разносятся к различным тканям и органам [6, 1999].
Липопротеиды.
Строение и химический состав.
Исходя из современных представлений, само понятие “липопротеиды” можно определить следующим образом: липопротеиды (ЛП) – высоко молекулярные водорастворимые частицы, представляющие собой комплекс белка и липида, образованный нековалентными связями, в котором белки совместно с полярными липидами формируют поверхностный гидрофильный слой, окружающий и защищающий внутреннюю гидрофобную липидную сферу от водной сферы и обеспечивающий транспорт липидов в кровяном русле и доставку их в органы и ткани. Согласно этому определению, одним из признаков ЛП является наличие в них наружного гидрофильного белково – липидного слоя и липидной гидрофобной сферы (ядра).
Плазменные ЛП-частицы имеют сферическую форму. Внутри находится жировая капля, содержащяя неполярные липиды (триглицериды и эстерефицированный холестерин) и формирующая ядро ЛП-частицы. Оно окружено оболочкой из ФЛ, НЭХС и белка. Целесообразность такой структуры объясняется тем, что неполярные липиды нерастворимы в водной среде и поэтому не могут транспортироваться в ток крови. Полярные же липиды (ФЛ, НЭХС) совместно с белком формируют поверхностный гидрофильный слой, который с одной стороны, защищает внутреннюю гидрофобную липидную сферу от водной среды, а с другой – обеспечивает растворимость и транспорт ЛП-частицы в этой же водной среде. ФЛ и НЭХС покрывают только 30 – 70 % поверхности частицы, остальную ее часть восполняет белок.
Основную массу ЛП-частицы составляет ее ядро, в котором помимо ТГ и ЭХС, обнаруживаются небольшие количества НЭХС. Именно ядро частицы определяет ее размеры и сферическую форму. В зависимости от класса ЛП изменяется соотношение между основными липидами: с увеличением плотности частиц уменьшается доля ТГ и возрастает доля ЭХС. Поскольку ТГ являются растворителями для последних, то в богатых ТГ липид – белковых комплексах (ХМ и ЛПОНП) эфиры ХС равномерно распределены по ядру, тогда как в ЛПНП и ЛПВП они образуют отдельные скопления. Образно , к ядру ЛП-частицы можно употребить выражение “липиды внутри липида ”. Наружная оболочка ЛП-частицы, в отличии от ядра, обладает относительно высокой электронной плотностью. Толщина этой оболочки составляет 2,1 – 2,2 нм, что соответствует половине толщины липидного бислоя клеточных мембран. Отсюда было сделано заключение , что в плазменных ЛП наружная оболочка, в отличии от клеточных мембран, содержит липидный монослой. ФЛ, а также НЭХС расположены в наружной оболочке таким образом, что их полярные группы ориентированны наружу, а гидрофобные жирно – кислотные “хвосты” – внутрь частицы, причем какая-то часть этих “хвостов” даже погружена в липидное ядро.
По всей вероятности , наружная оболочка ЛП представляет собой не гомогенный слой, а мозаичную поверхность с выступающими участками белка и , возможно, НЭХС. Именно такая структура делает ЛП-частицу менее обособленной по сравнению с клеткой, окруженной бислойной мембраной, и объясняет легкую подвижность НЭХС (в меньшей степени белка и ФЛ) и способность этих компонентов переходить из одного класса ЛП на другой, даже сердцевинно-расположенные ЭХС и ТГ могут переходить из ЛП-частиц одной плотности на ЛП-частицы другой.
Существует много различных схем строения ЛП-частицы. Предполагается , что входящие в ее состав белки занимают только часть наружной оболочки. На основании данных , полученных при изучении переноса энергии с остатков белка одного из классов ЛП (ЛПНП) на гидрофобный слой пирен , было сделано заключение, что глубина погружения триптофанилов в фосфолипидный монослой составляет всего лишь 1,16 ± 0,26 нм. Вместе с тем, допускается, что значительная часть каждой белковой молекулы погружены в ЛП-частицу глубже, чем толщина ее наружной оболочки. В целом положение белков в ЛП-частице напоминает картину белкового “айсберга”, плавающего в “липидном море”, предложенную ранее для объяснения структуры клеточных мембран.(рис. 1)
Схема строения ЛП-частицы имеет сходство со структурой плазматической мембраны. Некоторое количество ЭХС и ТГ (не показано) содержится в поверхностном слое, а в ядре частицы имеется небольшое количество НЭХС.
Такая структура может обеспечивать непосредственный контакт белковых молекул с липидами. Отдельные белки (апопротеины), входящие в состав ЛП , выполнят коэнзимную функцию в таких реакциях , как эстерификация ХС и гидролиз ТГ, протекающих непосредственно на ЛП-частице. Это требует прямого контакта липидов с апопротеинами и соответствующими энзимами [5, 1999]. Апопротеины обеспечивают растворимость ЛП и (благодаря их сигнальной роли) определяют пути метаболизма и судьбу каждого класса ЛП-частиц [3, 2000].
Липиды оболочки ЛП-частицы обладают более высокой микровязкостью, чем липиды ядра. Микровязкость липидов увеличивается , если в оболочке увеличивается содержание НЭХС, а в сердцевине – содержание ЭХС и ТГ с насыщенными ЖК. Увеличение микровязкости липидов может наблюдаться при скармливании животным ХС, а ее снижение – при содержании на диете , богатой полиненасыщенными ЖК. Микровязкость липидов , особенно оболочки ЛП-частицы , играет определенную роль в ее взаимодействии с мембраной клеток. В целом интегральность структуры ЛП-частицы обеспечивается гидрофобными , и в большей степени, ионными связями; при этом имеют место следующие взаимодействия: липид – липид, липид – белок, белок – белок.
В связи с тем, что плазменные ЛП представляют собой сложные надмолекулярные комплексы, в которых связи между компонентами комплекса носят нековалентный характер, применительна к ним вместо слова “молекула” употребляют выражение “частица”.
Классификация ЛП.
Существует несколько классификаций ЛП, основанных на различиях в их свойствах: гидратированной плотности, скорости флотации, электрофлоретической подвижности, а так же на различиях в апопротеиновом составе. Наибольшее распространение получила классификация, основанная на поведении отдельных ЛП в гравитационном поле в процессе ультрацентрифугирования. Гидратированная плотность ЛП колеблется в пределах 0,93 – 1,16 гр ¤ мл, что ниже гидратированной плотности плазменных белков, не связанных с липидами. Поэтому при ультрацентрифугировании в растворах с солевой плотностью, равной 1,21 или 1,25 г ¤ мл, ЛП всплывают, а белки, неассоциированные с липидами, остаются в инфрантанте.
При аналитическом ультрацентрифугировании разделения ЛП на фракции основано на скорости их флотации при плотности раствора 1,063 г¤мл для ХМ (Sf >400), ЛПОНП (Sf 20 – 400),и ЛПНП (Sf 0 – 20) и при плотности равной 1,20 г/мл для ЛПВП.
Различная электрофоретическая подвижность по отношению к глобулинам плазмы положена в основу другой классификации ЛП согласно которой различают ХМ (остаются на старте подобно g-глобулинам), b-ЛП (ЛПНП), пре-b-ЛП (ЛПОНП) и a-ЛП (ЛПВП), занимающие положение b-, a1-, a2-глобулинов соответственно.
Приведенные выше классификации не учитывают то обстоятельство, что каждый из классов ЛП отличается большой дисперсностью и гетерогенностью. Последнего недостатка в значительной степени лишена так называемая химическая классификация ЛП, основанная на оценке состава апопротеинов как специфических маркеров для рассматриваемых липид – белковых комплексов.
Данный подход и классификация ЛП предусматривает деление всех ЛП на первичные и вторичные (ассоциированные комплексы). К первичным относятся такие ЛП, которые содержат один индивидуальный белок – апопротеин (например, ЛП В-100, ЛП С-I, ЛП С-II и т.д.). Ко вторым ЛП относят ассоциаты первичных ЛП (например,ЛП А-I : А-II, ЛП А-II:В:С:D:Е).
Характерно, что доля ассоциированных комплексов чрезвычайно высока у ХМ и ЛПОНП и очень низка у ЛПВП, т.е. способность к образованию комплексов уменьшается с увеличением плотность ЛП.
Следует остановиться еще на одном подходе в разделении ЛП, учитывающем преобладание в них того или иного белка или липида. Согласно этому подходу, выделяют апо А- и апо В-содержащие ЛП, а также ЛП, богатые ТГ, ХС, ФЛ.
К ЛП, богатым ТГ относятся ХМ и ЛПОНП, ЛП , богатые ХС – это ЛПНП и ЛП ,богатые ФЛ – ЛПВП.
Состав и физико-химические свойства ЛП плазмы крови человека, богатых ТГ или ХС.[Климов, 1999]
| Показатели | ХМ | ЛПОНП | ЛПНП1 | ЛПНП2 |
| Средняя гидратированная плотность частиц, г / мл | 0,93 | 0,97 | 1,012 | 1,035 |
| Границы солевой плотности для выделения , г / мл | 1,006 | 1,006 | 1,006 – 1,019 | 1,019 – 1,063 |
| Диаметр частицы , нм | >100 | 25 - 75 | 22 - 24 | 19 – 23 |
| ММ * 10-6, Да | 500 | 5 - 13 | 3,9 – 4,8 | 2,7 – 4,0 |
| Скорость флотации, Sf | > 400 | 20 - 400 | 12 - 20 | 0 - 12 |
| Средний поверхностный потенциал, мВ | 0 | -7 | -7 | -7 |
| Подвижность в электрическом поле | остаются на старте | пре - b | b | b |
| Химический состав ЛП, % ТГ Белки ХС общий % ЭХС ФЛ |
80 – 95 1 – 2 0,5 – 3 46 3 - 9 |
50 – 70 5 – 12 15 – 17 57 13 - 20 |
24 – 34 14 – 18 35 – 45 66 11 - 17 |
5 – 10 20 – 25 45 – 48 70 20 - 30 |
| Основные апопротеины | В-48,С,Е,А | В-100,С,Е | В-100,С | В-100 |
| Содержание в плазме крови взрослых лиц натощак, мг/дл | след | 50 - 200 | 10 - 50 | 200 – 300 |
| Что переносят | ТГ пищи | Эндоген- ные ТГ | ЭХС, ТГ | ХС, ЭХС |
Состав и физико-химические свойства ЛП плазмы крови человека, богатых ФЛ [Климов, 1999].
| Показатели | Общая фрак- ция ЛПВП | ЛПВП2 | ЛПВП3 | ЛПОВП |
| Средняя гидратированная плотность частиц, г / мл | 1,130 | 1,090 | 1,150 | 1,230 |
| Границы солевой плотности для выделения, г / мл | 1,063 – 1,25 | 1,08-1,125 | 1,125-1,21 | 1,21-1,25 |
| Диаметр частицы, нм | 6 - 12 | 7 - 12 | 6 - 7 | 7 |
| ММ * 10-5, Да | 1,5 – 4,0 | 3,60 – 3,86 | 1,48 – 1,86 | 1,5 |
| Скорость флотации (Sf) | 0 - 9 | 3,5 – 9,0 | 0 – 3,5 | ¾ |
| Химический состав ЛП, % Белки ХС общий % ЭХС ФЛ ТГ | 45– 55 20– 27 78 2 – 40 3 - 5 | 33 – 41 18 – 28 74 30 – 42 4 - 8 | 45 – 59 12 – 25 81 23 – 30 2 - 6 | 62 3 90 28 5 |
| Основные апопротеины | А-I, А-II | А-I, А-II | А-I, А-II | ? |
| Содержание в плазме крови взрослых лиц натощак, мг/дл мужчины / женщины | 170 –350 220 - 470 | 50 – 120 70 - 200 | 120 –230 150 -270 | ~ 20 ~ 20 |
| Что переносят | ХС,ЭХС ФЛ | ЭХС, ФЛ |
Рис.2.
Роль ЛП.
ЛП плазмы крови являются уникальной транспортной формой липидов в организме человека и животных. Они осуществляют транспорт липидов как экзогеного (пищевого) происхождения, так и заново синтезируемых в печени и стенке тонкой кишки (т.е. эндогенного происхождения) в систему циркуляции и далее к местам утилизации или депонирования . Уже одного этого было достаточно, чтобы представить важную роль ЛП в жизнедеятельности организма. Вместе с тем нам известно теперь, что отдельные ЛП осуществляют “захват” избыточного ХС из клеток переферических тканей и его “обратный” транспорт в печень для окисления в желчные кислоты и выведение с желчью . Наконец, ЛП осуществляют транспорт жирорастворимых витамиов, гормонов и других биологически активных веществ. Среди них следует отметить соединения, в отношении липидов антиоксидантной активностью: a- ,g- токоферолы, a - и b - каротины, убихинон и т.д. Основными липидами , транспортируемыми в токе крови в составе липопротеидных комплексов, являются ТГ, НЭХС, ЭХС, ФЛ и небольшое количество НЭЖК. Основная масса НЭЖК транспортируется альбуминами крови [5,1999].
Наследственная недостаточность ЛП.
Существуют 3 редких вида наследственной недостаточности ЛП.
Абеталипопротеинемия. При абетолипопротеинемии имеется дефект синтеза апо-В, в плазме отсутствуют ХМ, ЛПОНП, ЛПНП. Клинически оно проявляется мальабсорбцией жиров, акантоцитозом, пигментным ретинитом и атаксической невропатией.
Гипобеталипопротеинемия. При этом состоянии наблюдается частичная недостаточность апо-В; ХМ, ЛПОНП и ЛПНП присутствуют, но в низких концентрациях.
Болезнь Танжье. При этой патологии снижена концентрация ЛПВП. Клинически это состояние характеризуется гиперпластическим, оранжевыми миндалинами и аккумуляцией эфиров ХС в других ретикулоэндотелиальных тканях. Патология связана с ускоренным катаболизмом апо А-I [8, 2000].
Переваривание и всасывание липидов.
Желчь.
Значение.
На заре формирования современного учения о внешнесекреторной функции печени, когда естествоиспытатели располагали лишь первыми научными факторами о количестве и качестве отделяемой на пищу желчи и о тех сдвигах, которые возникают в секреции желчи в связи с воздействиями на организм различных внешних и внутренних факторов, И. П. Павлов так оценил значение желчи: “. . . главная роль желчи – сменять желудочное пищеварение на кишечное, уничтожая действие пепсина как опасного для ферментов поджелудочного сока агента и черезвычайно благоприятствуя ферментам поджелудочного сока, в особенности жировому “.
С тех пор прошло много десятков лет и за истекшее время физиология, биохимия и клиника, широко используя новейшие физиологические, биохимические, физические и клинические приемы исследования, обогатились огромным количеством фактов, которые расширили наши знания относительно роли и значения желчи в организме.
Теперь мы следующим образом можем оценить значение желчи: она 1) сменяет желудочное пищеварение на кишечное путем ограничения действия пепсина и создания наиболее благоприятных условий для активности ферментов поджелудочного сока, особенно липазы ; 2) благодаря наличию желчных кислот эмульгирует жиры и , снижая поверхностное натяжение капелек жира, способствует увеличению его контакта с липолитическими ферментами; кроме того, обеспечивает лучшее всасывание в кишечнике нерастворимых в воде высших жирных кислот, холестерина, витаминов Д, Е, К и каротина, а также аминокислот; 3) стимулирует моторную деятельность кишечника, в том числе и деятельность кишечных ворсинок, в результате чего повышается скорость абсорбции веществ в кишечнике; 4) является одним из стимуляторов секреции поджелудочной железы, желудочной слизи, а самое главное – желчеобразовательной функции печени; 5) благодаря содержанию протеолитического, амилолитического и гликолитического ферментов участвует в процессах кишечного пищеварения; 6) оказывает бактериостатическое действие на кишечную флору, предупреждая развитие гнилостных процессов. Помимо перечисленных функций, желчь играет весьма активную роль в межуточном обмене веществ, например углеводном, жировом, витаминном, пигментном, порфириновом , особенно белка и содержащегося в нем фосфора, а также в регуляции водного и электролитного обмена, не говоря уже об ее обезвреживающей функции, функции кроветворения и функции свертывания крови. При голодании выделяющаяся желчь содержит до 600 – 800 мг белка, который, попадая в кишечник, подвергается переработке, после чего продукты его, главным образом аминокислоты, всасываются, поступают в кровь и используются клетками как пластический и энергетический материал. Тоже самое можно сказать и в отношении фосфора. Его количество доходит в печеночной желчи до 100мг % и в пузырной желчи до 200 мг % , а значительная часть фосфорных соединений, выделяемая с желчью в кишечник, вновь всасывается и по воротной системе поступает обратно в печень, осуществляя таким образом, гепато – энтеро – гепатический кругооборот. С желчью выделяются азотистые вещества, которые вновь всасываются и утилизируются организмом.
Кроме того , значение желчи определяется еще и экскреторной функцией, выведением из крови таких продуктов обмена, как серотонин, а также многих экзогенных веществ ( лекарственные вещества, соединения брома, йода, мышьяка, фенолфталеина и салициловой кислоты, соли тяжелых металлов и некоторые другие химические компоненты ).
Еще одним свойством обладает желчь: она раздражает чувствительные нервные окончания сосудов и мозговые центры и изменяет возбудимость нервно – мышечной системы.
Последствия нарушения секреции.
Значение желчи очень хорошо определяется и теми серьезными функциональными и даже структурными изменениями, которые возникают в организме при ее хронической потери. Впервые об этом стало известно из работ павловской лаборатории, когда у собак с желчными фистулами, систематически терявших желчь, на вскрытии оказалось размягчение костей ребер, позвоночника, таза и плечевого пояса. У больных людей , хронически теряющих желчь через послеоперационные свищи, также отмечены нарушения кальциевого обмена , изменения кислотно – щелочного равновесия крови и развитие геморрагического диатеза.
В клинике у больных с послеоперационными свищами наблюдалось закономерное снижение содержания альбумина в крови и уменьшение альбумино – глобулинового коэффициента, повышение гипергликемического и постгликемического коэффициентов. При этом возникали нарушения внешнесекреторной функции печени, появлялось анемия, кровоточивость, нарушение деятельности почек, нервной системы; появлялись симптомы расстройств трофических процессов.
Установлено, что при хронической потери желчи у собак в организме возникают нарушения обмена веществ, в частности кальциевого, липоидного, витаминного; тормозится синтез фибриногена в печени; развивается гипохромная гипорегенеративная анемия; в костном мозгу – нормобластический тип эритропоэза с умеренным нарастанием полихроматофильных нормобластов; в крови – снижается у ровень холестерина ( с 240 до 57 мг % ); в кишечнике нарушается всасывание питательных веществ; слизистая оболочка желудка и кишечника атрофируется; расстраивается нервно – гормональная регуляция функций внутренних органов, возникают трофические язвы и гистоморфологические изменения в яичниках, семенниках и передней доли гипофиза; в печени развивается цирроз, иногда некроз отдельных печеночных долек.
Но не только хроническая потеря желчи оказывает серьезное влияние на высшие мозговые центры. Изменения высшей нервной деятельности у животных возникают и при продолжительной задержке эвакуации нормальной желчи в кишку, когда значительно повышается концентрация биллирубина и желчных кислот в крови. При этом возникают изменения и в деятельности внутренних органов.
Проникая в ток крови, например при механической желтухе, желчь в начальной стадии немного снижает возбудимость коры головного мозга и вследствие этого уменьшаются пищевые условные рефлексы, а в последующей – значительно повышает возбудимость корковых клеток, что выражается в повышении уровня пищевых условных рефлексов. По мере накопления компонентов желчи в крови и тканях организма возрастает и степень угнетения высшей нервной деятельности и ряд сомато – вегетативных нарушений.
Таким образом , мы видим , как велико значение желчи, причем не только для обеспечения процессов пищеварения в желудочно – кишечном тракте, но и для нормальной работы клеток всего организма в целом, включая и клетки периферической и центральной нервной системы.[2,1980]
Химический состав желчи.
Желчь содержит несколько соединений, не встречающихся в других пищеварительных секретах: холестерин, желчные кислоты и желчные пигменты. Вещества в печеночной желчи можно разделить на два класса: 1) вещества, концентрации которых мало отличаются от их концентрации в плазме; 2) вещества, концентрации которых во много раз выше, чем в плазме. К первому классу относятся Nа+, К+, CI-, креатинин и холестерин; это свидетельствует о том, что полигональные клетки печени образуют безбелковый ультрафильтрат плазмы. Однако холестерин синтезируется в печени. К числу веществ второго класса относятся билирубин, а также вводимые в организм лекарственные препараты, которые выделяются с желчью, например бромсульфалеин, n-аминогипуровая кислота и пенициллин. Содержание желчных кислот в печеночной желчи составляет 2 – 5 мэкв/л.
У взрослого человека емкость желчного пузыря составляет 50 – 60 мл; он не только служит для хранения желчи, но и концентрирует ее путем абсорбции воды и электролитов, а также секретирует муцины. В результате получается раствор , содержащий лишь небольшие количества CI- и НСО3- ; он может быть нейтральным или слабокислым, достигая рН 5,6. В процессе реабсорбции [К+] слегка повышается, и конечная величина [Са2+] может составлять 15 – 30 мг/100мл. Объем желчи, выделяемой за день в норме, точно не известен, но через фистулу желчного пузыря можно собрать от 500 до 1000 мл за сутки.
Желчные кислоты, синтезируемые в печени, определяют главный вклад желчи в процесс пищеварения; эти кислоты находятся в желчи в виде желчных солей. В желчи, отобранной через фистулу ,концентрация желчных солей может варьировать от 0,5 до 1,5%. Два основных компонента ,гликохолевая (холилглицин) и таурохолевая (холилтаурин) кислоты, находятся в желчи человека в соотношении около 3:1. ежедневная секреция этих веществ составляет от 5 до 15 г.
Желчные пигменты образуются при деградации порфиринов в клетках ретикулоэндотелиальной системы, главным образом в печени. Присутствие билирубина придает свежей печеночной желчи золотисто-желтый цвет. Пузырная желчь может быть зеленой из-за окисления билирубина в биливердин. При стоянии любая желчь постепенно темнеет, изменяет цвет от золотистого к зеленому, синему и затем коричневому по мере окисления пигментов. Общее количество этих пигментов, выделяемое человеком за день , варьирует от 0,5 до 2,1 г. иногда желчь содержит небольшин количества копропорфирина, образующегося из гема.
Желчь содержит три липидных компонента , имеющих ограниченную растворимость, - желчные соли, фосфотидилхолин и холестерин. Пузырная желчь представляет собой эмульсию, в которой находятся смешанные мицеллы этих компонентов; растворимость ХС решающим образом зависит от концентрации желчных кислот и фосфотидилхолина. Основной составной частью желчи является НЭХС, впервые выделенный из желчных камней; его концентрация в пузырной желчи может достигать 1%. Пузырная желчь содержит также жирные кислоты, присутствующие в виде мыл в количествах, варьирующих от 0,5 до 1,2%, а также ТГ (0,5%) и фосфоглицериды(0,2%). Присутствие желчных солей, мыл и гликопротеидов обеспечивает стабилизацию пересыщенного раствора холестерина.
Гуморальная регуляция секреции.
На основании главным образом эксперементальных исследований в настоящее время стало очевидным, что буквально все железы внутренней секреции принимают участие в регуляции внешнесекреторной функции печени. Об этом свидетельствуют результаты опытов с введением животным таких гормональных веществ, как адреналин, инсулин, тироксин, питуитрин, эпинефрин, тиреоидин, гидрокортизон, адренокортикотропный гормон, а также опыты с частичной или полной экстирпацией гипофиза, щитовидной железы, ококлощитовидных желез, половых желез и надпочечников.
Инсулин, введенный вместе с пищей, например с яичным желтком, увеличивает количество выделяемой желчи по сравнению с тем, что бывает на прием одних желтков; причем такая повышенная реакция на пищу наблюдается и на второй, и на третий день после введения гормона, когда животные получают только яичные желтки. Эти наблюдения совпадают с тем, что отметил в своих опытах И. Х. Пасечник. В его опытах на собаках инсулин увеличивал на 50 – 67 % общее количество спонтанно выделяемой желчи и снижал уровень содержания в ней холатов и биллирубина.
Г. А. Петровский установил тормозящее влияние глюкозы крови на желчную секрецию; им же показано, что инсулин не только возбуждает печеночные клетки, но и снимает тормозной эффект, вызываемый глюкозой. Такой же эффект последствия М. А. Сукалло получила и в опытах с введением гормона кортизона.
Существенное влияние на внешнесекреторную функцию печени оказывает гормон щитовидной железы – тироксин, который при введении в кровь тормозит секрецию желчи. Тиреоидектомия или подавление функции щитовидной железы , наоборот, усиливает желчеобразовательную функцию печени. При экспериментальном тиреотоксикозе уменьшается, а при экспериментальном гипотериозе, наоборот, увеличивается количество секретируемой желчи.
Дискинетические явления в желчевыделительной системе и расстройства нормальной секреции желчи имеют место и при введении препаратов, содержащих гормон околощитовидных желез, или при удалении последних. Антидиуретический гормон в дозах 10 – 100 миллиединиц стимулирует желчеобразование, а удаление половых желез ведет к угнетению процесса желчеобразования с уменьшением спонтанной секреции желчи и снижением содержания в ней органических веществ. Но вместе с тем, количество выделяемой желчи на еду у кастрированных собак увеличивается на 15 – 29 % , латентный период желчевыделения укорачивается, удлиняется время желчевыделения, содержание билирубина в желчи повышается, и титрующаяся щелочность увеличивается.
Из гормональных веществ, образующихся в организме и оказывающих влияние на внешнесекреторную функцию печени, наиболее важное значение имеют собственные гормоны пищеварительной системы. К их числу относится прежде всего гормон холицистокинин. При ведении этого гормона человеку возникают сокращение желчного пузыря, расслабление сфинктера Одди и происходит выход желчи в кишку, этим он отличается от гормона секретина, который обладает способностью стимулировать желчеобразовательную функцию печени [2, 1980]. Секретин увеличивает объем желчи и содержание в ней бикарбонатов и хлоридов, но не влияет на секрецию желчных кислот.
Имеются некоторые данные о стимулирующем действии на желчевыделительную систему и гормона панкреозимина, если он вводится в больших дозах. Глюкогон также усиливает желчеотделение, но в отличии от секретина стимулирует образование желчи ,богатой хлоридами, и не вызывает значительных изменений в концентрации бикарбонатов. Холицистокинин ,церулеин, гастрин – II также стимулируют желчеотделение и увеличивают концентрацию бикарбонатов и хлоридов, однако холеретический эффект у них выражен слабее, чем у секретина. Есть сообщения об участии вазоактивного интестинального пептида (VYP) в регуляции процессов желчеобразования. VYP вызывает расширение сосудов печени и поджелудочной железы и стимулирует выделение жидкости, богатой бикарбонатами. По влиянию на секрецию желчи он является антогонистом секретина [4, 1986].
Наряду с гормонами в механизме желчевыделения могут играть роль и всосавшиеся в кровь продукты переваривания пищи. Однако приведенные в литературе по этому вопросу данные довольно противоречивы.
Таким образом многочисленные эксперементы на животных и наблюдения на человеке показали, что в механизме желчеобразования и желчевыведения существует нервно – гуморальная фаза , но обуславливается ли она только действием специфических гормонов, или действием продуктов переваривания пищи, или суммарным действием и тех и других, окончательно сказать не возможно [2, 1980].
ПАВ желудочно – кишечного тракта и механизмы
эмульгирования, значение.
Все ферменты, принимающие участие в гидролизе пищевых липидов растворены в водной фазе содержимого тонкого кишечника и могут действовать на молекулы липидов лишь на границе раздела липид/вода. Отсюда , для эффективного переваривания липидов необходимо увеличение этой поверхности с тем, чтобы большее количество молекул ферментов участвовало в катализе. Увеличение площади поверхности раздела достигается за счет эмульгирования пищевых липидов – разделение крупных липидных капель пищевого комка на мелкие. Для эмульгировани необходимы поверхностно-активные вещества – ПАВы , представляющие собой амфифильные соединения , одна часть молекулы которых гидрофобна и способна взаимодействовать с гидрофобными молекулами поверхности липидных капель, а вторая часть молекулы ПАВов должна быть гидрофильной, способной взаимодействовать с водой. При взаимодействии липидных капель с ПАВами снижается величина поверхностного натяжения на границе раздела липид/вода и крупные липидные капли распадаются на более мелкие с образованием эмульсии. В качестве ПАВов в тонком кишечнике выступают соли ЖК и продукты неполного гидролиза триацилглицеринов или ФЛ, однако основную роль в этом процессе играют желчные кислоты [6, 1999].
Они поступают в двенацатиперстную кишку с желчью в виде коньюгатов с глицином или таурином (гликохолевая, таурохолевая, гликохенодезоксихолевая, таурохенодезоксихолевая кислоты). У человека отношение глициновых коньюгатов к тауриновым составляет примерно 3:1.
В двенацатиперстную кишку вместе с пищевой массой заносится некоторое количество желудочного сока, содержащего соляную кислоту, которая в двенацатиперстной кишке нейтрализуется в основном бикарбонатами, содержащимися в панкреатическом соке и желчи. Образующиеся при разложении бикарбонатов пузырьки углекислого газа разрыхляют пищевую кашицу и способствуют более полному перемешиванию ее с пищеварительными соками. Одновременно начинается эмульгирование жира (ТГ). Соли желчных кислот адсорбируются в присутствии небольших количеств свободных ЖК и МГ на поверхности капелек жира в виде тончайшей пленки, препятствующей слиянию этих капелек. Кроме того , соли желчных кислот способствуют расчленению больших капелек жира на меньшие. Создаются условия для образовании тонкой и устойчивой жировой эмульсии с размером частиц 0,5 мкм и меньше. В результате эмульгирования резко увеличивается поверхность жиров, что облегчает взаимодействие их с липазой, т.е. ускоряет ферментативный гидролиз [5, 1999].
Расщепление липидов.Триглицериды.
В организм взрослого человека с пищей ежесуточно поступает 60 – 80 г жиров (ТГ) животного и растительного происхождения. Из этого количества подавляющая часть (более 85%) подвергается расщеплению в желудочно-кишечном тракте.
В полости рта ТГ не подвергаются никаким изменениям, так как слюна не содержит расщепляющих их ферментов. С желудочным соком выделяется липаза, получившая название желудочной, однако роль ее в гидролизе пищевых ТГ у взрослых людей не велика. Во–первых, в желудочном соке взрослого человека и других млекопитающих содержание липазы крайне низкое. Во-вторых, рН желудочного сока от оптимума действия этого фермента (оптимальное значение рН для желудочной липазы находится в пределах 5,5 – 7,5). В-третьих , в желудке отсутствуют условия для эмульгирования ТГ, а липаза может активно действовать только на ТГ, находящихся в форме эмульсии. Поэтому у взрослых людей неэмульгированные ТГ, составляющие основную массу пищевого жира, проходят через желудок без особых изменений.
Вместе с тем расщепление ТГ в полости желудка играет важную роль в пищеварении у детей, особенно грудного возраста. Умеренная кислотность желудочного сока у них ( рН около 5) способствует перевариванию эмульгированных ТГ молока желудочной липазой. Кроме того, при употреблении молока в качестве основного продукта питания возможно адаптивное усиление синтеза желудочной липазы.
Исследованиями М.Hmosh и соавт. Было показано, что слизистая оболочка корня языка и примыкающей к нему области глотки грудного ребенка секретирует свою собственную липазу в ответ на сосательные и глотательные движения при кормлении грудью. Эта липаза получила название лингвальной. Активность лингвальной липазы не успевает проявиться в полости рта, и основным местом ее действия является желудок. Оптимум рН лингвальной липазы лежит в пределах 4 – 4,5; он близок к величине рН желудочного сока у грудных детей. Лингвальная липаза наиболее активно действует на ТГ, содержащие ЖК с короткой и средней длиной цепи, что характерно для ТГ молока. Можно сказать, что жир молока – самый подходящий субстрат для этого энзима. Лингвальная липаза преимущественно расщепляет эфирную связь в sn – 3 положении ТГ,
в результате чего образуются 1,2(a, b)- ДГ и ЖК. ЖК с короткой цепью всасываются непосредственно в желудке, а длинноцепочечные вместе с ДГ поступают в тонкую кишку. Активность лиигвальной липазы у взрослых крайне низкая.
Очень близок по своим свойствам к лингвальной липазе энзим, секретируемый слизистой оболочкой гортани у новорожденных и получивший название преджелудочной липазы. Функция его та же , что и лингвальной липазы. Все три липазы (лингвальная, преджелудочная и желудочная), гидролизуя сложноэфирную связь преимущественно в sn-3 положении триглицерида, действуют на ФЛ и ЭХС.
Несмотря на то , что расщепление ТГ в желудке взрослого человека невелико, оно в какой-то степени облегчает последующее переваривание их в кишечнике. Даже незначительное по объему расщепление ТГ в желудке приводит к появлению свободных ЖК, которые, не подвергаясь всасыванию в желудке, поступают в кишечник и способствуют там эмульгированию жиров, облегчая, таким образом, воздействия на них липазы панкреатического сока. Кроме того, появление длинноцепочечных ЖК в двенацатиперстной кишке стимулирует секрецию понкреатической липазы.
Основная масса пищевых ТГ подвергается расщеплению в верхних отделах тонкой кишки при действии липазы панкреатического сока. Панкреатическая липаза является гликопротеидом, имеющим ММ 48 кДа ( у человека) и оптимум рН 8 – 9 . Она расщепляет ТГ, находящиеся в эмульгированном состоянии (действие ее на растворенные субстраты значительно слабее).Фермент катализирует гидролиз эфирных связей в a-,a1- положениях, в результате чего образуется b-МГ и освобождаются две частицы ЖК.Это отличает панкреатическую липазу от лингвальной, преджелудочной и желудочной липаз, при действии которых освобождается только одна ЖК.
Панкреатическая липаза , как и другие пищеварительные ферменты (пепсин, трипсин и химотрипсин), поступает в верхний отдел тонкой кишки в виде неактивной пролипазы. Превращение пролипазы в активную липазу происходит при участии желчных кислот и еще одного белка панкреатического сока – колипазы. Колипаза секретируется в виде проформы – проколипазы , и для ее превращения в активную колипазу требуется гидролиз специфических пептидных связей, который происходит при действии трипсина поджелудочного сока. Образовавшаяся активная колипаза образует с липазой комплекс в молярном отношении 1:1 за счет формирования двух ионных связей Lys – Glu и Asp – Arg. Образование такого комплекса приводит к тому, что липаза становится активной и устойчивой к действию трипсина.
Итак, основные продукты расщепления ТГ при действии панкреатической липазы - b-МГ и ЖК. На скорость катализируемого липазой гидролиза ТГ не оказывают существенного влияния ни степень ненасыщенности ЖК, ни длина ее цепи (С12 – С18).
Во время триптического гидролиза проколипазы освобождается пентапептид, названный энтеростатином, функция которого еще до конца невыяснена, но установлено, что, всасываясь в кровь, он угнетает аппетит:
Val – Pro – Asp – Pro – Arg
энтеростатин
Другими словами, энтеростатин можно рассматривать как своеобразный “кишечный гормон”, вызывающий чувство сытости при приеме и переваривании жирной пищи.
В панкреатическом соке , наряду с липазой, содержится моноглицеридная изомераза – фермент, катализирущий внутримолекулярный перенос ацила из b-положении МГ превращается в a-положение. В процессе переваривания пищевых жиров при участии этого фермента примерно 1/3 b-МГ превращается в a-МГ. Поскольку эфирная связь в a-положении глицерида чувствительна к действию панкреатической липазы, последняя расщепляет большую часть a-МГ до конечных продуктов – глицерина и ЖК (рис. 3). Меньшая часть a-МГ успевает всосаться в стенку тонкой кишки, минуя воздействие со стороны липазы.
Фосфолипиды.
Подавляющая часть ФЛ содержимого тонкой кишки приходится на фосфотидилхолин (лецитин), основная масса которого поступает в кишечник с желчью (11 – 12 г/сут) и меньшая (1 –2 г/сут) – с пищей.
Столь значительная разница в количествах экзогенных и эндогенных ФЛ, находящихся в тонкой кишке, послужила основанием для высказывания двух точек зрения относительно дальнейшей их судьбы. Согласно одной из них, и те, и другие ФЛ подвергаются в кишечнике атаке со стороны фосфолипазы А2, катализирующей гидролиз сложноэфирной связи в b-положении ФЛ. Фосфолипаза А2 выделяется в кишечник с панкреатическим соком в виде неактивного проэнзима и при воздействии на нее трипсина и ионов кальция превращается в активный энзим. В результате катализируемой фосфолипазой А2 реакции глицеро-ФЛ расщепляются с образованием лизофосфолипида (лизо-ФЛ) и ЖК. Для протекании этой реакции требуются соли желчных кислот. Таким образом, согласно приведенной точке зрения, судьба экзогенных и эндогенных ФЛ одна и та же. Авторы другой точке зрения считают, что ФЛ “желчного”(более того ,печеночного) происхождения, в отличии от пищевых ФЛ, не подвергаются воздействию фосфолипазы А2. При этом подчеркивается, что функция “желчных” ФЛ исключительно связана с гепатоэнтеральной циркуляцией желчи: с желчью они поступают в кишечник, с желчными кислотами участвуют в мицеллярной солюбилизации липидов и вместе с ним же возвращаются в печень. Таким образом, существуют как бы два пула ФЛ в кишечнике – “желчный” , защищенный от действия фосфолипазы, и “пищевой”, подверженный ее действию. Пока мы не можем объяснить причину существования двух пулов ФЛ и их различное отношение к действию фосфолипазы.
Образующиеся при действии фосфолипазы А2 лизо-ФЛ является хорошим ПАВом, и поэтому он способствует эмульгированию пищевых жиров и образованию смешанных жировых мицелл. Вместе с тем какая-то часть лизо-ФЛ может подвергаться расщеплению при действии другого фермента панкреатического сока – лизофосфолипазы, катализирующей гидролиз сложноэфирной связи в a-положении. В результате из лизолецитина освобождается последняя частица ЖК и образуется глицерилфосфохолин, который хорошо растворяется в водной среде и всасывается из кишечника в кровь.
СН2–О–СО–R1 СН2–О–СО–R1
½ Н2О R2СООН ½ Н2О R1СООН
СН–О–СО–R2 ¾¾¾¾¾¾¾® СН–ОН ¾¾¾¾¾¾¾®½ фосфолипаза А2 ½ ФОСФОЛИПАЗА А1
СН2–О–Р–О–СН2СН2N(CН3)3 СН2–О–Р–СН2СН2N(СН3)3
фосфотидилхолин лизофосфотидилхолин
СН2 – ОН
½
¾¾¾¾® СН – ОН
½
СН2 – О – Р – СН2СН2N(СН3)3
глицерилфосфохолин
Из других представителей ФЛ сфингомиелин всасывается в тонкой кишке в виде интактных молекул.
Холестерин.
В зависимости от рода пищи в организм взрослого человека вводится ежедневно 300 – 500 мг ХС ,содержащегося в пищевых продуктах частично в свободном (неэстерефицированном) виде, частично в виде эфиров с ЖК. Последние расщепляются на ХС и ЖК особым ферментом панкреатического сока – гидролазой ЭХС , или холестеринэстеразой. Активность фермента проявляется в присутствии желчных кислот [5,1999].
Химический состав и строение мицелл, механизм всасывания липидов.
В стенку кишечника легко всасываются вещества, хорошо растворимые в воде. Из продуктов расщепления липидов к ним относятся, например, глицерол, аминоспирты и жирные кислоты с короткими углеводородными радикалами (С8 - С10), натриевые или калиевые соли фосфорной кислоты. Эти соединения из клеток кишечника обычно поступают непосредственно в кровь и вместе с током крови транспортируются в печень.
В то же время большинство продуктов переваривания липидов:высшие жирные кислоты, моно- и диацилглицерины,холестерол, лизофосфолипиды и др. плохо растворимые в воде и для их всасывания в стенку кишечника требуется специальный механизм. Перечисленные соединения, наряду с желчными кислотами и ФЛ, образуют мицеллы. Каждая мицелла состоит из гидрофобного ядра и внешнего мономолекулярного слоя амфифильных соединений, расположенных таким образом, что гидрофильные части их молекул контактирует с водой, а гидрофобные участки ориентированны внутрь мицеллы, где они контактируют с гидрофобным ядром. В состав мономолекулярной амфифильной оболочки мицеллы входят преимущественно ФЛ и желчные кислоты, сюда же могут быть включены молекулы холестерола. Гидрофобное ядро мицеллы состоит преимущественно из высших жирных кислот, продуктов неполного расщепления жиров, эфиров холестерола, жирорастворимых витаминов и др [6, 1999].
Основная часть ТГ всасывается после расщепления их липазой на ЖК и b-МГ. Всасывание этих соединений происходит при участии желчи, которая содержит соли желчных кислот, ФЛ и НЭХС. Образовавшиеся при действии липазы ЖК и b-МГ формируют с этими компонентами желчи смешанные мицеллы.
Относительно механизма всасывания жировых мицелл или их ингредиентов нет единой точки зрения. Прежде всего признается возможность непосредственного проникновения мицелл в эпителиальные клетки ворсинок слизистой оболочки тонкой кишки. Согласно этой точки зрения, мицеллы проникают внутрь эпителиальных клеток целой частицей путем так называемой мицеллярной диффузии без затраты энергии. В клетках происходит распад мицеллярного комплекса, при этом желчные кислоты сразу же поступают в кровь и с током крови через воротную вену доставляются в печень, где снова переходят в состав желчи. Согласно другой точке зрения , проникновение жировых мицелл внутрь эпителиальных клеток частично или полностью осуществляется путем пиноцитоза. Наконец , допускается возможность перехода только липидных ингредиентов(молекулярная диффузия липидов) из жировых мицелл внутрь эпителиальных клеток при соприкосновении мицелл с поверхностью слизистой оболочки ворсинок кишечника. При этом соли желчных кислот не проникают внутрь клеток, а остаются в просвете кишечника и подвергаются обратному всасыванию , главным образом, в подвздошной кишке.
Каков бы ни был механизм всасывания липидов, исключительно важное значение при этом имеет гепатоэнтеральная циркуляция желчных кислот из печени в кишечник и обратно, обеспечивающая всасывание больших количеств ЖК и МГ (50 – 70, а иногда и более 100 г/сут) при относительно невысоком общем пуле желчных кислот (2,8 – 3,5 г). Последние в процессе гепатоэнтеральной циркуляции совершают 6 – 8 циклов, поступая, таким образом, в проксимальный отдел тонкой кишки в суммарном количестве около 18 г /сут при потере примерно 0,5 г . Эта потеря компенсируется образованием в печени такого же количества желчных кислот из ХС.
Продукты расщепления ТГ (ЖК и b-МГ) в процессе всасывания сначала поступают в мембрану эпителиальных клеток ворсинок тонкой кишки. Из внутренней поверхности мембраны они переносятся в цитоплазму к месту ресинтеза ТГ – в гладкий эндоплазматический ретикулум, причем ЖК транспортируются с помощью связывающего их белка.
ЖК со средней длиной углеводородной цепи (С6– С12), поступившие в эпителиальные клетки слизистой оболочки тонкой кишки, не участвуют в ресинтезе ТГ. Они непосредственно поступают в воротную вену, связываются альбуминами и транспортируются в печень. Такие ЖК оказались полезными в качестве пищевой добавки (взамен обычных пищевых жиров) больным с недостаточностью панкреатической липазы и липопротеидлипазы (ЛПЛ).
Как уже упоминалось, при переваривании жиров освобождаются некоторые количества глицерина (примерно 1/5 от его содержания в пищевых ТГ). Будучи хорошо растворимым в воде, глицерин легко всасывается в тонкой кишке и поступает в воротную вену.
ФЛ и лизо-ФЛ, которые участвуют в образовании смешанных жировых мицелл, всасываются в составе этих мицелл, не подвергаясь расщеплению. Основная часть их, как уже отмечалось, подвергается в полости тонкой кишки ферментативному гидролизу. При этом всасывание ЖК, образовавшихся при гидролизе ФЛ, лизо-ФЛ и ЭХС, происходит точно так же, как и всасывание ЖК, образовавшихся при расщеплении ТГ.
Отдельно следует остановится на вопросе о всасывании ХС, источником которого в тонкой кишке являются:
· ХС пищи(0,3-0,5 г/сут; у вегетарианцев значительно меньше);
· ХС желчи (ежедневно с желчью в тонкую кишку выделяется1-2 г эндогенного НЭХС);
· ХС, содержащийся в слущенном эпителии желудочно-кишечного тракта и в кишечных соках (до 0,5 г/сут).
В общей сложности в кишечник поступает 1,8-2,5 г эндогенного и экзогенного ХС. Из этого количества около 0,5 г ХС выделяется с фекалиями в виде востановленного продукта – капростерина и очень не большая часть в виде окисленных продуктов – холестеноно и др. И восстановление , и окисление ХС происходят в толстой кишке под воздействием ферментов микробной флоры. Основная часть ХС в неэстерефицированной форме подвергается всасыванию в тонкой кишке в составе смешанных жировых мицелл, состоящих, как уже отмечалось выше, из желчных кислот, ЖК, МГ, ФЛ и лизо-ФЛ.
Одной из причин уменьшенного всасывания жиров в тонкой кишке может быть не достаточно полное их расщепление вследствие либо пониженной секреции панкреатического сока (недостаток панкреатической липазы), либо недостаточного выделения желчи. Второй , наиболее частой причиной пониженного всасывания жира является нарушения функции кишечного эпителия, наблюдаемое при энтеритах, гиповитаминозах, недостаточности коры надпочечников и некоторых других патологических состояниях. В этом случае b-МГ и ЖК , образовавшиеся в полости кишечника, не могут нормально всасываться из-за повреждения эпителиального покрова кишечника.
Уменьшение всасывания ХС будет происходить, если с пищей употреблять большое количество растительных стеринов (главным образом, b-фитостерина), которые по конкурентному механизму препятствуют вхождению ХС в состав смешанных жировых мицелл, хотя сами по неизвестной причине не всасываются. На приеме фитостеринов видно, что малейшие изменения в структуре ХС ведут к существенному изменению физико-химических и физиологических свойств. В целом, при обычном смешанном питании и с учетом того, что принятый с пищей ХС образует в тонкой кишке общий пул с ХС, секретируемым желчью, всасыванию подвергается не более 60% пищевого ХС [5,1999].
Механизм ресинтеза липидов в энтероцитах, значение.
Более ста лет тому назад А. Перевозников в статье “К вопросу о синтезе жиров” сообщил, что он вводил через зонд в двенадцатиперстную кишку голодной собаки смесь мыла (соль ЖК) и глицерина и спустя 1,5ч на секционном материале наблюдал следующую картину: эпителий ворсинок тонкой кишки животного оказался наполненным “жировыми шариками” разной величины, а лимфа грудного протока выглядела как молоко. Автор сделал фундаментальный вывод, что в кишечной стенке собаки происходит синтез ТГ из ЖК и глицерина.
По современным представлениям ресинтез ТГ происходит в эпителиальных клетках (энтероцитах) слизистой оболочки ворсинок тонкой кишки двумя путями. Первый путь - b-моноглицеридный. Долгое время он считался единственным. Суть его состоит в том ,что b-МГ и ЖК, проникшие в процессе всасывания в эмителиальные клетки кишечной стенки, задерживаются в гладком эндоплазматическом ретикулуме клеток. Здесь из ЖК образуется их активная форма – ацил-КоА – и происходит ацилирование b-МГ с образованием сначала ДГ, а затем ТГ. Все реакции катализируются энзимнм комплексом – триглицерид-синтетазой, включающим в себя ацил-КоА-синтетазу, моноглицерид-ацилтрансферазу и диглицерид-ацилтрансферазу [5, 1999].
CH2–OH H2C–O–C Н2С–О–С
½ + R–CO-SKoA ½ + R–CO–SKoA ½
CH–O–C ¾¾¾¾¾¾¾® HC–O–C ¾¾¾¾¾¾® НС–О–С
½ - HC–KoA ½ - HC–KoA ½
CH2–OH H2C–OH Н2С–О-С
b-МГ ДГ ТГ [6,1999]
Второй путь ресинтеза ТГ - a-глицерофосфатный. Он протекает в шероховатом эндоплазматическом ретикулуме эпителиальных клеток и включает следующие реакции:
· Образование активной формы жирной кислоты – ацил-КоА – при участии ацил-КоА-синтетазы (тиокиназы);
· Образование a-глицерофосфата при участии глицеролкиназы;
· Превращение a-глицерофофсфата в фосфатидную кислоту при участии глицерофосфат-ацилтрансферазы;
· Превращение фосфатидной кислоты в ДГ при участии фосфатидат-фосфогидролазы;
· Ацилирование ДГ с образованием ТГ при участии ДГ-ацилтрансферазы [5, 1999].
Высшие жирные кислоты перед их включением в состав более сложных липидов , должны быть активированы. Процесс активации высших жирных кислот состоит из двухэтапов:
а) на первом этапе идет взаимодействие высших жирных кислот с АТФ с образованием ациладенилата:
R – COOH + АТФ ¾¾®R – CO ~ АМФ + Ф~Ф
Образующийся в ходе реакции пирофосфат расщепляется на два остатка фосфорной кислоты и реакция образования ациладенилата становится необратимой – термодинамический контроль направления процесса.
б) на втором этапе ациладенилат взаимодействует с HS-КоА с образованием ацил-КоА ( R – CO~SKоА):
R – CO ~ АМФ + HS – КоА ¾¾® R – CO ~ SkoA + АМФ
В ходе активации высшей жирной кислоты АТФ распадается до АМФ и двух остатков фосфорной кислоты, таким образом, активация жирной кислоты обходится клетке в два макроэргических эквивалента. Во всех своих превращениях в клетках жирные кислоты участвуют в активированной форме.
Далее идет активация глицерола при участии глицеролкиназы:
H2C – OH H2C – OH
½ ½
HC – OH + АТФ ¾¾® HC – OH + АДФ
½ ½
H2C – OH H2C – O – PO3H2
Затем при последовательном переносе двух ацильных остатков образуется фосфатидная кислота:
Н2С–ОН Н2С–О–СО–R H2C-О-CO–R
½ + R-CO–SКоА ½ + R–СО–SKoA ½
НС–ОН ¾¾¾¾¾® НС–ОН ¾¾¾¾¾¾® HC-О-CO–R
½ - HS–KoA ½ -HS-KoA ½
Н2С–О–РО3Н2 Н2С–О–РО3Н2 H2C-О-PO3H2
Далее от фосфатидной кислоты гидролетическим путем отщепляется остаток фосфорной кислоты с образованием ДГ:
H2C–O–CO–R H2C – O – CO – R
½ + H2O ½
HC–O–CO–R ¾¾¾® HC – O – CO – R
½ - H3PO4 ½
H2C–O–PO3H2 H2C – OH
К образовавшемуся ДГ присоединяется остаток высшей жирной кислоты:
H2C – O – CO – R H2C – O – CO – R
½ + R–CO–SKoA ½
HC – O – CO – R ¾¾¾¾¾¾® HC – O – CO – R
½ - HS – KoA ½
H2C – OH H2C – O – CO – R
В результате образуется ТГ [6, 1999].
Как видно , первая и последняя реакции a-глицерофосфатного пути ресинтеза ТГ повторяют аналогичные реакции b-глицерофосфатного пути. Протекание того или иного пути ресинтеза ТГ зависит от состава продуктов расщепления пищевых липидов, поступивших в кишечную стенку. a-Глицерофосфатный путь преобретает значение , когда в стенку поступили преимущественно одни ЖК. Если в стенку поступили ЖК вместе с b-МГ, тогда запускается b-моноглицеридный путь. Более того, наличие в эпителиальных клетках избытка b-МГ тормозит протекание a-глицерофосфатного пути.
Какая-то часть ТГ может образоваться в кишечной стенке целиком из эндогенных предшественников. C.Mansbach и S.Parthasarathy считают, что если ТГ кишечной стенки образуются из метаболитов пищевых жиров, то они идут на образование хиломикронов и быстро поступают в лимфу. Если же ТГ образуются из эндогенных метаболитов , то они в лимфу непоступают, а секретируются в просвет тонкой кишки. Этим авторы объясняют развитие стеатореи при некоторых болезненных состояниях у пациентов, находящихся на без жировой диете.
В энтероцитах , наряду с ресинтезом ТГ, происходит также и ресинтез ФЛ. В образовании фосфатидилхолинов и фосфатидилэтаноламинов участвует ресинтезированный a-,b-ДГ, а в образовании фосфатидилинозитов – ресинтезированная фосфатидная кислота. Участие этих субстратов в образовании ФЛ в стенке кишечника происходит по тем же закономерностям, что и в других тканях. В процессе всасывания в кишечную стенку поступает какое-то количество лизо-ФЛ, главным образом, лизофосфотидилхолина. Судьба последнего может быть двоякой: или он подвергается расщеплению с образованием сначала глицерилфосфохолина, а затем a-глицерофосфата, или же подвергается ацилированию с образованием фосфатидилхолина (лецитина) (схема 1) [5, 1999].
Фосфотидилхолин
Н2О ½
½фосфолипаза А2
R2СООН ¯
Лизофосфотидилхолин
Н2О ½
½лизофосфолипаза
R1СООН ¯(фосфолипаза В?)
Глицеринфосфохолин
Н2О ½
½глицерилфосфохолингидролаза
¯
a-глицерофосфат + холин
схема 1. Превращение фосфотидилхолина в
a-глицерофосфат.[Климов,1999]
Клетки кишечника способны ресинтезировать ФЛ и из поступающих в них при пищеварении свободных ЖК, глицерола и аминоспиртов. Этот процесс можно разбить на три этапа:
а) образование диацилглицерида, ранее рассмотреное;
б) активация аминоспирта: аминоспирт, например, этаноламин подвергается при участии этаноламинкиназы энергозависимому фосфолирированию :
NH2–CH2–CH2–OH + АТФ ¾¾® NH2–CH2–CH2–O–PO3H2 + АДФ
Затем при взаимодействии фосфорилированного аминоэтанола с ЦТФ идет образование активированной формы аминоспирта – ЦДФ-этаноламина:
NH–CH–СH–O–Ф + ЦТФ ¾® ЦДФ-этаноламин + пирофосфат
Реакция катализируется фосфоэтаноламинцитидилтрансферазой. Образовавшийся в ходе реакции пирофосфат расщепляется пирофосфатазой – термодинамический контроль направления процесса, с которым мы уже знакомились.
в) образование глицерофосфолипида:
ЦДФ-этаноламин + диглицерид ¾¾® фосфотидилэтаноламин + ЦМФ
Реакция катализируется фосфоэтаноламин-диацилглицеролтрансферазой.
С помощью подобного механизма может синтезироваться и фосфотидилхолин [6, 1999].
В кишечной стенке происходит также реэстерификация ХС. До недавнего времени считали, что эта реакция осуществляется при участии панкреатической холестерин-эстеразы (гидролазы) и что этот фермент в зависимости от условий может не только гидролизовать ЭХС, но и синтезировать их. В последние годы установлено, что образование ЭХС происходит в микросомах энтероцитов и что этот процесс катализируется другим ферментом – ацил-КоА-холестерин-ацилтрансферазой:
АХАТ
ХС + Ацил-КоА ¾¾¾® ЭХС + КоА
Эффективность эстерификации ХС в энтероцитах имеет большое значение для его всасывания. Предложены препараты, угнетающие активность указанного фермента и , следовательно, уменьшающие всасывание ХС.
Таким образом, продукты расщепления пищевых жиров, образовавшиеся в полости кишечника и поступившие в его стенку, снова используются для ресинтеза жиров. Биологический смысл этого процесса сводится к тому, что в стенке кишечника синтезируются жиры, более специфичные для данного вида животного и отличающиеся от пищевого жира. В известной степени это обеспечивается тем, что в синтезе ТГ и ФЛ и в эстерификации ХС в кишечной стенке принимают участие, наряду с экзогенными (пищевыми), и эндогенные ЖК, доставляемые в клетки следующими путями : а) синтезированные заново в самих клетках или “модифицированные”, например, путем удлинения цепи; б) доставленные в клетки из кровеносного русла; в) синтезированные в печени и попавшие сначала в кишечник в составе ФЛ желчи, а затем проникшие в эпителиальные клетки кишечника в составе жировых мицелл. Кроме того , в клетках кишечника происходит своеобразное перераспределение общего пула ЖК, например, ЖК эндогенного происхождения может быть использована для эстерификации ХС как экзогенного , так и эндогенного происхождения; ЖК, ранее входившая в состав ФЛ, может быть использована для ресинтеза ТГ , и наоборот.
В заключении следует подчеркнуть, что ресинтезированные и вновь синтезированные в стенке кишечника липиды не поступают в чистом виде в кровь, а используются для образования особых, богатых триглицеридами , липид-белковых комплексов – ХМ и в таком виде всасываются сначала в лимфу, а затем в кровь [5, 1999].
Образование и обмен хиломикронов, значение.
Хиломикроны (ХМ) известны с 1774 г., когда английский врач У. Хьюсон обнаружил белесоватый вид крови при кровопускании и установил, что причиной этого является абсорбционная липемия. В 1920 г. С. Кейдж локализовал ХМ под микроскопом после приема жирной пищи, как “танцующие в сыворотки частицы, диаметром в несколько раз меньше эритроцитов” и дал им современное название [3, 2000].
Основной функцией ХМ является транспорт экзогенных, поступающих с пищей ТГ, которые составляют до 90% липидных компонентов этих липопротеиновых частиц. Образование ХМ происходит в энтероцитах поверхностного слизистого слоя кишечника и зависит во многом от количества потребляемых жиров и характера содержащихся в них ЖК. Способность стенки кишечника синтезировать ХМ проявляется только при наличии высших ЖК с числом атомов углерода не менее 12. В таком случае главные продукты гидролиза плазмы – ЖК , 2-моноглицериды и диглецериды поступают путем диффузии (энергозависимый процесс) в энтероциты, где происходит ресинтез ТГ на гладком эндоплазматическом ретикулуме в апикальной части клеток. При наличии в пище ТГ с ацилами короткой длины ХМ не образуются , а ЖК после всасывания в кишечнике поступают в кровь воротной вены печени, не попадая в лимфатическую систему. Электронномикроскопические исследования позволяют выявить осмиофильные включения – предшественники ХМ – в цитоплазме энтероцитов и проследить их перемещение от гладкого эндоплазматического ретикула к аппарату Гольджи в супроядерную часть клетки. Возможно, аппарат Гольджи ответственен за присоединение углеводных компонентов к апапротеинам липопротеиновых частиц. Из аппарата Гольджи сформированные частицы ХМ перемещаются в составе везикул к плазмолемме , где посредством экзоцитоза покидают клетку и переходят в межклеточное пространство , а далее – в лимфу.
Секретируемые в лимфу ЛП-частицы претерпевают ряд превращений до их окончательного формирования в ХМ, наблюдаемые в токе крови. указанные изменения заключаются во взаимообмене отдельных апопротеинов, в первую очередь с ЛПВП. Показано , что при взаимодействии ХМ с ЛПВП2 с последних переходят на ХМ апо-ЛП С и Е , в то время как апо-ЛП А-IV покидают Хми участвуют в формировании в сосудистом русле ЛПВП. Природа таких перемещений заключается в большем сродстве апо-ЛП С к поверхностным слоям триглицерид-обогащенных липопротеиновых комплексов и в его более высокой поверхностной активности (рис. 4) [12,1990].
Ключевую роль в сборке частиц играет апопротеин В48. В них также широко представлены апопротеины С I – III и имеют апопротеины А I – II кишечного и печеночного происхождения. Однако, свежесекретированные частицы ХМ, практически, лишены апопротеинов С и А и приобретают их в результате контакта с челночными ЛПВП уже в плазме крови.
ХМ – это первый транспортер экзогенных пищевых липидов, прежде всего, ТГ, на пути через лимфу крови. Их метаболические превращения известны как экзогенный путь кругооборота ЛП.
С кровью ХМ переносятся, в первую очередь, в правое сердце и легкие, а затем в большой круг кровообращения. При этом они все время теряют значительные количества ТГ за счет липопротеид-липолиза и их гидрофобное ядро заметно “худеет”. ХМ превращаются в остаточные частицы , в которых ФЛ, ХС и апопротеины находятся в относительном избытке. На поверхности похудевшей частицы ХМ возникают складки избыточной оболочки, которые могут отрываться от частицы , замыкаться в особые малые богатые фосфолипидами и апопротеинами С, Е и А “насцентные диски” и служат затем основой для наполнения гидрофобными липидами и образования ЛПВП. Механизм эффекта , обеспечивающего просветление липемической плазмы, которое интенсивно идет уже в первые 15 минут и за 12 – 14 ч после приема жирной пищи полностью убирает из плазмы ХМ, обеспечивается ферментом липопротеиновой липазой (ЛПЛ). Этот энзим капиллярной стенки и освобождается в плазму в ответ на жировую нагрузку и гепарин. Наибольшую липолитическую активность проявляют капилляры жировой ткани , легких и сердца, кроме того ЛПЛ выделяется в печени , селезенке, почках, лактирующей молочной железе и диафрагме. Очевидно , что это связано с интенсивным отложением ТГ в адипоцитах, с секрецией липидной эмульсии в молоко и с энергетикой миокарда и диафрагмы, использующих в топливных целях много ЖК. Любопытно , что в легких процесс частичного метаболизма ХМ играет ключевую роль для обеспечения высокой активности альвеолярных макрофагов и существенно необходим для синтеза ФЛ сурфактанта. В связи с этим , при легочных инфекциях благотворно действует жировая диета. Еще народные знахари применяли барсучий и медвежий жир и собачье сало при чахотке. Такая процедура, как искусственный лечебный пневмоторакс , опосредует свой эффект не только через возникающую в спавшемся легком венозную гиперемию и усиление фибропластических процессов. Известное значение имеет и усиление недыхательных функций легких при снижении вентиляции. Традиционное питание северных народов , находящихся под воздействием климатических факторов повышенного риска бронхита и пневмонии , не случайно богато жирами. Эта особенность экологии повышает резистентность чукчей , эскимосов и других представителей малых реликтовых этносов к бронхолегочной патологии. К сожалению, чтобы осознать это, понадобился печальный опыт “окультуривания” советского и американо-канадского Севера , когда форсированный переход на европеизированную диету и образ жизни , не смотря на формально “улучшенные условия существования” , привел к значительному возрастанию патологической пораженности болезнями дыхательной системы у аборигенов. Конечно, здесь сыграло роль и учащение контактов с носителями новых для северян изолятов штаммов инфекционных возбудителей. Но, по крайней мере, во многих случаях для индивидов оказывается биологичеки выгодно оставаться в рамках привычной экологии.
Активность ЛПЛ стимулируется инсулином и СТГ. У человека более 80% липогенеза в адипоцитах идет на основе готовых ЖК, поставляемых ЛПЛ-реакцией и только 20% синтезируется из углеводных предшественников в самих жировых клетках.
Гепарин не является кофактором ЛПЛ, но запускает ее секрецию. Коэнзимную роль для ЛПЛ выполняет компонент ХМ, апопротеин С – II. Вместе с тем, апопротеин С – III , наоборот , ингибирует ЛПЛ. Таким образом, от соотношения С – II и С – III может зависеть скорость просветления постгепариновой плазмы. Альбумин ,подхватывая и удаляя из сферы действия реакции НЭЖК, также значительно ускоряет просветление липемической плазмы. В связи с этими фактами , липемия очень часто сопровождается тромбофилитическим состоянием. Более того, гипоальбуминемия , свойственная голоданию и нефротическому синдрому , протекает с задержкой катаболизма ХМ и других ЛП в плазме и гиперлипопротеинемиями. Кроме ХМ, ЛПЛ аналогичнодействует и на ЛПОНП. Остаточные частицы ХМ теряют апопротеины С и А, переходящие на частицы ЛПВП. Через обмен апопротеинов ЛПВП могут регулировать скорость катаболизма ХМ и ЛПОНП, так как служат челноком , снабжающим богатые триглицеридами ЛП активаторами ЛПЛ апопротеинами С. В конце концов, остатки ХМ приобретают из состава ЛПВП апопротеин Е, который способствует их захвату печенью через особый апо-Е-чувствительный и комбинированный , апо-В/Е-чувствительный рецепторы , и подвергаются рецепторному эндоцитозу в гепатоциты , где расщепляются. При этом ХС и другие липиды поставляются в печень, которая использует их для продукции желчных кислот и прямой экскреции липидов в желчь, для собственных пластических и энергетических нужд и для продукции ЛПОНП [3,2000].
В результате ферментативного воздействия ЛПЛ на частицу ХМ, направленного главным образом на ТГ ядра, происходит их гидролитическое расщепление до МГ и свободных ЖК. Последние диффундируют в клетки подлежащих тканей путем латеральной диффузии через мембраны и используются либо сразу же для энергообеспечения (функционирующие мышечные ткани, паренхиматозные органы) , либо запасаются в форме жировых включений (адипоциты жировой ткани). Учитывая , что при гликолизе ХМ расщепляется свыше 70% ТГ их сердцевины, содержащей в норме 90 – 95 % данного класса липидов, становятся очевидными наблюдаемые при этом резкие конформационные нарушения всей липидно-белковой гетеромолекулы в целом. Отмечается интенсивное высвобождение ФЛ, белков , НЭХС из поверхностных слоев ХМ.
ЛП-частицы , образовавшиеся из ХМ после их взаимодействия с ЛПЛ, получили специальное название – “ремнантные”.
Метаболическая судьба “ремнантных” частиц состоит в их утилизации печенью. Исследования последних лет показывают , что скорость поглощения “ремнантных” частиц печенью зависит от ихапопротенового состава. Апо-ЛП Е (в частности , Е-III И Е-IV изоформы) ускоряют данный процесс , в то время как апо-ЛП С ингибирубт, даже в присутствии адекватных количеств апо-ЛП Е [12, 1990].
Нарушение переваривания и всасывания липидов.
Стеаторея .
Во всех случаях нарушения переваривания и всасывания липидов появляется стеаторея. Это случается:
· Из-за отсутствия желчи в кишечнике (синдром ахолии);
· Из-за нарушения поступления в кишечник панкреатического сока. Отметим, что ухудшению эмульгирования и переваривания жиров способствует и низкая кислотность желудочного сока;
· Из-за первичной мальабсорбции (целиакия, тропическая спру, болезнь Уиппла, другие хронические энтериты, гиповитаминоз по фолиевой кислоте);
· При приеме значительных количеств тугоплавких липидов животного происхождения (например, бараньего жира), особенно у детей;
· При ускоренной перистальтике кишечника;
· При ингибирующем действии антибиотиков (неомицин, хлортетрациклин) и блокаторов фосфорилирования (монойодацетат, флоридзин) на функции энтероцитов;
· При избытке двухвалентных щелочноземельных катионов в пище и воде, что способствует образованию труднорастворимых кальций-магниевых солей ЖК. Задержка липидов, в частности, ХС в просвет кишечника достигается и с помощью ионообменных смол, связывающих желчные кислоты (холестирамин, квестрол, холестипол).
При стеаторее стул становится частым и липким, из-за ахолии – часто глинистым на вид, содержит липидные капли и беловатые комочки мыл (кальциевых и магниевых солей ЖК). При хронической стеаторее вторично развивается гиповитаминоз по жирорастворимым витаминам [3, 2000] и нехватка незаменимых ЖК [5, 1999]. Довольно закономерны коагулопатия и остеопороз.
Если стеаторея вызвана ахолией , то она сопровождается обесцвечиванием кала. Стул содержит не всосавшиеся , но полупереваренные (за счет действия мыл) липиды. При панкреатической стеаторее нет ахолии , кроме того, нарушено переваривание и всасывание и других , не липидных компонентов пищи. При нарушении переваривания и всасывания липидов возможно понижение содержания ХМ и других ЛП, а значит – ТГ и ХС – в лимфе и плазме крови.
Компенсаторным механизмом при нарушении абсорбции липидов в верхних отделах тонкой кишки является активизация их всасывания в нижних отделах [3, 2000].
Хиломикронемия.
Установлено, что при задержке катаболизма ХМ, из-за низкой активности ЛПЛ ,развивается продленная или стабильная хиломикронемия (гиперлипопротеинемия (ГЛП) ( I и V типа) .
Первичная гиперлипопротеинемия I типа или болезнь М. Бюргера – О. Грютца описана в 1932 г. и связана с наследственным аутосомно-рецесивным дефектом ЛПЛ. В крови накапливается значительное количество ХМ из-за блока их катаболизма. ХМ провоцируют тромбоз и ишемические микронекрозы, которые особенно характерны для поджелудочной железы. Характерны наблюдаемые с раннего детства абдоминальные колики. Формируется хронический рецидивирующий панкреатит. Бывают случаи молниеносных смертельных обострений. Большое значение имеет частичный гидролиз ХМ панкреатической липазой, который проходит в микроциркуляторном русле органа. Лизолицетин и ЖК в избытке оказывают на панкреатические клетки местное токсическое действие, связанное с детергентным эффектом и разрушением клеточных мембран.
На коже видны характерные стигмы заболевания – желтовато-розовые папулы на плечах, спине, ягодицах. Этот результат фагоцитоза ХМ гистоцитами дермы и образования эруптивных ксантом. Макрофаги тоже перегружаются ХМ ,что ведет к гепатоспленомегалии и появлению пенистых клеток в костном мозге. На бледном глазном дне видны белые сосуды, что известно как lipemia retinalis. В плазме, которая натощак остается мутной и дает сливкообразный слой при стоянии , повышен уровень ТГ , но ХС в норме. ГЛП I не реагирует на гепарин. Уровень апопротеина С II остается в норме. Так как ХМ не проникают через эндотелий, атеросклероз не ускоряется.
Приобретенная фенокопия ГЛП I формируется у больных с аутоимунными заболеваниями соеденительной ткани, особенно часто – при системной красной волчанке. Антитела против гликозаминогликанов при этих болезнях нарушают процесс гепариновой активации ЛПЛ.
Первичная гиперлипопротеинемия V типа развивается при аутосомно-рецессивном отсутствии апопротеина С II, важного кофактора ЛПЛ. В отличие от ГЛП I , в крови накапливаются оба главных субстрата ЛПЛ – ХМ и ЛПОНП. Основные симптомы болезни сходны с ГЛП I . Однако, панкреатит бывает менее тяжелым и клиника болезни развивается гораздо позже , во взрослом состоянии. Больные не имеют апо-С II. Гепарин не эффективен. При введении свежей донорской сыворотки здоровых лиц, изобилующей этим апопротеином, наступает быстрое , но временное облегчение. Эруптивные ксантомы менее выражены , чем при ГЛП I. Вторичная приобретенная ГЛП, соответствующая паттерну ГЛП V , бывает при гликогенозе Гирке, алкоголизме и использовании пероральных противозачаточных средств, если печень пациентов вырабатывает очень много ЛПОНП. Возможно , имеет место вторичное торможение активности ЛПЛ избытком ХМ [3, 2000].
Заключение.
Одно из основных заболеваний современности – атеросклероз – заболевание, вызванное нарушением обмена липидов, в частности липопротеидов. Значительная часть населения страдает ожирением, которое, в свою очередь, провоцирует развитие болезней сердечно-сосудистой системы.
Для лечения этих заболеваний необходимо понимание механизма их развития, что невозможно без знания нормальных процессов обмена липидов. Все это показывает актуальность моей темы и ее пользу для меня в моей будущей врачебной практике.
ПРИЛОЖЕНИЕ.
Работа №1. Исследование эмульгирующих свойств ПАВ желудочно-кишечного тракта.
Принцип метода: ПАВ снижают поверхностное натяжение липидных частиц в водной среде и они распадаются на более мелкие, образуя эмульсию с размером частиц до 0,5 мк, в результате площадь соприкосновения с липолитическими ферментами увеличивается и скорость липолиза возрастает.
Ход работы: В чистой пробирке приготовить смесь ПАВ, состоящую из: 0,5 мл 1% р-ра мыла; 0,5 мл 1% р-ра кислого углекислого натрия; 0,5 мл 1% р-ра белка; 3,0 мл желчи. Эту смесь , близкую к той которая эмульгирует липиды в кишечнике, использовать в качестве одного из эмульгирующих средств. Для исследования взять 6 чистых пробирок и заполнить их реагентами как указано в таблице.
| Реагенты (мл) | 1 | 2 | 3 | 4 | 5 | 6 |
| Желчь | 2,0 | |||||
| Смесь ПАВ | 2,0 | |||||
| Р-р NaНСО3 | 2,0 | |||||
| Р-р мыла | 2,0 | |||||
| Р-р белка | 2,0 | |||||
| Н2О | 2,0 | |||||
| Масло | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 |
Все пробирки встряхнуть до образования эмульсии, отметить время расслоения, сделать вывод об эффективности эмульгаторов, расположив их в ряд в порядке убывания.
Вывод: в результате проделанного опыта я установила, что наилучшей эмульгирующей способностью обладает р-р NаНСО3, затем идет смесь ПАВов, потом – желчь, далее р-р мыла и замыкает этот ряд р-р белка и Н2О.
Т.к. у нас в организме очень мало оснований, то отсюда следует , что наилучшей эмульгирующей способностью в ЖКТ обладает смесь ПАВов, куда также входят и основания.
Работа №2. Влияние желчи на активность панкреатической липазы.
Принцип метода: При воздействии липазы на эмульгированные жиры молока происходит их гидролиз, который можно наблюдать по обеспечиванию розовой окраски предварительно добавленного фенолфталеина, вследствие подкисления реакционной среды освобождающимися высшими жирными кислотами.
Сравнить скорость липолиза в присутствии и отсутствии желчи в реакционной среде. Сделать соответствующий вывод.
Ход работы: В две чистые пробирки поместить реагенты как указано в таблице.
| Реагенты | 1 | 2 |
| Молоко (мл) | 2,0 | 2,0 |
| 20%водный р-р желчи – (мл) | ¾ | 2,0 |
| Н2О | 0,2 | ¾ |
| 1% р-р фенолфталеина (кап) | 4 | 4 |
| Р-р панкреатина – 6 мг ¤ мл (мл) | 0,2 | 0,2 |
1 н и 0,5 н р-ры NаОН, до слабо-розового цвета (по 2 кап).
Вывод: в ходе работы я еще раз подтвердила тот факт, что желчь активирует панкреатическую липазу.
Клинико – диагностическое значение: При нарушении поступления желчи в 12-ти перстную кишку нарушается переваривание и всасывание липидов, в том числе незаменимых. Это сопровождается стеатореей, развитием гиповитаминозов жирорастворимых витаминов , поступлением желчных пигментов в кровь и мочу с последующим развитием желтухи.
Список литературы.
1. Васьковский В. Е. Липиды. ¤¤ Соросовский образовательный журнал. – 1997. №3. С. 32 – 33.
2. Горшкова С. М., Курицин И. Т. Механизмы желчевыделения. – Л. “Наука”. – 1980 . – 287 с.
3. Зайчик А. Ш., Чурилов Л. П. Основы общей патологии. Часть 2. Основы патохимии (учебное пособие для студентов медицинских ВУЗов) – СПб. – 2000. – С. 132 – 155.
4. Иванченкова Р. А. Нейрогуморальная регуляция процессов желчеобразования и желчевыделения. ¤¤ Клин. Медицина. – 1986, т. 64 № 4. С. 27 – 29.
5. Климов А. Н., Никуличева Н. Г. Липиды, липопротеиды, атеросклероз. – СПб. “Питер”. – 1999. – С. 36 – 48.
6. Кононов Е. И., д. м. н. Лекция “Обмен липидов” . Архангельский медицинский институт, 1999 – информация из I-net (www. lipid.ru).
7. Мазурин А. В., Воронцов. И. М. Пропедевтика детских болезней – СПб: ИКФ “Фолиант”. – 1999. – С. 836.
8. Маршал В. Дж. “Клиническая биохимия” ¤ Пер. с англ. – М. – СПб.: “Издательство бином” – “Невский Диалект”, 2000. С. 260.
9. Сорока Н. Ф. Питание и здоровье. – Минск . “Беларусь”. – 1994. – С. 44.
10. Трубачев С. Д. Лекция “Обмен липидов”. УГМА. 2001.
11. Уайт А.., Хэндлер Ф., Смит Э. Основы биохимии. – М: Мир. – 1981. т.№3, С.1359 – 1361.
12. Холодова Ю. Д. , Чаяло П. П. Липопротеины крови. – Киев “Наука думка”. – 1990. – С. 82 – 84 .
Похожие работы
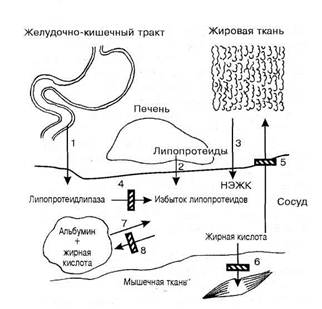
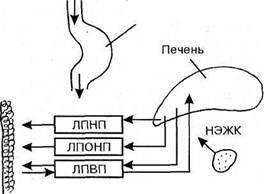
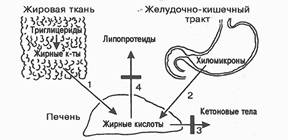
ального переваривания и всасывания их в кишечнике определяющее значение имеет взаимодействие таких факторов, как: 1) выработка поджелудочной железой липолитического фермента липазы; 2) поступление с желчью желчных кислот, эмульгирующих жиры и продукты их распада, активирующих панкреатическую липазу и участвующих во всасывании жирных кислот (всасывается комплекс жирных и желчных кислот); З) ...

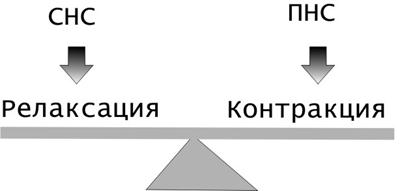

... , лактазная недостаточность Гиперэкссудация Неспецифический язвенный колит, болезнь Крона толстой кишки Повышенная скорость транзита по кишечнику Синдром раздраженного кишечника, функциональная диарея, эндокринные дискинезии Значение нарушения метаболизма желчных кислот в патогенезе хронической диареи В проксимальных отделах тонкой кишки желчные кислоты, экскретируемые печенью, ...
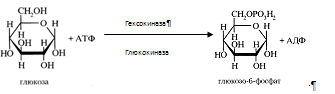
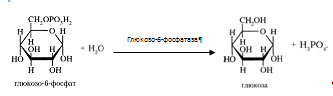
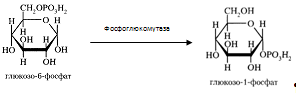

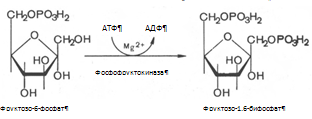
... 3) гепатодепрессии (печёночно-клеточной недостаточности, малой недостаточности печени, недостаточности синтетических процессов); 4) воспаления; 5) шунтирования печени; 6) регенерации и опухолевого роста. При подозрении на конкретную патологию учитываются основные биохимические синдромы, характерные для данного заболевания. За основу берётся стандартная программа функционального обследования, но ...
... ревматизма обусловила значительное снижение заболеваемости — до 0Д8 на 1000 детского населения. В разработку проблемы детского ревматизма внесли большой вклад отечественные педиатры В. И. Молчанов, А. А. Кисель, М. А, Скворцов, А. Б. Воловик, В. П. Бисярина, А. В. Долгополова и др. Эпидемиология, Установлена связь между началом заболевания и перенесенной стрептококковой инфекцией, в основном в ...
0 комментариев