Введение в квантовую фононную биологию
С. Н. Семёнов
Резюме: В рамках “Молекулярно-механической модели строения и функционирования биологических мембран” рассмотрены основные квантово-механические аспекты функционирования мембран. Показано, что биологические мембраны – квантовые системы и квантами, передающими взаимодействия в этих системах, являются фононы – звуковые кванты, а мембрана, как единая квантовая система, характеризуется своими специфическими квантовыми энергетическими уровнями, обусловленные их липидным составом и особенностями молекулярной структуры мембранных белковых систем. Обсуждается фононной механизм передачи внутриклеточной информации, в частности нервного импульса и способы его дестабилизации.
Рассматривая любые внешние воздействия на живой организм нужно всегда помнить, что он состоит из клеток, пусть даже это будет одноклеточный организм, но он тоже состоит их клетки – из одной. Поэтому, нельзя не учитывать внешнего воздействия на клеточном уровне. Более того, на уровне клеточной мембраны, являющейся уникальным образованием, т.к. именно где-то в мембране и проходит граница между живым и неживым – граница между принципиально разными системами. Вероятно, что это единственная известная нам структура с такими уникальными свойствами, но это не единственная уникальность биомембран.
Не будет преувеличением сказать, что биологические (клеточные) мембраны являются одной из важнейших структур, ответственных за обеспечение основных процессов жизнедеятельности клетки. Прежде всего, они отделяют живую клетку от её окружения. Мембраны первыми сталкиваются с различными внешними воздействиями и реагируют на них, они обеспечивают поступление в клетку (и клеточные органеллы) всех необходимых веществ и вывод из клетки продуктов её жизнедеятельности. Не смотря на многообразие выполняемых операций, мембраны характеризуются общими структурными и функциональными чертами. Все они построены, в основном, из липидов и белков. При этом липиды служат основным изолирующим и структурообразующим компонентом всех биологических мембран. А белковые молекулы, главным образом, ответственны за выполнение мембранами многообразных функций присущих живым клеткам (1 – 3). Толщина самой мембраны (40 – 60 Ǻ) меньше суммы линейных размеров 2-х образующих её молекул липида, расположенных по обе её стороны. В тоже время биологические мембраны являются весьма динамичными структурами, т.к. входящие в их состав молекулы находятся в непрерывном движение (4 – 8). Но, не смотря на такую подвижность, липидная часть мембраны сама по себе является превосходным диэлектриком и просто отличным барьером, препятствующим свободному проникновению через неё различных молекул. Более того, такой тонкий и динамичный барьер, видимый только в электронный микроскоп, обладает очень сложной внутренней структурой. В мембране, с точки зрения термодинамики и молекулярной физики, можно обнаружить участки с “кристаллической” структурой (9), характерной для твердых кристаллических тел, в то же время, центральная часть мембраны обладает “неупорядоченным” состоянием, близким по свойствам к жидкостям или, быть может, даже газам. Конечно, нужно понимать, что это говорится с известной долей упрощения, сделанного для облегчения понимания сути материала.
В последнее время вновь возник большой интерес к установлению корреляции механических свойств различных мембран с особенностями их строения и функционирования, обусловленный обнаружением широкого круга механозависимых явлений в биологических мембранах, в частности, связанных с различными каналами, включая ионные каналы в мембранах нервных клеток (10 – 16). Для объяснения наблюдаемых явлений была предложена “Молекулярно-механическая модель строения и функционирования биологических мембран” (в дальнейшем Модель) (17, 18). Модель позволяет описывать многие наблюдаемые структурные и функциональные свойства биомембран не только качественно, но и количественно. Более того, Модель предсказывает, что межмолекулярные взаимодействия в мембране должны сопровождаться излучением или поглощением квантов “механического взаимодействия” – фононов. Многие эффекты вокруг нас выглядят классическими потому, что классические физические законы на самом деле базируются на квантовой механике (19). Необходимость учета квантовых явлений возникает в последнее время при рассмотрении различных аспектов молекулярной биологии, например, генома (20). Данная работа посвящена описанию роли квантовых явлений при рассмотрении структуры и функций биологических мембран.
Модель показывает, что при рассмотрении строения и функционирования мембранных белков необходимо учитывать латеральное сжатие, которое они испытывают со стороны окружающей мембраны. Экспериментально подтверждено, что разные биологические мембраны характеризуются разным поверхностным давлением и сжимаемостью, обусловленными различиями в их липидном составе (17.1, 18.1). Под поверхностным давлением и сжимаемостью надо понимать именно давление и сжимаемость в их классическом физическом значении. При этом межмолекулярные взаимодействия в мембране можно описывать как взаимодействие молекул двумерного квазигаза (17.2, 18.2), который и создаёт внутримембранное латеральное давление. Отсюда сразу становится понятной необходимость поддержания постоянной температуры тела различных животных. Постоянная температура означает постоянство внутримембранного давления и, следовательно, постоянство добавочной свободной энергии сжатия, получаемой внутримембранной молекулой со стороны её окружения. Более подробно это рассмотрено в разделе “Квазистатика” (17.3) или “Quasistatic” (18.3). В противном случае, при непостоянной температуре, клетка будет нестабильно функционировать или ей придётся менять липидный состав своей клеточной мембраны, для поддержания необходимого поверхностного давления и сжимаемости, что бы не менялась добавочная свободная энергия мембранных белков, и соответственно – их структура и функциональные особенности, что и наблюдается на опыте (21 – 23).
Зависимость структуры и свойств мембранных белков от механических свойств мембраны позволило предположить, что процесс функционирования белковых систем сопровождается поглощением или излучением квантов этого взаимодействия – фононов. Т.е. в процессе активации мембранных белков под действием внешнего воздействия, например, при связывании мембранного фермента с субстратом из окружающего раствора, и образованием субстрат-ферментного комплекса выделяется энергия. Которая затем, при возвращении белковой молекулы и исходное (невозбуждённое) состояние может быть излучена в виде мембранного фонона. Выделившийся фонон может потом активировать иную белковую системы в мембране, будучи поглощенным ею, т.е. осуществив передачу информации вдоль мембраны и координацию деятельности различных мембранных белковых систем. Причём совсем необязательно, что бы эти мембранные системы были пространственно близки между собой или имели иную “видимую” связь. Скорее, не будет видимого сопряжения различных мембранных белковых систем между собой. Мы просто будем наблюдать событие, обычно вызывающее некий эффект (или эффекты), в том числе и иной природы, в некотором отдалённом участке биомембраны. Главное, чтобы в мембране существовала упорядоченная структура, похожая на структуру кристалла, обеспечивающая условия передачи фононной информации, а такие структуры, как отмечено выше, существуют в биологических мембранах. В противном случае, фонон может просто рассеяться в мембране, потратив свою энергию на её нагрев. Иными словами, при облучении клетки фононами с разными частотами (энергиями) будут обнаружены пики поглощения ультразвука, соответствующие собственным квантовым энергетическим уровням данной биомембраны. Можно назвать это и пиками “резонансного” поглощения ультразвука, когда его частота совпадает с частотой собственных мембранных фононов – полная аналогия с традиционным спектральным анализом химических соединений. Необходимо подчеркнуть, что речь идёт о биомембранах живых клеток, т.к. именно живая клетка стремится поддерживать постоянство своих параметров при изменении окружающих условий. Если клетке не удаётся это сделать, то она погибает.
Для проверки высказанного предположения были сконструированы спектрофононометры и методики их применения (24, 25). Затем определены ультразвуковые спектры мембран различных живых клеток. Предварительно пришлось разработать способ измерения поверхностного давления различных биологических мембран (17.1, 18.1). Эти величины были необходимы для количественной оценки ожидаемых явлений, как и результаты по оценки латеральной сжимаемости биомембран при поверхностных давлениях, равных давлениям реальных мембран, которые были легко получены на основании липидного состава различных клеток и литературных данных по сжатию липидных монослоёв различного состава на границе раздела полярной и неполярной фаз. На рис. 1 приведён типичный образец ультразвукового спектра живых клеток (спектр поглощения ультразвука в относительных единицах с.u. для водной суспензии микроорганизмов Bac. subtilis).
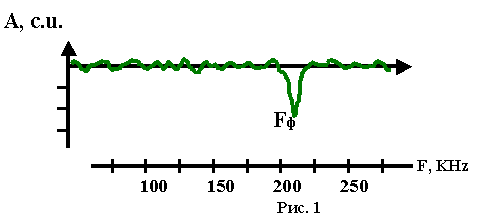
Часть ультразвукового спектра клеток Bac. subtilis в водной суспензии (физиологический раствор).
Естественно, что, так как механические параметры клеточной мембраны уникальны для различных клеток, то и фононные спектры разных клеток уникальны и могут служить для их идентификации. Таким образом, биологические мембраны – квантовые системы и квантами, передающими взаимодействия в этих системах, являются фононы – звуковые кванты. Именно фононы осуществляют межмолекулярный обмен сигналами внутри биологических мембран. Причём обмен сигналами и, соответственно, координация деятельности происходит не только между соседними молекулами, но и молекулами находящихся в разных участках биомембраны. Иными словами, мембраны, как единые квантовые системы, характеризуются своими специфическими квантовыми энергетическими уровнями (с точностью до kT, где k – постоянная Больцмана, а Т – температура в градусах Кельвина), обусловленные их липидным составом и особенностями молекулярной структуры мембранных белковых систем.
Надо отметить, что фононные спектры в биологии могут играть ту же роль, что и обычная, традиционная, фотонная спектроскопия в различных областях науки и техники при исследовании и идентификации разнообразных химических соединений. Активируя или инактивируя те или иные системы в биологических мембранах, мы будем менять и их функционирование, то есть, воздействовать на свойства клеток и их состояние. Самое простое, что можно сделать с клетками при этом – уничтожать выбранные клетки, не затрагивая других клеток в системе (например – раковые клетки).
Многие аспекты традиционной оптической спектроскопии справедливы и для предлагаемой “ультразвуковой”, фононной спектроскопии. Правда, надо учитывать, что фононы и фотоны отличаются не только одной буквой, но и относятся ещё и к разным типам квантовых частиц. Одни – Бозоны, другие – Фермионы и, следовательно, имеют некоторые различия в своём поведении.
В рамках фононной квантовой механики биомембран становятся понятными давно известные многочисленные факты, когда одинаковое ультразвуковое воздействие на разные клетки приводит к различным результатам, иногда противоречащим друг другу (26, 27). При традиционных исследованиях биологических объектов авторы не учитывали возможность “резонансного поглощения” ультразвука мембранными белками, которое может менять структурное и функциональное состояние последних, хотя и не отрицали, что поглощение ультразвука обусловлено белками и нуклеиновыми кислотами (28 – 30). Вероятно, с этим связаны прямо противоположные оценки влияния клеток на распространение ультразвука в структурированных и неструктурированных (лизированных или гомогонезированных) образцах тканей животных и человека (31 – 33).
Фонон – квант внутриклеточной информации
Одной из актуальных проблем современной клеточной биологии является изучение путей и способов распространение информации (обмен сигналами) внутри и между клетками. Важность этого вопроса очевидна – это и вопросы функционирования нейронов и всей нервной системы в целом и, соответственно, работы подконтрольных им иных клеточных систем организма, как в норме, так и при различных патологиях, где обнаружены изменения активности медиаторных систем и их рецепторов, а также связанные с этим нарушения мембран и ионных каналов нейронов. Интересно, что периферические органы и ткани обладают некоторой автономностью, т.е. способностью ограниченно функционировать в отсутствие нейрогуморальных регуляторных влияний. “Изолированные” таким образом органы работают на минимальном режиме их функциональных возможностей благодаря внутри- и межклеточным регуляторам (34).
Кроме того, в последние годы появились данные, что клетка, неадекватно реагирующая на “социальные сигналы”, поступающие от других клеток организма, может дать начало злокачественной опухоли (35).
Наиболее полно исследованы химические пути обмена межклеточной информацией при помощи нейомедиаторов и иных химических соединений. В настоящее время известно не менее 74 различных химических реакций, разделённых, как минимум, на 25 групп, в которых задействованы те или иные молекулы, принимающие участие в передаче межклеточной информации (36). Отмечено, что эти группы реакций можно рассматривать как “векторные реакции”. Т.е. это однонаправленные пути.
Гораздо меньше известно о процессах трансформации внемебранного – обычно химического, но не обязательно (в начале работы упомянуто о чувствительности биомембран к механическим воздействиям) – сигнала во внутримембранный сигнал. Под сигналом понимается любое внешнее воздействие, вызывающее изменения в клеточной мембране и/или уже внутри самой клетки, в её цитоплазме. Более подробно вопросы трансформации внешнего сигнала во внутримембранный и происходящие при этом процессы будут рассмотрены в отдельной работе (37). В данной работе рассмотрим только возможный механизм быстрого распространения сигнала, полученного клеткой в одном месте до его получателя (адресата), расположенного на другом, пространственно удалённом участке биомембраны. Самый известный процесс такого рода – распространение нервного импульса в нейронах.
Первой стадией таких трансформаций является связывание молекулы сигнала с соответствующей ей молекулой рецептором, расположенной на поверхности клетки. Для такого связывания сигнальная молекула и её рецептор должны специфично соответствовать друг другу (избитое, но, в общем, верное сравнение – подходить друг к другу, как ключ к замку). Всякая клетка имеет специфичный для нее набор рецепторов, и этот набор определяет круг химических сигналов, на который эта клетка реагирует. До этой стадии всё понятно; связывание сигнальной молекулы с её рецептором вызывает некие изменения в мембране – происходит трансформация внешнего химического сигнала во внутримембранный сигнал. Как конкретно происходит трансформация внешнего сигнала и во что он трансформируется в биомембране до сих пор не известно. Традиционные подходы не дают ответа. Для мембраны как квантовой системы не так важно, является химическая молекула сигнал ключом для мембранного рецептора или отмычкой. Для квантовой системы более существенно, что самопроизвольная реакция идет с выделением энергии, которая может быть излучена в виде мембранного фонона. Этот процесс схематично изображен на рисунке в работе, описывающей Модель, в разделе “Квазидинамика (Quasidynamics)” (17.4, 18.4). Естественно, что обратная реакция – отделение молекулы сигнала от молекулы мембранного рецептора – будет идти только с поглощением энергии, которой может быть энергия поглощённого комплексом рецептора с сигнальной молекулой мембранного фонона. Это можно представить в виде простых уравнений, принципиально описывающих процесс, без детализации:
Прямая реакция:
(внешний химический сигнал) + (мембранный рецептор) è
(комплекс рецептора с молекулой сигналом) +
(мембранный фонон = ħω);
Обратная реакция:
(комплекс рецептора с молекулой сигналом) +
(мембранный фонон = ħω) è
(мембранный рецептор) +
(химический сигнал, выделенный из мембраны).
Переименование прямой реакции в обратную и наоборот сути процесса не меняет. Более того, если вместо химического сигнала будем рассматривать любой другой сигнал, воздействующий на мембранный рецептор, то уравнения (и процессы, которые они описывают) не изменятся. Возможно только одно отличие в схемах: внешний сигнал вызывает активацию рецептора, его переход в возбужденное состояние с последующей релаксацией в исходную форму, которая и сопровождается излучением мембранного фонона. Естественно, что здесь может и не образовываться “относительно стабильный комплекс рецептора с внешним сигналом” и, следовательно, отсутствует обратная реакция – вся схема только упрощается.
Хорошо известно, что есть специализированные клетки, задача которых заключается в максимально быстрой и точной передачи сигналов от разных частей организма в мозг и обратно. Речь идет о нервных клетках (нейронах), связывающих мозг со всеми частями организма. Нейроны отличаются разнообразием, но всем им присуще наличие отростков. Короткими отростками (дендритами) нервные клетки контактируют друг с другом. Кроме коротких отростков нейрон обладает ещё и длинным отростком, называемым аксоном, по которому нервные импульсы идут от тела клетки к иннервируемым органам и другим нервным клеткам. Аксоны некоторых клеток тянутся на 50 – 70 см. Аксоны покрыты миелиновой оболочкой белого цвета. Миелиновая оболочка состоит из белков (миелина) и липида. Поверх миелиновой оболочки есть ещё швановская оболочка. Миелиновая оболочка, являясь изолятором, предотвращает рассеивание нервных импульсов и их переход на другие нервные волокна. Миелиновое покрытие по длине волокна имеет сегментарное строение; на границе двух сегментов имеются участки безмиелиновых перетяжек – так называемые узлы нервного волокна или перехваты Ранвье. За счет этого нервный импульс распространяется по волокну не непрерывно, а скачками: электрические импульсы “перепрыгивают” от одного перехвата Ранвье к другому. Скорость передачи нервного импульса по аксонам меняется от 150 м/сек для человека до 50 м/сек для лягушки (38 – 40).
Реально, как распространяется непосредственно сам нервный импульс на основании приведённых результатов говорить некорректно. В экспериментах измеряется так называемый “потенциал действия”, последовательно регистрируемый в разных перехватах Равнье после раздражения нейрона. Вот именно этот “потенциал действия”, и сопутствующие ему электрические процессы, распространяется по аксону скачками. Но то, что электрические явления сопровождают передачу сигнала нейроном, совсем не означает, что эти явления сами непосредственно и являются внутриклеточными информационными сигналами. (Очень это напоминает ёлочную гирлянду, там тоже внешне кажется, что разноцветные светлячки перепрыгивают из одного запаянного стеклянного шарика – лампочки, если кто не знает – в другой.) Здесь наблюдается предсказанный выше в данной статье квантовый процесс фононного обмена между различными мембранными белковыми системами в мембране аксона нервной клетки, когда воздействие в одном месте мембраны вызывает ответ в другом месте, причём воздействие и ответ могут иметь различную физико-химическую природу: общее между ними – квантово-механическое сопряжение через фононный обмен.
Нервная система, как и любая иная система, предназначенная для передачи информационных сигналов, должна удовлетворять ряду очевидных требований. Она должна быстро и без искажений передавать информационный сигнал. Желательно, чтобы система была универсальной, т.е. могла свободно передавать сигналы в противоположных направлениях и т.д. Кроме того, необходимо, чтобы в то время, когда сигналы не передаются – нервная клетка находится в покое – система потребляла минимум энергии, и самопроизвольно “автоматически” возвращалась в исходное работоспособное состояние после передачи сигналов. Ну и естественно, что система должна легко “обслуживаться”, что бы все необходимое для её нормального функционирования достаточно быстро и адресно попадало туда, где это всё необходимое требуется, создавая минимум помех процессу передачи полезного информационного сигнала. Проше всего это сделать, разделив процессы передачи информации и обслуживания в пространстве в пространстве.
Для снижения искажения и затухания фононного сигнала необходима регулярная структура с минимумом “посторонних включений”. Структура миелиновой оболочки, отличающаяся от других биомембран высоким содержанием липидов и низким – белков, удовлетворяет этому условию (40 – 42). Очевидно, что эта структура позволяет фонону двигаться в любом направлении – нет никакой разницы, что двигаться в одну строну, что – в противоположную. Любая биомембрана является замкнутой, поэтому, в какую бы сторону не был излучён фонон, он всё равно может достигнуть самого удалённого, от места своего зарождения, участка мембраны своей клетки, если только раньше не достигнет своего адресата и не будет поглощен им, потратив свою энергию на активацию данной белковой системы.
Сигнал при распространении может подвергаться затуханию и/или искажениям, поэтому на линиях передачи сигналов необходимо периодически иметь усилители, способные подкорректировать, усилить полученный сигнал и передать его дальше. Естественно, что в случае биомембраны это будут молекулярные усилители, которые должны автоматически запускаться при получении фононного сигнала и выключаться после того, как передали его дальше. Очевидно, что процесс усиления сигнала идёт с затратой энергии, которая должна быть запасена клеткой предварительно, желательно, в легкодоступной форме, при этом усилитель должен контактировать с окружающей мембрану средой, иначе ему просто неоткуда получать внешнюю энергию для своей работы. Такие периодические системы реально существуют на поверхности аксонов – это уже упомянутые перехваты Ранвье. Фонон, в процессе своего движения по аксону, периодически активирует квантовые усилители, которые должны быть расположены в перехватах Ранвье. Эти квантовые усилители включаются для усиления полученного сигнала, а затем, передав его дальше, выключаются. Таким образом, нервный импульс никуда и нигде не прыгает, а спокойно распространяется по аксону, включая для своего усиления периодически расположенные в мембране квантовые усилители, которые, передав фононный сигнал дальше, затем выключаются. Иными словами, фонон достигая мембранного квантового усилителя, поглощается последним. Такое поглощение фонона означает, что усилитель получает порцию энергии, переносимую квантом звука. Получив энергию, усилитель активируется и передаёт усиленный (и скорректированный) сигнал в виде нового фонона. Этот процесс должен сопровождаться использованием энергии. В данном конкретном случае это трансмембранный электрохимический градиент – заранее запасённая энергия. После чего, отдав энергию, инактивируется, т.е. просто выключается. Следовательно, молекулярный усилитель должен содержать механозависимую часть, которая должна выключаться под действием сжатия, обусловленного наличием латерального поверхностного давления мембраны. Именно эта механозависимость и наблюдается в мембране нейронов для ионных каналов (10). Вполне возможно, что канал буквально схлопывается под действием сжатия со стороны своего окружения. Т.е. имеем, что молекулярный усилитель активируется под действием сигнала, который необходимо усилить и передать дальше, а затем, выполнив свою функцию, “автоматически” выключается под действием сжатия со стороны окружающей его мембраны. Естественно, что в выключенном (неактивном) состоянии усилитель не нуждается в энергии. А клетка может восстанавливать трансмембранный электрохимический градиент, служащий источником энергии для молекулярного усилителя; просто нужны две независимые молекулярные системы. В настоящее время нет данных для анализа конкретного механизма действия квантового фононного усилителя, роли пассивного и активного транспорта ионов через мембраны в этом процессе и т.д. Кстати, изменения величины трансмембранного электрического поля, характерные для нервных клеток, влияли только на полярные головки липида, но не затрагивали ни структуру, ни динамику углеводородных цепей в мембране (43). Иными словами, процессы, сопровождающие распространения мембранного фононного сигнала не влияли на условия его распространения – не влияли на углеводородный компонент биомембраны.
Для обслуживания телекоммуникационной клеточной системы можно использовать клеточную цитоплазму и внеклеточное пространство, чтобы не создавать помех распространению мембранных фононов. Внеклеточное пространство может играть особенно заметную роль в мозгу, где половина клеток является нейронами, а вторая половина – клетки глии – не заняты непосредственно в передаче и обработке нервных сигналов и могут быть задействованы для обеспечения тех или иных потребностей нейронов. В этом случае должна наблюдаться между ними взаимная информационная связь и взаимовлияние, что и обнаружено в последнее время (44). Такой механизм позволяет разделить пути распространения информации по аксону и химических соединений, необходимых для поддержания жизнедеятельности и работоспособности нервной клетки, как единого целого, так и её частей.
Даже такое краткое рассмотрение показывает, что квантовый фононный вариант предложенной Модели позволяет описать работу различных биологических систем, в том числе и нервной системы живых организмов. Передача информации нервными клетками с помощью фононов позволяет передавать клетке сразу несколько сигналов, причём в разных направлениях наиболее быстрым и простым и единым способом. Если фонон при своём движении в мембране и влияет на последнюю, то только уж в совсем локальной области, никак не влияя на распространение других фононов. Единственное возможное узкое место – ограниченное число усилителей в мембране и время их срабатывания. Предположим, что самая медленная стадия в процессе передачи информации это цикл срабатывания квантового фононного усилителя, равный ~0,002 сек (известные данные (38) о том, что не удаётся пропустить более ~500 нервных импульсов через аксон), то скорость распространения нервного импульса (0,2-0,4 м / 0,002 сек) будет в пределах 100 – 200 м/сек, что близко к известным значениям, приведённым выше. Для биологии это хорошее совпадение, которое получено, при допущении, что нервный импульс передаётся фононами у разных организмов и механизм работы усилителей у них близок. Скорость распространения самого ультразвука в тканях, равная ~ 1600 м/сек (28) и слабо зависящая от вида конкретной биологической ткани (45), не может быть лимитирующей стадией для скорости распространения фононного импульса в аксоне.
Аксон только передаёт информацию, а основное тело нервной клетки, имеющее множество отростков-дендритов, контактирующих с другими нервными клетками, должно анализировать информацию и передавать её определённым адресатам – другим нервным леткам. Естественно, эти более сложные функции должны выполняться более сложными клеточными квантовыми системами. А это потеря скорости передачи информации, что и наблюдают в экспериментах. Но существенно картина не меняется.
В итоге получили, что нервная клетка является аналогом любой телекоммуникационной системы, Клетка подчиняется тем же законам, что и иные системы, например телефонные сети (может быть, более близкий аналог – локальные компьютерные сети и интернет). Имея разные участки нервной телекоммуникационной системы в организме, для функционирования всей системы в целом нужна нормальная работа её частей. Не может быть нормальной работы всей информационной системы, если нарушена работа хотя бы одной из её частей.
В работе “Алкоголь и другие органические растворители” сделан вывод, что, эти соединения ухудшают работу фононопроводящих участков клеточных мембран, нарушая липидную структуру последних (46, 47). Но можно нарушать работу квантовых усилителей и других систем передачи информации, т.е. тех, которые ответственны за её анализ и/или адресацию. Причём это можно сделать, как минимум, двумя способами: перегрузив систему ложными сигналами, когда уже белковые квантовые системы не успевают обрабатывать поступающую информацию, или нарушив нормальную работы самих белковых квантовых систем.
1-я ситуация возникнет, если в мембране образуется избыток фононов. Это можно сделать, облучая организм ультразвуком, и когда, какие-то частоты ультразвука совпадут с собственными частотами клеток, соответственно, они и будут влиять на работу клетки, нарушая, скорее всего, её функционирование. Известно, что случайное вмешательство в работу сложной системы, чаше всего на пользу последней не идёт.
При этом можно перегрузить всю систему избыточными сигналами так, что она просто не сможет функционировать, у молекулярных усилителей просто не будет возможности вернуться в исходное состояние.
Возможно, перегрузить систему частично: где-то она будет успевать срабатывать нормально, а где-то нет. При более низкой интенсивности этого постороннего ультразвука, возникающие фононы будут хаотично распространяться по мембранам в разные стороны, внося хаос в работу управляемых этими нервами органов. Фононов еще мало для полной блокировки молекулярных усилителей. Сигналы распространяются по нейронам хаотично, вызывая хаотичные отклики на них в тканях животных, в том числе и избыточный, хаотичный выброс в межклеточное пространство различных нейромедиаторов – будет наблюдаться “самонаркотизация” организма. Если не удалось перегрузить систему посторонними фононными сигналами полностью, то она может работать, истощая созданный ранее энергетический запас клетки, а затем всё-таки выключится, теперь уже от истощения. Организму потребуется отдых. Теперь сравним эти выводы с экспериментальными наблюдениями.
Ультразвук обычно плохо влиял на поведение животных. У них менялось поведение: некоторые сразу после начала облучения проявляли сильное беспокойство, выраженное в рывках, страх, за которыми уже через короткое время (~ 1 мин.) следовало состояние полной неподвижности. Менялась деятельность внутренних органов, наблюдается состояние, сходное с наркотическим состоянием, когда животные, например, не реагируют на прикосновения. Если в это время облучение прекращали, то часть животных могла ещё прийти в нормальное состояние, иначе – они погибали. Кратковременное облучение лягушек ультразвуком вызывало у них состояние паралича, аналогичное действию яда КУРАРЕ, который блокирует передачу нервного импульса нейронами. Не обнаружили внутренних кровоизлияний и повреждений центральной нервной системы. Полагали, что непосредственная причина смерти заключается в прямом действии ультразвука на центральную нервную систему, которое не вызывает заметных морфологических изменений тканей животных. В пользу этого говорят и данные микроскопического исследования влияния ультразвука на дафнии – рачки зоопланктона. При облучении сначала парализуются конечности, затем жабры, глаза и, наконец, останавливается сердце. В то же время в некоторых случаях, в малых дозах ультразвук стимулировал жизненные процессы некоторых маленьких рыбок (48). При этом большинство данных получено для холоднокровных животных и в жидкой среде. Возможно, это связано с тем, что у них работа молекулярных усилителей в нейронах не так строго зависит от частоты внешнего воздействия, т.е. они срабатывают при поглощении фононов с более широким частотным диапазоном, чем теплокровные. Холоднокровные вынуждены функционировать (и функционируют, пусть “как сонные мухи”, но функционируют) при разных температурах, т.е. при разных величинах латерального поверхностного давлениях их биомембран. Соответственно, мембранные системы их клеток должны обладать работоспособностью, когда добавочная свободная энергия сжатия, действующая на мембранные системы не постоянна, а меняется в некотором диапазоне при изменении температуры.
2-я ситуация возникает при нарушении работы, например, самих молекулярных усилителей. Для этого введем в организм, например, КУРАРЕ. Дальше можно и не рассматривать последствия.
В рамках данной работы невозможно даже частично рассмотреть различные следствия, вытекающие из предложенной Модели. Это будет сделано в следующих работах.
Приведённые выше материалы хорошо описываются в рамках квантово-механической фононной Модели строения и функционирования клеточных мембран, что позволяет утверждать: “ФОНОН – КВАНТ биологической (клеточной) мембраны”. Модель пригодна для объяснения широкого круга наблюдаемых явлений. При этом наблюдаемые явления описываются в рамках единого понятийного аппарата и не требуют специфических допущений для описания внешне разных явлений в живых системах, и вытекающие из Модели следствия могут быть проще и понятнее их традиционных объяснений, например, действия алкоголя на организм (46, 47).
Есть один экстремальный момент в жизни клеток – их деление, где сложно предсказать действие ультразвука на клеточную мембрану. В момент деления мембрана испытывает существенную перестройку: в какой-то момент времени она не является замкнутой системой. В области деления вместо одной непрерывной мембраны получается две, принадлежащие разным клеткам. Естественно, что деление клеток – процесс, характерный для развивающегося организма. Облучение такого организма может привести к непредсказуемым пока в рамках Модели последствиям. Речь идёт об УЗИ плода беременных женщин. Мне не удалось найти более менее достоверных данных о возможной связи патологии в развитии детей и УЗИ плодов. Буду благодарен, если кто-то сможет сообщить такие сведения и/или учтёт результаты, вытекающие из предложенной Модели в своей деятельности. Есть отдельны косвенные признаки, указывающие, что не всё так хорошо с безвредностью УЗИ, как в этом хотят уверить некоторые.
Список литературы
A FLUID LIPID-GLOBULAR PROTEIN MOSAIC MODEL OF MEMBRANE STRUCTURE. S.J. Singer, “Ann. N.Y. Acad. Sci.”, (1972), 195, 16 –23.
MEMBRANE STRUCTURE. Roberson J.D., "J. Cell Biol.", (1981), 91(3, part 2), 189 – 204.
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ, Л.Д. Бергельсон, изд. “Наука”, Москва, 1975.
Kinetic and Thermodynamic Aspects of Lipid Translocation in Biological Membranes. S. Frickenhaus, R. Heinrich, “Biophys. J.”, March 1, (1999), 76(3), 1293 – 1309.
RAPID TRANSPORT OF PHOSPHOLIPIDS ACROSS THE PLASMA MEMBRANE OF Leishmania infantum. Josй M Araъjo-Santos, Francisco Gamarro, Santiago Castanys, Andreas Herrmann, Thomas Pomorski, “Biochem. and Biophys. Res. Commun.” (2003), 306(No 1), 250 – 255, (www.elsevir.com/locate/ebbre)
TRANSMEMBRANE MOVEMENT OF DIETHER PHOSPHOLIPIDS IN HUMAN ERYTHROCYTES AND HUMAN FIBROBLASTS. P. Fellmann, P. Hervй, T. Pomorski, P. Mьller, D. Geldwerth, A. Herrmann, P.F. Devaux, “Biochemistry”, (2000), 39, 4994 – 5003.
TRANSBILAYER MOVEMENT OF FLUORESCENT AND SPIN-LABELED PHOSPHOLIPIDS IN THE PLASMA MEMBRANE OF HUMAN FIBROBLASTS: A QUANTITATIVE APPROACH. T. Pomorski, P. Mьller, B. Zimmermann, K. Burger, P.F. Devaux, A. Herrmann, “J. Cell Sci.” (1996), 109, 687 – 698.
FLUIDITY OF CELL MEMBRANES – CURRENT CONCEPTS AND TRENDS. Shinitzky M., Henkart P., International Review of Cytology, (1979), 60, 121 – 147.
BIOLOGICAL MEMBRANE AS A LIQUID CRYSTAL MECHANISM. Petrov A.G., BPU-5: Fifth General Conference of the Balkan Physical Union, August 25-29, (2003), Vrnjaèka Banja, Serbia and Montenegro, 1737 – 1743.
MODULATION OF MEMBRANE TRAFFIC BY MECHANICAL STIMULI. G. Apodaca, “Am. J. Physiol. – Renal Physiol”, (2002), 284, No 2, pp. F179 – F190.
TECHNIQUES FOR MECHANICAL STIMULATION OF CELLS IN VITRO: A REVIEW. Brown N.D., “J. Biomech.” (2000), 33, 3 – 14.
MECHANORECEPTION AT THE CELLULAR LEVEL: THE DETECTION, INTERPRETATION, AND DIVERSITY OF RESPONSES TO MECHANICAL SIGNALS. Banes A.J., Tsuzaki M., Yamamoto J., “Biochem. Cell Biol.”, (1995), 73, 349 – 365.
CELL SWELLING-INDUCED ATP RELEASE AND GADOLINIUM-SENSITIVE CHANNELS. F. Boudreault, R. Grygorczyk, “Amer. J. Physiol.” (2002), 282(1), C219 – C226, (www.ajpcell.org)
SYNAPTIC TRANSMISSION AT VERTEBRATE HAIR CELLS. Fuchs P.A., “Curr. Opin. Neurobiol.” (1996), 6, 514 – 519.
MECHANOTRANSDUCTION IN VERTEBRATE HAIR CELLS: STRUCTURE AND FUNCTION OF STEREOCILIARY BUNDLE. Hackney C.M., Furness D.N., “Am. J. Physiol. Cell Physiol.” (1995), 268, C1 – C13.
MECHANOELECTRICAL TRANSDUCTION BY HAIR CELLS. Howard J., Roberts W.M., Hudspeth A.J., “Annu. Rev. Biophys. Biophys. Chem.” (1988), 17, 99 – 124.
Молекулярно-механическая модель строения и функционирования биологических мембран. ВВЕДЕНИЕ В КВАНТОВУЮ ФОНОННУЮ БИОЛОГИЮ. Семёнов С.Н., Internet: SciTecLibrary.com. (http://www.sciteclibrary.ru/rus/catalog/pages/6013.html)
Introduction to quantum phonon biology – THE molecular mechanical model of structure and functions of biological membranes. semenov S.N., Internet: SciTecLibrary.com. (http://www.sciteclibrary.ru/eng/catalog/pages/6646.html)
Quantum engineering: Protecting the quantum world. PAZ J.P., “Nature”, (2001), 412, 869 – 870.
A Dynamical Theory Describing Superconductant DNA. Pensinger W., Paine D., “Int. J. Quantum Chem.”, (1979), Vol. XV, 333-341.
EFFECT OF LIPID COMPOSITION ON STABILITY OF CELLULAR MEMBRANES DURING FREEZE-THAWING OF Lactobacillus acidophilus GROWN AT DIFFERENT TEMPERATURES. Fernбndez Murga M.L., Font de Valdez G., Disalvo E.A., “Arch. Biochem. and Biophys.” (2001), 388(1), 179 – 184, (http//www.idealibrary.com).
STRUCTURE OF MEMBRANES LIPIDS AND PHYSICO-BIOCHEMICAL PROPERTIES OF PLASMA MEMBRANES FROM Thermoplasma Acidophilum, ADAPTED TO GROWTH AT 37oC. Yang L.L., Hang A., “Biochim. Biophys. Acta”, (1979), 573(2), 308 – 320.
MEMBRANES CHANGES DURING GROWTH OF Tetrahymena IN THE PRESENCE OF ETHANOL. Nandini-Kishore S.G., Mattox S.M., Martin Ch.E., Thomson G.A., “Biochim. Biophys. Acta”, (1979), 551, 315 – 327.
THE ACOUSTICAL METHODS AND APPARATUS FOR IDENTIFICATION AND SELECTIVE TREATMENT OF A CELLULAR SYSTEM. Semenov S.N., Int. Patent Application PCT/CZ01/00046, (dated 26.08.2001).
POPULAR DESCRIPTION OF INVENTIONS “THE ACOUSTICAL METHODS AND APPARATUS FOR IDENTIFICATION AND SELECTIVE TREATMENT OF A CELLULAR SYSTEM”. Semenov S.N., Internet: SciTecLibrary.com. (http://www.sciteclibrary.ru/eng/catalog/pages/6554.html),
УЛЬТРАЗВУК И ЕГО ПРИМЕНЕНИЕ В НАУКЕ И ТЕХНИКЕ. Л. Бергман. “Иностранная Литература”, (1957) Москва.
DER ULTRASCHALL UND SEIN ANWENDUNG IN WISSENSCHAFT UND TECHNIK. Dr. L. BERGMAN, 6 vцllig ьberarbeitete und erweiterte Auflage mit 609 Bildern, Zьrich, 1954.
БИОФИЗИЧЕСКИЕ ОСНОВЫ УЛЬТРАЗВУКОВОЙ МЕДИЦИНСКОЙ ДИАГНОСТИКИ. А.П. Сарвазян, “УЛЬТРАЗВУКОВАЯ ДИАГНОСТИКА. Сборник трудов”, (1983), Институт прикладной физики АН СССР, г. Горький, 80 – 95.
DEPENDENCE OF THE ULTRASONIC PROPERTIES OF BIOLOGICAL TISSUE ON CONSTITUENT PROTEINS. Goss S.A., Frizzell L.A., Dunn F. “J. Acoust. Soc. Am.” (1980), 67, 1041 – 1044.
ACOUSTIC PROPERTIES OF NORMAL AND CANCEROUS HUMAN LIVER. I. DEPENDENCE ON PATHOLOGICAL CONDITION. Bamber J.C., Hill C.R., “Ultrasound Med. Biol.” (1981), 7, 121 – 133.
MECHANISM OF ABSORPTION OF ULTRASOUND IN LIVER TISSUE. Pauly H., Schwan H.P., “J. Acoust. Soc. Am.” (1971), 50, 692 – 699.
ULTRASONIC ATTENUATION AND BACKSCATTERING BY MAMMALIAN ORGANS AS A FUNCTION OF TIME AFTER EXCISION. Bamber J.C., Fry M.J., Hill C.R., Dunn F., “Ultrasound Med. Biol.” (1977), 3, 15 – 20.
КОРРЕЛЯЦИЯ АКУСТИЧЕСКИХ ХАРАКТЕРИСТИК И ВЕРОЯТНОСТИ ВОЗНИКНОВЕНИЯ ОПУХОЛИ ПЕЧЕНИ МЫШЕЙ. Маленков А.Г., Асоян К.В., “Биофизика” (1983), 28, 326 – 329.
НАРУШЕНИЯ МЕХАНИЗМОВ НЕЙРОГУМОРАЛЬНОЙ РЕГУЛЯЦИИ ВИСЦЕРАЛЬНЫХ ФУНКЦИЙ ОРГАНИЗМА В ПОСТ РЕАНИМАЦИОННОМ ПЕРИОДЕ. Лобов В.В., Диссертация на соискание ученой степени доктора медицинских наук (14.00.16. - патологическая физиология), (1998), г. Челябинск, (Интернет: http://www.ic.omskreg.ru/~metabolism/lobov/index.htm).
СОЦИАЛЬНОЕ ПОВЕДЕНИЕ НОРМАЛЬНЫХ КЛЕТОК И АНТИСОЦИАЛЬНОЕ ПОВЕДЕНИЕ ОПУХОЛЕВЫХ КЛЕТОК. 1. СИГНАЛЬНЫЕ МОЛЕКУЛЫ, ВЫЗЫВАЮЩИЕ РАЗМНОЖЕНИЕ И ГИБЕЛЬ КЛЕТОК. Васильев Ю.М., “БИОЛОГИЯ. Соросовский образовательный журнал”, (1997), МГУ им. М.В. Ломоносова.
THE CHEMICAL ORGANIZATION OF SIGNALING INTERACTIONS. U.P. Bhalla, “Bioinformatics”, (2002), 18(6), 855 – 863. (http://www.ncbc.res.in/~bhalla/examples/EGFR_example.html)
ТРАНСФОРМАЦИЯ ВНЕШНЕГО СИГНАЛА ВО ВНУТРИМЕМБРАННЫЙ И ВОЗМОЖНЫЕ МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ЭТОГО ПРОЦЕССА. Семёнов С.Н., Рабочее название статьи, готовящейся для публикации на Интернет сайте SciTecLibrary.com.
НЕРВНАЯ СИСТЕМА. “Детская Энциклопедия”, Академия Педагогических Наук РСФСР, т.6, стр. 159 – 173, Москва, 1960.
IONIC PROCESSES IN EXCITABLE MEMBRANES. Vaccaro S.R., Green H.S., "J. Theor. Biol.", (1979), 81(4), 771 – 802.
COMPARATIVE DIFFRACTION STUDIES ON MYELIN MEMBRANES. Kirshner D.A., Caspar D.L.D., "Ann. N.Y. Acad. Sci." (1972), 195, 309 – 320.
CONSTITUTION AND PROPERTIES OF AXONAL MEMBRANES OF CRUSTACEAN NERVES. M. Balerna, M. Fosset, R. Chicheportiche, G. Romey and M. Lazdunski, "Biochemistry", (1975), 14, 5500 – 5511.
X-RAY DIFFRACTION STUDY OF KINETIC OF MYELIN LATTICE SWELLING. EFFECT OF DIVALENT CATIONS. Padron R., Mateu L., Kirschner D.A., "Biophys. J." (1979), 28, 321 – 240.
ELECTRIC FIELD EFFECTS ON LIPID MEMBRANE STRUCTURE. Stulen G. “Biochim. Biophys. Acta” (1981), 640, 621 – 627.
NEW INSIGHTS INTO NEURON-GLIA COMMUNICATION. R. D. Fields, B. Stevens-graham, “Science”, (2002), 298, No 5593, pp. 556 – 562.
УЛЬТРАЗВУКОВАЯ ИНТЕРФЕРОМЕТРИЯ МЯГКИХ БИОЛОГИЧЕСКИХ ТКАНЕЙ. В.А. Клемин, А.П. Сарвазян, “УЛЬТРАЗВУКОВАЯ ДИАГНОСТИКА. Сборник трудов”, (1983), Институт прикладной физики АН СССР, г. Горький, 95 – 105.
Алкоголь и другие органические растворители. Введение в квантовую фононную биологию. С.Н. Семёнов, Internet: SciTecLibrary.com. (http://www.sciteclibrary.ru/rus/catalog/pages/6518.html)
Alcohol and other organic solvents – Introduction to quantum phonon biology. S.N. Semenov, Internet: SciTecLibrary.com. (http://sciteclibrary.ru/eng/catalog/pages/6727.html)
БИОЛОГИЧЕСКОЕ И ЛЕЧЕБНОЕ ДЕЙСТВИЕ УЛЬТРАЗВУКА. Глава 12, “Ультразвук и его применение в науке и технике”, Л. Бергман. “Иностранная Литература”, (1957) Москва, стр. 545 – 573.
Для подготовки данной работы были использованы материалы с сайта http://www.sciteclibrary.ru
Похожие работы
... ряде случаев после полной химической очистки воды ее УФ-спектр также сохраняет определенные изменения (главным образом, повышенное УФ-поглощение по сравнению с чистой водой). Известно, что ультрафиолетовая часть спектра является управляющим элементом для живых систем. Его отсутствие (поглощение) ведёт к нарушению функций управления. В связи с этим, возникает необходимость пересмотра существующих ...
0 комментариев