А.В.Конарев, В.Г.Конарев, Н.К.Губарева, Т.И.Пенева.
Обсуждаются основные направления и перспективы использования белковых и ДНК-маркеров в решении прикладных и теоретических проблем генетических ресурсов растений, селекции, сортоиспытания, семеноводства и семенного контроля. Рассмотрены конкретные примеры практического использования белковых и ДНК-маркеров для решения важнейших проблем прикладной ботаники, генетики и селекции.
Мировые генетические ресурсы растений рассматриваются во всем мире как основной источник улучшения сельскохозяйственных культур на ближайшие десятилетия. Создание источников и доноров селекционно важных признаков, т.е. организация предселекционной работы, в большинстве случаев базируется на мировых генетических ресурсах или коллекциях культивируемых растений и их диких сородичей. Раскрытие потенциала генетических ресурсов по основным биологическим и селекционным признакам обеспечивает генетическую базу для реализации селекционных программ различных направлений. В целом же предселекционная работа включает все этапы работы с генофондом от сбора, поддержания и изучения до правовых аспектов авторства на доноры и источники ценных признаков. Чтобы служить эффективной базой для улучшения культур, сохраняемое в центрах генетических ресурсов растений (ГРР) и генных банках генетическое разнообразие должно быть не только тщательно и всесторонне изучено. Сами коллекции должны быть рационально организованы. Строго говоря, каждый образец коллекции должен быть идентифицирован и паспортизирован. Сохранение генетической целостности образцов также входит в разряд принципиальных проблем. Речь идет о сохранении не только образца как такового, но с его ценными свойствами, в частности адаптивными и др. признаками.
Во Всероссийском НИИ растениеводства им. Н.И.Вавилова (ВИР) познание генетического разнообразия культурных растений и их дикорастущих сородичей традиционно осуществляется с использованием комплекса методических подходов на базе тесного взаимодействия кураторов культур и специалистов методических отделов [1,2]. Принципиальное преимущество такой системы оценки исходного материала для селекции в возможности достаточно глубокого и всестороннего изучения максимального разнообразия культуры. В эффективности познания генофонда решающая роль принадлежит методам исследования.
С момента своего основания ВИР являлся методологическим и методическим центром в области фундаментальных и прикладных проблем растениеводства. Эта роль вытекает из его структуры и основных задач. Наличие мирового генофонда по большинству ведущих культур создает благоприятные условия для развития методологий, подходов и, как следствие, методов исследований генофонда - исходного и селекционного материала. Практически это было реализовано в развитии методов генетики, иммунологии, физиологии, биохимии, технологической оценки применительно к проблемам исходного и селекционного материала[1]. Именно по такой логике закладывались и развивались в ВИРе молекулярно-биологические исследования. В основу всех методических подходов также положен охват всего доступного генетического разнообразия культуры, включая дикорастущих сородичей [3,4].
Развиваемые в ВИРе молекулярно-биологические исследования ориентированы на решение теоретических и прикладных проблем интродукции, изучения, хранения, воспроизведения, идентификации и регистрации и паспортизации генетических ресурсов растений. Особое внимание уделяется разработке эффективных методов для использования в сортоиспытании, селекции, семеноводстве и семенном контроле (Табл. 1).
Таблица 1.
Использование белковых и ДНК-маркеров (молекулярных маркеров - ММ) на различных этапах работы с генетическими ресурсами растений (ГРР) и с селекционным материалом
I. Интродукция ГРР.
Поиск с использованием ММ нового разнообразия для привлечения в коллекцию. Использование ММ для контроля процесса включения в коллекцию нового образца (для предотвращения дублирования).
II. Структура коллекции
Выяснение с использованием ММ внутривидовых связей и межвидовых отношений:
а) использование полиморфизма белков и ДНК для анализа внутривидовых связей;
б) анализ родства видов и геномов с использованием белковых и ДНК-маркеров.
III. Технологии создания коллекций
Использование ММ для идентификации и регистрации ГР важнейших культур, создания каталогов формул и баз данных по молекулярным маркерам, для выявления ошибок в определении образцов коллекции, для поиска дублетных образцов, при формировании стержневых коллекций, для контроля стабильности форм при создании коллекций in vitro.
IV. Сохранение ГРР ex situ
Использование ММ для контроля за генетической целостностью образцов при их репродукции и разных способах хранения.
V. Охрана авторских прав ВИР на источники, доноры, формы из генетических коллекций Идентификация и регистрация с использованием ММ источников и доноров ценных признаков, образцов генетических коллекций.
Использование ММ в решении спорных вопросов авторства - участия того или иного исходного материала в создании новых форм растений.
VI. Использование молекулярных маркеров в селекции Поиск и отбор ценных генотипов по белковому фенотипу. Контроль за включением желаемых генотипов в процессе селекции. Контроль за динамикой популяционного (генотипного состава).
Определение гибридности семян, предсказание скрещиваемости, предсказание степени гетерозиса, прочие задачи селекции.
VII. Использование ММ в сортоиспытании
Регистрация и документация районированных и снятых с районирования сортов в виде белковых формул; создание баз данных сортов по белковым формулам. Определение происхождения и оригинальности сорта. Определение однородности и стабильности сорта (генетическая целостность сорта).
VIII. Использование ММ в семеноводстве и семенном контроле Проверка типичности при отборе лучших растений в первичном семеноводстве. Выяснение природы нетипичных растений.
Контроль за спонтанным переопылением или засорением.
Контроль за генетической целостностью (за генотипным составом в процессе семеноводства, процентом гибридности и т.п.).
Молекулярное маркирование основано на полиморфизме, найденном в белках и нуклеиновых кислотах. Перед тем, как маркер может использоваться для решения каких-либо конкретных задач, корректность его использования должна быть аргументирована с генетических и биохимических позиций [4-6]. Солидное генетическое обоснование использованию проламинов как генетических маркеров было дано в работах академика А.А.Созинова и его школы (7-9). Как нет идеальных молекулярных маркеров, так и последние не являются инструментами, использование которых в изоляции от других подходов, обеспечат успех в решении той или иной проблемы.
В работе с ГРР, а равно и в решении проблем селекции и семеноводства, где анализируются и сравниваются множества объектов, одним из важных аспектов в оценке эффективности ММ и маркерных техник является воспроизводимость результатов и повторяемость опытов, т.е. возможность стандартизации метода. Так многие изоферментные системы обладают крайне нежелательной для маркеров онтогенетической, тканевой либо органной специфичностью, а также изменчивостью в зависимости от температуры и кислотности среды, режима питания и т.п. Это сильно снижает значимость изоферментов как генетических маркеров, особенно если ставится задача разработки стандартных и арбитражных методов [2,4].
Иначе обстоит дело с запасными белками семян. Принципиально важно, что семя - фиксированная фаза онтогенеза. Запасные белки семян остаются неизменными часто в течение многих лет. Воспроизводимость результатов и повторяемость опытов по электрофорезу запасных белков хорошие [6,7,10,11]. С использованием запасных белков семян в качестве маркеров связаны реальные практические достижения в идентификации и регистрации сортов важнейших сельскохозяйственных культур, в семеноводстве и семенном контроле, что закреплено в решениях такой авторитетной международной организации как ISTA и ряда других [12-14].
Использование ДНК-маркерных технологий привлекает исследователя прежде всего возможностью работать с самим носителем наследственной информации. Однако для наиболее простых в исполнении методов (RAPD) характерны плохая воспроизводимость и доминантный характер маркера (невозможность различения гомо- и гетерозиготных генотипов). Более совершенные методы трудоемки и дороги в исполнении. Подробнее об этом [2,15,16]. Заключение о недостаточной готовности ДНК-маркерных технологий для широкого использования в сортовой идентификации и семенном контроле сделано специальной Рабочей Группой ISTA (ISTA NB. No 113, 1997). Практически к аналогичным выводам, но только в связи с решением широкого круга проблем генетических ресурсов растений (особенно практических проблем генных банков) пришли участники рабочего совещания, организованного Международным Институтом Генетических Ресурсов Растений (IPGRI) [16 ].
В трудах этого совещания [17], а также в техническом бюллетене IPGRI [18], многих других работах зарубежных исследователей последних лет справедливо отмечается, что разные ДНК-маркерные системы (в первую очередь, наиболее доступные такие как RFLP, RAPD, AFLP) достаточно широко и эффективно используются для выяснения степени родства (или генетических связей) на внутривидовом и межвидовом уровнях. Не смотря, как было сказано выше, на достаточно пессимистическую оценку перспектив и возможностей ДНК-маркерных систем для решения большинства прикладных проблем генетических ресурсов растений, полностью игнорируются реальные достижения и практическое широкое и многолетнее использование в этих целях полиморфизма запасных белков (табл.2) [2-15,19]. Так, например, все проблемы идентификации и регистрации сортов (и генетических ресурсов растений в целом), практические проблемы рациональной организации коллекций планируется решать исключительно с использованием ДНК-маркеров [16-18]. При этом не приводится ни одной разработанной системы идентификации и регистрации генетических ресурсов культуры или группы культур, ни одного примера реального практического использования ДНК-маркерных систем в сортоиспытании или семенном контроле. Создается впечатление, что сторонники исключительного использования ДНК-маркеров в решении проблем генетических ресурсов растений, семеноводства и семенного контроля по какой-то причине совершенно не знакомы с большим объемом информации (мировой литературой), касающейся практического использования белков в качестве маркеров, не имели дело с реальным генетическим разнообразием вида или культуры, а также с практическими проблемами идентификации сортов, семеноводства и семенного контроля.
Таблица 2.
Результаты деятельности ВИР им. Н.И.Вавилова по изучению и регистрации генетических ресурсов растений с использованием белков семян в качестве маркеров
| Название рода | Число изученных н зарегистрированных: | Издано: | ||
| Видов | Сортов н дикорастущих популяции | Каталогов белковых формул | Метод, указании | |
| Пшеница | 20 | 4300 | 12 | 4 |
| Ячмень | 17 | 255 | 1 | 3 |
| Роясь | 7 | 180 | 1 | |
| Овес Тритикале | 19 1 | 215 500 | 1 | 3 |
| Этнлопс | 25 | 2080 | 4 | |
| Кукуруза | 1 | 410 | 2 | 1 |
| Рнс | 17 | 1776 | ||
| Сорю | 28 | 155 | ||
| Пыреи | 40 | 120 | ||
| Элнмус | 33 | 68 | ||
| Житняк | 4 | 25 | ||
| Колосник | 8 | 17 | ||
| Овсяница | 50 | 26 | 1 | |
| Плевел | 9 | 168 | 1 | |
| Ежа Мятлик Другие злаковые | 3 30 131 | 173 120 1260 | 1 | |
| Фасоль Другие бобовые Картофель | 88 118 44 | 102 510 300 | 1 | |
| Свекла | 13 | 300 | 2 | |
| Капуста Лук Амарант | 20 27 5 | 209 18 | 105 | 1 |
| Подсолнечник Лен | 30 6 | 700 40 | 2 | 2 |
| Гречиха | 4 | 150 | ||
| Плодовые Ягодные Цитрусовые | 33 21 13 | 333 160 47 | ||
| Куфня | 26 | 36 | ||
| Хохоба | 1 | 50 | ||
| Цитрусовые | 13 | 47 | ||
Распространенной методической ошибкой, встречающейся в работах зарубежных и отечественных исследователей является смешение понятий идентификация и дифференциация или различение сортов, генотипов и т.п. Исследователи, предлагающие новые методы идентификации сортов, образцов, гибридов и т.п., часто вообще не вдаются в подробности требований, предъявляемых к такого рода методам в государственных или иных структурах, имеющих дело с идентификацией и регистрацией сортов, семенным контролем. Идентифицировать сорт (генотип, гибрид, линию, клон) в том числе значит гарантированно узнать его с использованием данного метода в любой ситуации. Для этого надо иметь каталоги и базы данных, охватывающие генетическое разнообразие культуры или в целом вида (см. ниже). Надежный метод записи спектров ДНК и белков (номенклатура компонентов спектра ММ) - принципиальный элемент любой стандартной системы идентификации и регистрации сортов по ММ [3,4,7-13]. Только в этом случае возможен обмен данными между контролирующими лабораториями независимо от их принадлежности (генные банки, коммерческие и государственные контрольно-семенные лаборатории и т.п.)[14].
Во ВННИР им. Н.И.Вавилова еще в начале 70-х годов В.Г.Конаревым, И.П.Гаврилюк и Н.К.Губаревой была разработана номенклатура компонентов электрофоретических спектров глиадина [3-6]. Принципиальное отличие предложенной номенклатуры состоит в том, что она базируется на результатах обстоятельного изучения внутривидовой изменчивости маркерного белка в мировой коллекции. В дальнейшем эта идея была реализована для запасных белков семян большинства важнейших культур (Табл.2). Используя эталонный спектр, составленный для культуры можно записать спектр любого сорта, биотипа, образца в виде так называемых белковых формул [4,14]. В первую очередь это было осуществлено для большого числа родов злаков, включающих важнейшие зерновые и кормовые культуры (табл.2). Был составлен единый эталонный спектр пролами-нов злаков триб пшеницевых (Triticeae Dum.), овсовых (Aveneae Dum.), тимофеевковых (Phleeae Dum.) и мятликовых (Роеае R. Br.). Этот спектр был основан на изучении спектров пролами-нов десятков тысяч отдельных семян (генотипов), включающих все возможные позиции этого белка. Кроме того, для каждой культуры был составлен рабочий эталонный спектр проламина [4,19].
Полиморфизм запасных белков семян бобовых - глобулинов был использован в сортовой идентификации зерновых бобовых культур - гороха, сои, вики, люпина, кормовых бобов, а также кормовых бобовых трав (клевер, люцерна, донник). У бобовых как и у ряда других двудольных растений для целей идентификации были привлечены 7S (вицилиноподобные) и 11S (легумино-подобные) глобулины [4,19]. В настоящее время во ВНИИР им. Н.И.Вавилова сортовая идентификация по спектрам запасных белков (глобулинов) осуществляется у многих других двудольных растений, в частности подсолнечника, свеклы, капусты, салатных растений, рапса, горчицы, гречихи и многих других (табл.2) [4,19]. Так по результатам сравнительного анализа 150 представителей различных видов капустных (Brassica L.) был составлен эталонный спектр круциферина (глобулина) для представителей данного рода, включающего важные кормовые, овощные и технические культуры. По такому спектру записываются формулы круциферина видов, сортов, линий, гибридов представителей этого рода [20].
Аналогичным образом был составлен суммарный (эталонный) спектр гелиантинина (глобулина подсолнечника). Спектр состоит из всех возможных позиций полипептидов этого белка, обнаруженных к настоящему времени при анализе внутривидовой изменчивости [4,19], что позволяет идентифицировать и регистрировать все внутривидовое разнообразие подсолнечника, включая сорта, линии, гибриды [21]. Номенклатуры электрофоретических спектров глиадина были разработаны также специалистами Франции, Канады и ряда других стран [10-12]. Генетическая номенклатура спектров и компонентов проламинов пшеницы и ячменя была предложена и успешно развита в работах А.А.Созинова и его учеников [7-9].
Во ВНИИР им. Н.И.Вавилова генофонд сортов и дикорастущих образцов документируется в виде белковых формул. Полученная информация сохраняется в виде каталогов формул (табл.2) и компьютерных баз данных формул. Для надежной идентификации и регистрации сортового генофонда ведущих зерновых культур и их дикорастущих сородичей в большинстве случаев достаточно электрофореза запасных белков. Иногда приходится использовать другие типы белков, например глютенины или ингибиторы протеолитических ферментов [4, 6, 19].
Принципиальной является проблема соответствия понятий статуса сорта в общепринятом смысле и с использованием молекулярных маркеров. Для корректного подхода к вопросу предварительно проводятся специальные исследования на большом объеме сортов и на внутривидовом разнообразии диких сородичей. Наиболее сложно дело обстоит с перекрестноопыляющимися культурами, поскольку каждый сорт или образец представляет собой сложную смесь различных генотипов, которым соответствуют разные типы спектра белка или ДНК. Такая сортовая или дикорастущая популяция может быть идентифицирована и зарегистрирована по наличию определенных типов спектра и частоте их встречаемости [4,19]. После сформирования баз данных (компьютерных либо каталожных) открываются реальные возможности использования белковых или ДНК-маркеров для решения ряда практических вопросов коллекций, например, некоторых проблем интродукции или пополнения коллекции (табл.1). На основании сравнительного анализа информации заложенной в каталоги или в базы данных в виде белковых формул, а также информации, полученной при анализе вновь поступившего семенного материала, может быть сделан предварительный вывод о степени оригинальности последнего (с целью предотвращения дублирования образцов). Многолетний опыт ВИР показывает, что такая информация, как правило, в дальнейшем подтверждается так называемыми традиционными методами [2].
Для оценки степени генетических различий между образцами, например, образцами разного географического происхождения, или культурными и дикими формами широко используются ДНК- и белковые маркеры. Во ВНИИР им. Н.И.Вавилова мы использовали ПДРФ-маркеры (RFLP) в изучении генетической дифференциации ячменя. Однако, для анализа обширных коллекций метод ПДРФ достаточно трудоемок. Здесь удобнее использовать более простой и достаточно чувствительный RAPD-анализ. Такие работы, в частности, были проведены ВИРом совместно с Национальным Институтом Сельскохозяйственных Исследований (Япония) на образцах дикого и культурного ячменей из коллекции ВИР, а также на ряде других коллекций [22]. Целью работы была оценка взаимоотношений между различными формами культурного и дикого ячменей по полиморфизму фрагментов ДНК, амплифицированных в полимеразной цепной реакции с произвольными праймерами (RAPD). Полученные результаты позволили разделить изученные сорта и местные популяции на три основные группы, которые, очевидно, отражают основные тенденции в эволюции и географическом распространении культурного ячменя. Обнаружено соответствие выделившихся групп и кластеров культурного ячменя мировым центрам разнообразия культурных растений (генцентрам), выявленным Н.И.Вавиловым, а также современной эколого-географичес-кой классификации ячменя [22].
Аналогичный экспериментальный подход был применен для выяснения структуры коллекций гексаплоидных пшениц. Методом RAPD-анализа изучена степень родства между образцами гексаплоидных пшениц разных эколого-географических групп (более 400 образцов из коллекции ВИР). Исследования проводились также в том числе и в связи с возможностью маркирования генотипов с высоким уровнем зимостойкости, а также в связи с отработкой технологии создания стержневых коллекций. Результаты координатного и кластерного анализа этих данных продемонстрировали, что все изученные образцы гексаплоидных пшениц представляют собой единый генный пул, который делится на четыре большие группы. Обнаруженное деление находится в полном соответствии с эколого-географической классификацией предложенной Н.И.Вавиловым [23]. Удалось идентифицировать праймеры, позволяющие выделять группы образцов с ценными адаптивными свойствами (например, холодостойкость). Это свидетельствует о перспективности данного подхода к анализу мировой коллекции как исходного материала для селекции по важнейшим признакам. Подобные исследования с использованием ДНК-маркеров проведены недавно сотрудниками ВИР на коллекциях овса, проса, вики, риса. Использованию белковых маркеров для решения вопросов внутривидовых связей посвящено большое число публикаций, в том числе сотрудников ВИР [4,7,10,11,19,24].
Анализ межвидового (междугеномного) родства имеет значение не только для решения вопросов филогении и систематики, но, главным образом, для селекции, базирующейся на методах отдаленной гибридизации. Молекулярно-биологические методы (ДНК-гибридизация, имму-нохимические методы, электрофорез белков) были на первом этапе применены для анализа родства видов и геномов, ревизии схем филогении и происхождения геномов, созданных на базе классических, в том числе цитогенетических методов. В ВИРе применение эффективных имму-нохимических подходов и методов для геномного анализа пшениц и эгилопсов привело в 70-е годы к принципиально новому решению вопроса происхождения первых геномов мягкой и твердой пшениц [4,19,25], что позволило отделу пшениц ВИР создать новую систему рода Triticum L.[26]. Два года спустя наши данные о происхождении первого генома мягкой и твердой пшениц от диплоидного вида близкого к современной однозернянке T.urartu Thum. были подтверждены британскими цитогенетиками [27] и лишь 14 лет спустя в 1988 году и американскими генетиками и молекулярными биологами с использованием ДНК-геномных маркеров [28]. Анализ межвидового и геномного родства был осуществлен в ВИРе с использованием геномноспецифичных белковых маркеров у многих культурных растений и их диких сородичей [4,6,19]. Для анализа межвидовых (межгеномных) отношений, решения вопросов филогении в ВИРе используются также RAPD- и RFLP-маркеры [29,30].
Для нормального существования коллекций, а также для повышения эффективности их использования в селекционном процессе и исследовательской работе важную роль играет соблюдение определенных норм и принципов формирования таких коллекций.
Так при поиске ошибок в определении образцов коллекции в ряде случаев очень полезны могут быть белковые либо ДНК-маркеры. Характерным является пример работы с коллекцией вида T.persicum Vav., проведенной еще в 70-е годы. Представители этого тетраплоидного вида пшеницы морфологически очень трудно отличимы от так называемой «персикоидной» формы гексаплоидной пшеницы. После изучения белков семян коллекции T.persicum методом электрофореза выяснилось, что из 116 образцов 38 оказались гексаплоидами типа «персикоидес» [2]. Конечно, можно было изучить всю коллекцию кариологическим методом, но это достаточно трудоемкий процесс.
Диплоидные пшеницы T.urartu и T.boeoticum отличить по морфологии в состоянии только узкие специалисты. С помощью электрофоретических и, особенно, серологических маркеров сделать это не представляет труда [4,19]. До опубликования наших работ в 1974 году[25], многие зарубежные генетики и тритикологи вообще не признавали пшеницу Урарту за самостоятельный вид. Сохранение ГРР ex situ. Задача ex situ консервации есть поддержание образцов без изменения их генетической конституции [17]. Необходимо свести к минимуму возможность изменений, происходящих с образцами посредством мутаций, селекции, случайного дрейфа или засорения. Это одно из важнейших условий полноценного функционирования генных банков любых организмов. Контроль за генетической целостностью - подлинностью, чистотой хранимого (и периодически пересеваемого) материала - важнейшая задача генного банка. Проблема идентификации в мировых коллекциях дублетов и так называемых очень сходных образцов остро встала в генных банках, особенно с крупными коллекциями. В решении всех этих задач молекулярные технологии оказались очень полезными [2,4, 17,19, 31-33]. Так в ВИРе накоплен большой опыт по использованию спектров запасных белков семян для анализа динамики популяций перекрестноопыляющихся культур (и генотипического состава самоопыляющихся) в процессе репродукции образцов. В коллекции ВИР сохраняется большое число образцов старых русских местных сортов мягкой озимой пшеницы и других местных сортов и форм. Было показано, что старые сорта, сорта народной селекции имеют более высокий уровень популяционного полиморфизма по сравнению с современными сортами, что делает их ценным источником генетического разнообразия для улучшения современных сортов [4,19,34,35]. Задача генных банков и центров генетических ресурсов растений не только собрать, зарегистрировать и изучить эти образцы, но и сохранить все богатство генетической изменчивости этих уникальных форм.
Специальные исследования были проведены в ВИРе на 6 образцах стародавних мягких пшениц - Банатках. В течение трех лет (1988-89, 89-90,90-91) образцы высевались на Кубанской ОС ВИР. Контроль за составом генотипов осуществлялся по спектрам глиадинов. Было показано, что у образца к-4816 в течение трех лет концентрация доминирующего генотипа «а» снизилась с 78% до 52%, а генотипа «в» повысилась с 6 до 33%. У образца к-10230 концентрация доминирующего генотипа «а» снизилась с 84% до 57%, а генотипа «б» повысилась с 7% до 31%. Таким образом, через три года репродукции генотипный состав образцов несколько изменился [35,36].
Генетические коллекции - особо ценный материал, сохранность которого сопряжена с многими методическими трудностями. Белковые маркеры используются в ВИРе для контроля за стабильностью генотипов растений, представляющих генетические линии, для идентификации чужеродных транслокаций, мутаций, изменения числа и состава хромосом. Так сравнительный анализ спектров глиадинов соротов-оригиналов мягкой пшеницы с транслокациями и замещениями пшеничной хромосомы на ржаную (1B/1R) и их репродукций показал, что что у ряда репродуцированных образцов встречаются генотипы с отсутствием глиадиновых маркеров короткого плеча хромосомы 1RS ржи. Эти и многие другие примеры указывают на необходимость контроля чистоты и целостности коллекций в процессе их репродукции.
Любой хранитель (собственник) генетических ресурсов заинтересован обеспечить как охрану своих авторских прав на исходный материал, источники и доноры, так и официальное признание участия этого генетического материала в создании тех или иных сортов в своей стране и за рубежом. ММ оказывают здесь реальную помощь, обеспечивая независимую информацию о происхождении и степени родства генетического материала сравниваемых образцов. В ВИРе накоплен большой опыт использования белковых маркеров в решении спорных вопросов авторства сортов, их оригинальности, подлинности, чистоты, природы исходного материала и т.д., когда использование набора традиционных методов, в том числе и грунт контроля, не давало желаемого результата [4,19]. Эффективность такой работы во многом обеспечивается наличием в ВИР каталогов белковых формул и баз данных этих формул с информацией о сотнях сортов, в том числе давно снятых с районирования, а также о многих образцах различного происхождения, хранящихся в коллекции (табл.2). Важно подчеркнуть, что даже не являясь тестом, который может быть официально признан как единственный аргумент, молекулярный метод обеспечивает предварительную, независимую и оперативную информацию.
Использование молекулярных маркеров в селекции. Белковые маркеры на протяжении последних десятилетий используются в селекционных программах для решения многих вопросов (табл. 1). Этому посвящено большое число отечественных и зарубежных публикаций [6,7]. В ВИРе проламиновые спектры, в частности, используются для отбора определенных генотипов (по соответствующим типам спектра) при селекции различных культур. Так в ходе селекции сорта озимой пшеницы «Тюменская ранняя» с помощью спектров глиадина формировался «желаемый» генотипный состав создаваемого сорта (НИИСХ Северного Зауралья).
Весьма наглядным является пример связи между белковой формулой генотипов у сортов озимой мягкой пшеницы и устойчивостью этих генотипов (сортов, имеющих данные генотипы) к низким температурам. Одним из основных свойств, которым должна обладать озимая мягкая пшеница является зимостойкость. Из характеристик, обуславливающих зимостойкость, наиболее изучена морозостойкость. Последняя контролируется многими генами, локализованными в разных хромосомах. Это, естественно, затрудняет исследование признака зимостойкости, его маркирование и соответствующую селекцию. По данным А.А.Созинова и сотрудников [7] сорта озимой пшеницы, в составе спектров глиадина которых присутствуют определенные блоки компонентов, обладают повышенной зимостойкостью. Согласно биохимической номенклатуре компонентов глиадина, разработанной в ВИРе это соответствует компонентам: г2щ78 (блок Gld 1A1), г1щ67 (блок Gld 1A2), г13щ5819 10 (блок Gld 1D5), 62467в1 (блок Gld 6A3) и 657в245 (блок Gld 6D2). Исследования были проведены на большом числе сортов озимых мягких пшениц (около 300) разных экологических групп (групп селекции) [36]. Морозостойкость растений определялась методом прямого промораживания в посевных ящиках. Дифференциирующими температурами были -15С и -18С. Действительно, большинство сортов с высокой и повышенной морозостойкостью характеризуется указанными выше блоками (группами) компонентов глиадина. Так на большом числе сортов показано, что наличие генотипа с компонентами 62467 и щБ^ 10 придает сорту повышенную морозостойкость (табл. 4) [36]. Дело, однако, усложняется в ходе анализа генотип-ного состава сортов. Как правило, не удается обеспечить 100% концентрацию выдающихся генотипов по одному признаку и селекционеры вынуждены «разбавлять» сорт другими генотипами, обеспечивающими другие характеристики, но, обладающими меньшей зимостойкостью. Особенно хорошо это заметно при анализе родословных некоторых наших отечественных сортов озимой мягкой пшеницы. В родословной озимых мягких пшениц одесской селекции на первом этапе присутствует морозостойкий сорт Гостианум 237 (группа морозостойкости 1). Выяснено, что если после последующих скрещиваний полученные сорта по проламиновым спектрам были близки к Гостианум 237 (в частности, по наличию и частоте встречаемости генотипов с компонентами 62467 и компонентами 8Х9 10 в щ-зоне), они также обладали хорошей морозостойкостью. Таким образом, спектры глиадина можно использовать для определения потенциальной морозостойкости сортов озимой мягкой пшницы в пределах определенных групп селекции. При этом необходимо знать белковую формулу генотипа (генотипов) морозостойкого сорта, который явился источником данного признака в ходе селекции. Это позволит вести контроль за включением его генетического материала во вновь создаваемые сорта.
В данном случае казалось бы речь идет об использовании простой маркерной системы для маркирования полигенного признака (морозостойкость). Но фактически маркируется не признак, а то что называется интегральным состоянием генома. Маркируется генотип вместе с его адаптивным генным комплексом. Поэтому упомянутый специфический спектр не является в строгом смысле маркером морозостойкости (хотя нельзя отрицать и адаптивного характера полиморфизма проламинов). Трактовать такого рода данные надо очень осторожно и применительно к конкретным маркерам и обстоятельствам.
В Вире за последние 20 лет проведены широкомасштабные исследования генофонда культурной и дикорастущей ржи по полиморфизму секалина. Наряду с идентификацией и паспортизацией мирового генофонда значительное внимание было уделено проблемам селекции. Фактически разработаны подходы к селекции ржи, сопровождаемой молекулярными маркерами [2,19,33].
Примером использования полиморфизма проламинов для сопровождения селекционного процесса является история создания сорта ржи Ильмень. Использование в селекции короткостебельных сортов донора короткостебельности ЕМ 1 для скрещивания с обычными высокорослыми сортами обычно завершается выделением и отбором короткостебельных растений. Этот процесс приводит к уменьшению уровня популяционного полиморфизма, что выражается в соответствующих характеристиках молекулярного полиморфизма [33]. Практическим следствием является снижение продуктивности, уровня адаптивности (снижается устойчивость к неблагоприятным факторам), ухудшаются другие хозяйственные признаки. Причины данных негативных явлений были объяснены на основе данных молекулярного анализа популяционного полиморфизма семейства сортов Малыш. На этом этапе селекции не удалось достигнуть оптимального уровня генетического полиморфизма, что отразилось на адаптивных свойствах популяции, а также на продуктивности сорта.
Оптимальный уровень полиморфизма был достигнут фактически менее жестким отбором после повторного скрещивания с Вяткой-2 (по данным молекулярного полиморфизма обогащение популяции проходило за счет редких генотипов). По мере возрастания внутрипопуляционно-го разнообразия увеличивалась и продуктивность у последующих вариантов сорта - Малыш 77-79 гг и особенно далее у сортов Россиянка и Ильмень.
Таким образом показано, что анализируя скрытую генетическую изменчивость можно не только контролировать состав популяций в ходе искусственного отбора, семеноводства, влияния среды и т.д., но и основываясь на основных положениях популяционной генетики вскрывать причины тех или иных явлений, ставить диагноз, указывать способы решения и предсказывать практически важные селекционные последствия для разных культур.
Белковые маркеры с успехом используются для определения гибридности семян (сортов) злаковых трав [2,19,24], свеклы [37], различных видов капусты и горчицы [20,38], подсолнечника [2,19,21], кукурузы [2,19,39] и многих других культур, а также для прогнозирования эффекта гетерозиса сорго по продуктивности [40,41].
Использование белковых маркеров в сортоиспытании. С конца 70-х годов белковые маркеры наряду с полевыми и лабораторными методами используются в системе Государственной комиссии по сортоиспытанию сельскохозяйственных культур при министерстве сельского хозяйства СССР для установления оригинальности, однородности и константности сортов пшеницы, ячменя и овса [31,42]. Следует отметить, что одновременно с методикой электрофореза белков в полиакриламидном геле, разработанной в ВИРе, для идентификации сортов пшениц применялась методика электрофореза в крахмальном геле, предложенная А.А.Созиновым и его учениками [7,8,42]. Формулы проламинов, записанные на основе анализа семян соответствующего сорта, полученного от учреждения оригинатора, были включены в Государственный реестр сортов (введен с 1993 года), допущенных к использованию в производстве [43].
С середины 80-х годов электрофорез проламинов привлекался в ряде случаев Государственной комиссией по сортоиспытанию для решения отдельных спорных вопросов, касающихся оригинальности, однородности и константности новых сортов кормовых злаковых трав - ежи сборной (Dactylis glomerata L.), райграса (Lolium L.), овсяницы луговой (Festuca pratensis L.). Метод использовался также для определения гибридной (амфидиплоидной) природы семян, при решении вопроса о соотношении гибридных и негибридных зерновок в сортовой популяции.
Использование в семеноводстве и семенном контроле. Белковые маркеры наряду с традиционной схемой (полевая апробация и др.) успешно используются для определения сортовой чистоты при семеноводстве и семенном контроле. Метод позволяет вести отбор элитных растений и потомств. Особенно эффективны белковые маркеры в последующих звеньях первичного семеноводства - питомниках испытания потомств и питомниках размножения, где обязателен контроль за появлением нетипичных для сорта растений. Нетипичные формы растений могут быть выявлены путем позернового анализа. В ряде случаев при принятой схеме элитного семеноводства может нарушаться биотипный (генотипный) состав сорта, а значит, могут измениться его характеристики. Контроль посредством электрофореза белков отдельных семян очень эффективен для решения этих практических вопросов и может резко поднять качество семеноводства и сократить сроки создания элиты с 7 до 5 лет, что продемонстрировано на многих примерах [4,19]. Наиболее наглядно это продемонстрировано на примере первичного семеноводства сорта ячменя Криничный [44]. Использование электрофореза белков в семеноводстве и семенном контроле и, в частности, для конкретных культур и групп культур описано в соответствующих методических указаниях и рекомендациях (табл.2) [20,21,37,39,44,45]. Детально эти вопросы изложены в специальной работе В.Г.Конарева[46]. Здесь же приведен полный перечень названий каталогов белковых формул, методических указаний и рекомендаций по использованию белковых маркеров, изданных в ВИРе с 1970 года (см. также табл. 2).
Список литературы
1. Конарев А.В. Всероссийский НИИ растениеводства и его вклад в развитие сельскохозяйственной науки и селекции страны. Сельскохозяйственная биология. 1994, 3: 13-75.
2. Конарев А.В. Использование молекулярных маркеров в работе с генетическими ресурсами растений. Сельскохозяйственная биология, 1998, 5: 3-25.
3. Конарев В.Г. Принцип белковых маркеров в геномном анализе и сортовой идентификации пшеницы
4. Молекулярно-биологические аспекты прикладной ботаники, генетики и селекции. Теоретические основы селекции.Том 1.(под ред. В.Г.Конарева) М., Колос, 1993, 447с.
5. Конарев В.Г. Принцип белковых маркеров в генетическом анализе исходного и селекционного материала. Физиология растений в помощь селекции. М., 1974 : 242-269
6. Конарев В.Г. Белки растений как генетические маркеры. М.: Колос, 1983, 320с.
7. Созинов А.А. Полиморфизм белков и его значение в генетике и селекции. М.: Наука, 1985.
8. Созинов А.А., Попереля Ф.А. Полиморфизм глиадина и возможности его использования. Растительные белки и их биосинтез. М.: Наука, 1975: 65-76
9. Созинов А.А.,Попереля Ф.А., Стаханова А.И. Использование электрофореза глиадинов в селекции на качество. Вестник с.-х. науки, 1974, 7: 99-108.
10. Autran J.C. and Bourdet A. L'identification des varietes de ble: etablissement d'un tableau general de determination fonde sur le diagramme electrophoretique des gliadines du grain. Ann. Amelior. Plantes, 1975,25, 3:227-301.
11. Bushuk W. and Zillman R. Wheat cultivar identification by gliadin electrophoregrams . I. Apparatus, method and nomenclature. Can. J. Plant Sci., 1978, 58: 505-515.
12. Cooke R.J. The standartizaton of electrophoresis methods for variety identification. In: Biochemical Identification of varieties (Materials III International Symposium ISTA, Leningrad, USSR, 1978), VIR, Leningrad, USSR, 1988:14-27.
13. Cooke R.J. Modern methods for cultivar verification and the transgenic plant challenge. Abstracts of 25th International Seed Testing Congress (Pretoria, April 15-24, 1998), ISTA, Zurich, 1998: 9-10.
14. International Rules for Seed Testing. Rules 1996. Verification of species and cultivar. Seed Sci.& Technol, 1996, 24 (Supplement): 253-270.
15. Конарев В.Г. Морфогенез и молекулярно-биологический анализ растений. Спб., ВИР, 1998: 370с.
16. Karp A. and Edwards К Molecular techniques in the analysis of the extent and distribution of genetic diversity. Molecular genetic techniques for plant genetic resources. Report of an IPGRI Workshop, October 1995, Rome, Italy, 1997, 11-22.
17. Molecular genetic techniques for plant genetic resources. Report of an IPGRI Workshop 9-11 October 1995 Rome, Italy. Editors: Ayard W.G., Hodgkin Т., Jaradat A., and Rao V.R. IPGRI, 1997:137p.
18. Karp.A, Kresovich S, Bhat К V. and Hodgkin T. Molecular tools in plant genetic resources conservation: a guide to the thechnologies. IPGRI technical bulletin, 1997,2: 47.
19. Molecular biological aspects of applied botany, genetics and plant breeding. Theoretical basis of plant breeding. Vol. I. St.-Petersburg, VIR, 1996: 228 p.
20. Идентификация, регистрация и оценка чистоты сортов, линий и гибридов капусты методами электрофоретического анализа изоферментов и запасных белков. Методические указания (под ред. И.П.Гаврилюк). Ленинград, ВИР, 1991 : 26с.
21. Идентификация, анализ и регистрация сортов, линий и гибридов подсолнечника методом электрофореза гелиантинина. Методические указания ( под ред. И.П.Гаврилюк). Ленинград, ВИР, 1988:23с.
22. Стрельченко П.П., Ковалева О.К, Окуно К. Оценка биологического разнообразия культурного и дикого ячменей с использованием полиморфизма амплифицированных фрагментов ДНК. Проблемы ботаники на рубеже XX-XXI веков. Тезисы докл.П(Х) съезда Русского бот. Общ-ва.. СПб, 1998, 1:380-381.
23. Strelchenko P.P., Mitrofanova O.P., Konarev A. V. and Terami F. RAPD characterization of relationships among cultivated hexaploid wheats. Abstracts submitted to XVI International Botanical Congress, August 1-7,1999, St.Louis, MO, USA.
24. Konarev A.V, Vvedenskaya I.O., Nasonova E.A. and Perchuk I.N. Use of prolamine polymorphism in studing genetic resources of forage grasses. Genetic Resources and Crop Evolution., 1995,42:197-209.
25. Конарев А.В., Гаврилюк И.П., Мигушова Э.Ф. Дифференциация диплоидных пшениц по данным иммунохимического анализа. Докл. ВАСХНИЛ, 1974, 6:12.
26. . Дорофеев В.Ф., Удачин Р.А., Семенова Л.В. и др. Пшеницы мира. Агропромиздат, Ленингр. отд., 1987: 560с.
27. Chapman V., Miller Т. andRiley R. Equivalence of the A-genome of bread wheat and that of Triticum urartu. Genetic Resources, 1976, 27, 1: 69-76.
28. Dvorak J., McGuire P. and Cassidy R. Apparent sources of the A genome of wheat inferred from polymorphism in abundance and and restriction fragment length of repeated nucleotide sequences. Genome, 1988, 30: 680-689.
29. Potokina E., Tomooka N., Dunkan A. Vaughan, Alexandrova T. & Ru-QiangXu. Phylogeny of Vicia subgenus Vicia (Fabaceae) based on analysis of RAPDs and RFLP of PCA-amplified chloroplast genes. Genetic resources and crop evolution. 1998, 3: 1-13.
30. Перчук И.Н., Лоскутов И.Г., Окуно К, Эвана К. RAPD-анализ в изучении межвидового полиморфизма в роде Avena L. В материалах IV Совещания по кариологии и кариосистематике растений (Спб, 25-27 мая 1999г).
31. Алтухов Ю.П. Генетические процессы в популяциях. М.: Наука, 1988, 328с.
32. Marshall D. and Brown A. Optimum sampling strategies in genetic conservation. In: Genetic resources for today and tomorrow. Cambridge University Press, Cambridge, UK, 1975.
33. Пенева Т.И., Конарев B.F., Кобылянский В.Д., Лапиков КС Анализ полиморфизма по спектрам секалина в процессе становления сорта озимой ржи Ильмень. Сельскохозяйственная биология, 1998,1 : 55-62
34. Vvedenskaja I. O., Alpatyeva N. V., Gubareva N.K. and Konarev A. V. Use of storage protein electrophoresis in the analysis of genetic resources of some cereals. Erhaltung and nutzung pflanzengenetischer ressourcen - eine internationale aufgabe fur naturschutzer, genbanken und pflanzenzuchter. Vortrage fur pflanzenzuchtung, 1993, 25: 187-201.
35. Алпатьева Н.В., Губарева H.K. Изучение динамики генотипного состава стародавних сортов озимой мягкой пшеницы по белкам зерна в разных условиях хранения и репродукции образцов. В тезисах международной конференции: «Интродукция и отдаленная гибридизация растений». Москва, ГБС, РАН, 1998 : 24-26.
36. Идентификация отечественных сортов озимой мягкой пшеницы по электрофорети-ческим спектрам глиадина (под ред. В.Г.Конарева). Каталог мировой коллекции ВИР. Ленинград, ВИР, 1983. Вып.386 : 57с.
37. Применение белковых маркеров для идентификации селекционных материалов сахарной свеклы. Методические указания (под ред. В.Г.Конарева). Ленинград, ВИР, 1991: 17с.
35. Фарбер СП., Артемьева A.M. Полиморфизм круциферина семян видов рода Brassica L. Сельскохозяйственная биология, 2000 (в печати).
39. Анализ и регистрация линий, сортов и гибридов кукурузы по зеину методом электрофореза. Методические указания и каталог белковых формул (под ред. В.Г.Конарева). Санкт-Петербург, ВИР, 1998 :49с.
40. Малиновский Б.Н., Конарев А.В. Использование электрофореза запасных белков в селекции сорго. Тезисы докладов научно-практической конференции (Зерноград, 2-4 сентября 1999). Зерноград, 1999 : 63.
41. Konarev А. К, Khomutnikova L.A., Malinovskiy B.N. The use of prolamine electrophoresis in breeding sorghum for heterosis and in marking sorghum biotypes of value for breeding. Extended synopses FAOMAEA International Symposium on the Use of Induced Mutations and Molecular Techniques for Crop Improvement. Austria, Vienna, 1995:201-202.
42. Методические материалы (Изменения и дополнения к методике государственного сортоиспытания сельскохозяйственных культур). М., 1984, 1(33): 68с.
43. Сорта сельскохозяйственных культур, впервые включенные с 1993 года в государственный реестр сортов, допущенных к использованию в производстве. МСХ РФ. М., 1993: 164 с.
44. Применение электрофореза белков в первичном семеноводстве зерновых культур. Методические указания (под ред. В.Г.Конарева и В.Г.Еникеева). Санкт-Петербург, ВИР, 1993: 42с.
45. Рекомендации по использованию белковых маркеров в сортоиспытании, семеноводстве и семенном контроле (под ред. В.Г.Конарева). Москва-Ленинград, Госагропром СССР, ВИР, 1989: 20с.
46. Конарев В.Г. Молекулярно-биологические исследования генофонда культурных растений в ВИРе (1967-1997). СПб, ВИР, 1998: 99с.
Для подготовки данной работы были использованы материалы с сайта http://www.vir.nw.ru/
Похожие работы
... Здесь изобилие кажущееся. Органика рассеивается по огромному пространству. И уже из-зз одного этого не обеспечивается хотя бы относительное постоянство среды, без которого не могла возникнуть жизнь. Ее низкомолекулярные органические предшественники должны были находиться в очень концентрированном состоянии, чтобы образовались биополимеры. И последних тоже должно быть достаточно много при ...
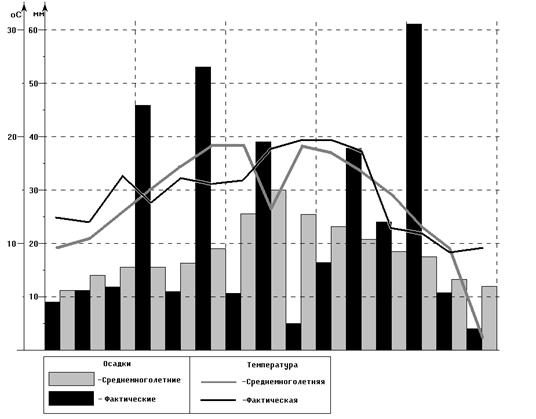
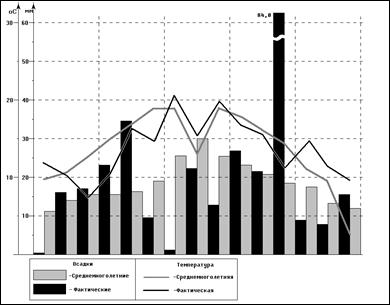
... пшеницы сорта эритроспермум 59 Проведенные исследования (таблица 10) свидетельствуют о том, что наименьший урожай яровой пшеницы Эритроспермум 59 наблюдается на безгербицидном варианте, без Гуми-М (1,88 т/га). Таблица 10 - Влияние обработок посевов гербицидом Пума супер 100 и адаптогенным препаратом на урожайность яровой пшеницы (2001-2003 гг.) Вариант (норма расхода гербицида, л/га) ...
... % Р=ЧД/С, (2) Стоимость продукции является произведением валового сбора зерна с 1 га на его закупочную рыночную стоимость. Таблица 6 Показатели экономической эффективности возделывания сортов мягкой яровой пшеницы Показатели Тулунская 12 Омская 26 Фора Урожайность с 1 га, т/га 2,9 3,2 3,3 Производственные затраты, руб. 2393,93 2396,26 2397,03 Стоимость продукции, руб. 4350 ...
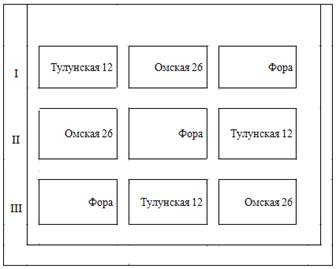
... . Исследования проводились в 2000-2001 годах. Объектами исследований в проведенных опытах были яровая пшеница скороспелого сорта Фора, возделываемая на черноземах выщелоченных тяжелосуглинистых Красноармейского района Челябинской области. Характеристика сорта ОРИГЕНАТОРЫ: КНИИЗХ, ВИР Апробационные признаки: Разновидность лютесценс. Колос пирамидальный, среднерыхлый, средней длины. Остевые ...
0 комментариев