План:
Введение
Классические условные рефлексы
Выработка условного рефлекса
Привыкание
Инструментальное научение
Научение избеганию врагов
Научение типа инсайта
Ассоциативное научение
Импринтинг как научение
Заключение
Список использованной литературы:
Введение
Научение – это адаптивное изменение индивидуального поведения в результате предшествующего опыта. Механизмы научения животных сложны потому, что простые формы научения, такие, как привыкание, сосуществуют с более сложными процессами. Например, у людей мы находим привыкание, условные рефлексы, инструментальное научение, рассудочную и когнитивную деятельность. Животные, может быть, не так легко научаются на основе опыта и рассудочной деятельности, однако многие из них в известной степени, несомненно, обладают этими способностями. Правда, разные виды животных значительно различаются по своей экологии и эволюционной истории. Не значит ли это, что естественный отбор формировал способности к научению каждого животного в соответствии с его экологической нишей? Многие биологи поддерживают эту точку зрения. Психологи склонны считать, что некоторые механизмы научения являются общими для многих видов. Оправдан ли такой взгляд? В настоящей работе мы пытаемся ответить на этот вопрос. Психологи пытались установить общие законы научения, но до сих пор все эти попытки были неудачными. Некоторые свойства среды не меняются, и мы обычно видим, что реакция животных на них стереотипная, врожденная. Некоторые свойства среды меняются на основе суточных, сезонных и т. д. циклов. Обычно животное реагирует на предсказуемые с эволюционной точки зрения изменения среды с помощью заранее запрограммированных изменений своей структуры. В некоторых случаях поведение животных меняется при созревании без участия научения, однако иногда научение для этого необходимо, но является заранее запрограммированным, т. е. Происходит на определенной стадии развития животного более или менее независимо от изменчивости индивидуального поведения. Когда проблемы, встающие перед молодым животным, слегка меняются от поколения к поколению, особи, предрасположенные к узнаванию определенных особенностей среды, близких родичей и источников пищи, могут получить определенное преимущество. Такой вид научения является формой «сопряженного созревания», при котором процессу развития сопутствует приобретение определенного опыта. «По-видимому, научение представляет собой индивидуальный процесс и осуществляется у разных видов и в разных обстоятельствах по-разному. В этой работе я постаралась охватить весь спектр существующих представлений о типах научения и их экспериментальное подтверждение.
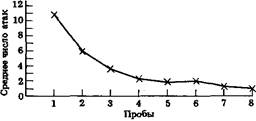
Типичная кривая научения. На графике показано число нападений каракатицы на креветку, находящуюся в стеклянном сосуде. Креветку показывали каракатице 8 раз, каждый раз по 3 мин. Ток как попытки напасть на креветку не приводили к успеху, число атак уменьшалось, по мере того как каракатица усваивала то, что она не может схватить креветку. Приведенная кривая основана на усреднении результатов, полученных на 40 каракатицах. (Messenger J.B., Symp. Soc. Lond., 1977).
Классические условные рефлексы Выработка условного рефлексаВ своих исходных опытах по условным рефлексам Павлов ограничивал движения голодной собаки лямками и давал ей небольшие порции пищи через правильные промежутки времени. Когда он предварял подачу пищи внешним стимулом, например звонком, поведение собаки по отношению к этому стимулу постепенно менялось. Животное начинало поворачиваться к звонку, облизываться и выделять слюну. Когда Павлов стал систематически регистрировать слюноотделения, вводя животному фистулу для сбора слюны, он обнаружил, что ее количество возрастало по мере увеличения числа сочетаний звонка и пищи. Стало видным, что собака научилась связывать звонок с пищей. Павлов назвал звонок условным стимулом (УС), а пищу безусловным (БС), слюноотделение в ответ на предъявление пищи он назвал безусловным рефлексом (БР), а в ответ на звонок - условным рефлексом (УР).
Условные рефлексы, при выработке которых применяется мотивационно благоприятное, или положительное, подкрепление, аналогичное БС, являются примерами положительных условных рефлексов. Но условные рефлексы можно выработать и в опытах с применением отрицательного подкрепления, например удара тока, который животное стремится избежать. Так, включение звукового стимула (определенного тона) перед направлением в глаз кролика струи воздуха начнет вызывать условный мигательный рефлекс, т.е. смыкание век (точнее говоря, мигательной перепонки). Первоначально мигание вызывает только воздушный толчок (БС), но после нескольких повторений мигание (УР) происходит в ответ на данный тон (УС) и в отсутствие действия воздуха. Это пример отрицательного УР.
Подкрепление характеризуется не столько присущими ему стимуляционными свойствами, сколько своим мотивационным значением для животного. Так, пища действует как положительное подкрепление, только если собака голодна, а воздушный толчок действует как отрицательное подкрепление, только если он вреден или неприятен животному. Во многих случаях подкрепление является врожденным в том смысле, что его мотивационное значение и способность поддерживать условный рефлекс являются составной частью нормальной реакции животного на окружающие условия. Однако это не обязательно так, и Павлов показал, что как подкрепление может действовать и УС. Допустим, что при обычной выработке УР звонок становится УС и всегда вызывает этот УР, например слюноотделение. Если затем второй УС, например свет, многократно сочетается со звонком в отсутствие пищи, то у животного образуется УР на один только свет, даже если пища никогда прямо не сочеталась с ним. Такой метод называется выработкой условного рефлекса второго порядка.
Павловские, или классические, условные рефлексы очень широко распространены в животном мире и пронизывают все стороны повседневной жизни высших животных, включая человека. Павлов показал, что условные рефлексы могут вырабатываться у обезьян и мышей; сообщалось также о выработке условных рефлексов у очень многих беспозвоночных. Однако, оценивая такие сообщения, мы должны отличать истинные классические условные рефлексы от других форм научения и от псевдонаучения.
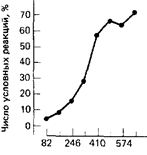
сочетания
Рис. 1. Выработка мигательного условного рефлекса. (По Scheidermann et ai., 1962)
ПривыканиеМногократное применение стимула ведет к повышенной реактивности. Этот феномен, называется привыканием, представляет собой форму не ассоциативного научения. Например, реакции избегания у рыбы в ответ на проходящую над ее головой тень постепенно ослабевает при повторении этого стимула через каждые несколько минут, пока рыба не перестает реагировать на него. Подобным же образом ориентировочная реакция жабы на потенциальную добычу постепенно снижается, если не съедобные, похожие на пищу предметы предъявляются многократно. Садоводы хорошо знают, что чучела, поставленные для отпугивания птиц, действуют только короткое время и птицы скоро привыкают к ним. Попытки отпугивать птиц от аэродромов передаваемыми по радио криками тревоги натолкнулись на те же проблемы привыкания.
Реакции, на которые развилось привыкание, спонтанно восстанавливаются при отмене стимуляции. Если привыкание с последующим восстановлением реакции повторять неоднократно, то привыкание постепенно становится более быстрым. Если в процессе привыкания предъявить или стимул, то реактивность возрасту как показано на рис.2. Это отвыкание, как считают, объясняется изменениями в уровне активации ЦНС животного и очень похожа на Павловское растормаживание. Привыкание обычно рассматривают как форму научения, и его можно экспериментально отличить от затухания реакции при сенсорной адаптации или утомлении. Привыкание сходно с угасанием тем, что животное научается тормозить акции, за которыми не следует подкрепления. В обоих случаях наступают спонтанное восстановление и растормаживание под действием посторонних стимулов. Реакции, на которые обычно развивается привыкание, являются врожденными, а не выработанными с участием УС.
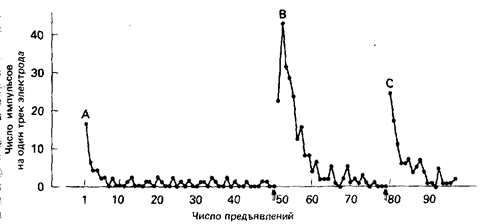
Рис. 2. Привыкание и стимулированное отвыкание в нейроне, реагирующем на двигательные стимулы. А - исходное привыкание; В и С - привыкание, вызванное внешними стимулами (стрелки). (По Rowell, Horn, 1968.)
Инструментальное научениеВ то время как исследование классических условных рефлексов было начато в России, принципы инструментальных условных рефлексов были открыты и разработаны в США.
Торндайк провел ряд опытов, в которых кошки должны были нажимать на задвижку или тянуть за пружину, чтобы, открыв дверцу, выйти из ящика и получить снаружи пищу. Ящики были сделаны с вертикальными щелями так, что пища была заметна. Голодная кошка, впервые посаженная в ящик, производит множество действий, в том числе тянется к пище через щели и скребет предметы, находящиеся внутри ящика. Наконец она случайно ударяет по запирающему механизму и выскакивает наружу. При последующих пробах действия кошки постепенно концентрируются вблизи этого механизма, и прочая активность со временем прекращается. Наконец, кошка становится способной правильно вести себя, как только ее поместят в ящик.
Торндайк[1] назвал научение такого типа “методом проб, ошибок и случайного успеха”. Теперь оно называется инструментальным научением, поскольку правильная реакция, ведущая к вознаграждению, требует использования “инструментов”. Цирковые дрессировщики знали о научении такого типа в течение столетий, но Торндайк первый исследовал его систематически и создал на основе своих наблюдений стройную теорию.
Для объяснения изменений, происходящих в поведении животного в этих опытах, Торндайк[2] предложил свой “закон эффекта”. Он гласит, что для реакции, за которой следует вознаграждение или состояние удовлетворения, вероятность повторения возрастает, а для реакции, вызывающей вредное или неприятное последствие, вероятность повторения снижается. Таким образом, успех инструментального научения приписывается тому факту, что выработанное поведение может быть непосредственно изменено его последствиями. Торндайк[3] считал, что подкрепление повышает вероятность реакции, с которой оно сочетается, потому что усиливает установленную связь между ней и присутствующей стимульной ситуацией. Этот взгляд стал известен как “теория научения по принципу стимул-реакция”, и варианты этой теории господствовали в течение многих лет. Признавая значение закона эффекта как эмпирического факта, современные психологи в то же время сомневаются в том, что поведение непосредственно модифицируется своими последствиями, как предполагали Торндайк и его школа.
Научение избеганию враговРеакции избегания представляют собой форму оборонительного поведения, за счет которого животные сводят к минимуму попадание в ситуации, представляющиеся опасными. Стимулы, вызывающие страх, могут быть сигнальными и вызывать реакцию без какого-либо предварительного опыта. Так же могут действовать и стимулы, с которыми реакция страха или избегания образовала условную связь. К вызывающим врожденное поведение избегания относятся стимулы, связанные с хищниками (например, силуэты, похожие на ястреба, которых избегают птенцы) и с ядовитыми растениями и животными, например змеями и грибами. Врожденное поведение избегания весьма неодинаково у разных видов. Оно выражается в оцепенении (в особенности у животных, обычно хорошо замаскированных), в бегстве, поиске укрытия и тигмотаксисе (т.е. в прижимании к предметам и избегании открытых пространств), в предупреждающих сигналах, служащих для отпугивания нападающего или для оповещения других животных.
В естественной среде обитания животное, обучающееся избегать хищников посредством проб и ошибок, пережило бы лишь несколько таких проб. Поэтому животные большей частью обладают врожденными защитными реакциями, рефлекторными по своей природе, хотя и изменяемыми научением. Р. Боллс (R.Во1les[4], 1970) указал, что экспериментатор в опыте по научению избеганию намеренно или в силу устройства аппаратуры выбирает эффективную реакцию избегания, которая будет или не будет совпадать с врожденной реакцией животного. Степень совместимости врожденного и вырабатываемого в эксперименте поведения сильно влияет на легкость, с какой приобретается реакция избегания.
В течение многих лет психологи сообщали о значительных различиях в легкости, с какой животные научаются реакциям избегания (Mackintosh, 1974[5]). Так, крысы приблизительно за пять проб научаются перебегать из одного ящика в другой, избегая удара током, но им требуются сотни проб, чтобы для избегания тока научиться нажимать на рычаг. Голуби в этих условиях с большим трудом научаются клевать ключ, но с меньшим - нажимать на педаль. Еще один усложняющий фактор-разница врожденных реакций избегания в зависимости от ситуации. Так, в ответ на удар током крысы обычно убегают, когда есть возможность убежать, и замирают, когда ее нет. Если в такой ситуации присутствует какая-то “мишень”, например другой представитель этого же вида, действие тока может вызвать агрессивное поведение (Logan, Boice, 1969[6]). Если ситуация требует, чтобы животное приблизилось к стимулу, связанному с током, тогда выработка реакции избегания становится очень трудной. Такое наблюдается, когда крыса должна нажать на рычаг прямо под источником света, предупреждающим о токе. Если же сигнальный свет находится далеко от рычага, выработать условную реакцию гораздо легче (Biederman et al., 1964[7]). Применение тока в аппарате с открытым пространством продлевает время, которое крысы проводят в тесном соприкосновении со стенкой (Grossen, Kelley, 1972[8]). Избегая удара током, эти животные быстрее научаются прыгать на возвышение, находящееся рядом со стенкой, чем на расположенное в середине такого аппарата.
Согласно традиционной теории научения, врожденные оборонительные реакции представляют собой БР. Павловский анализ научения такого типа включает два кардинальных положения (Mackintosh, 1974[9]). Во-первых, успешные реакции избегания тесно связаны с реакциями, которые обычно вызываются неблагоприятным стимулом. Как мы видели, научение избеганию идет гораздо быстрее, если рефлекс, вырабатываемый экспериментатором, совместим с естественной реакцией животного на применяемое “наказание”. Мы вкратце проиллюстрируем этот принцип в другом месте.
Второе павловское положение гласит, что выработка реакции избегания не должна непосредственно зависеть от возможности избегания как такового. Именно применение отрицательного подкрепления усиливает реакцию, вызываемую этим подкреплением. Отсутствие подкрепления, являющееся или не являющееся результатом избегания, не улучшит процесс классической выработки УР. Здесь, однако, стоит указать, что отсутствие подкрепления иногда не оказывает действия, а иногда оказывает сильное влияние на научение избеганию (Mackintosh, 1974[10]). Так, например, серебряные караси плывут из одного отделения аквариума в другое, когда это избавляет их от удара током, но они плывут с той же готовностью, когда этот удар неизбежен. И напротив, отключение тока служит эффективным подкреплением, заставляющим морских свинок бегать в колесе (Brogden et al., 1938[11]) или крыс перебегать из одного отделения ящика в другое (Miller, 1948[12]). Эта методика лучше применяемой при классической выработке УР (рис. 3)
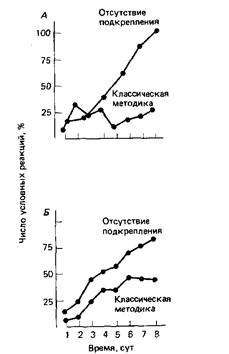
Рис 3. Сравнение классической методики подкрепления и методики его отмены. А. - Бег морских свинок в колесе. Б. - условно-рефлекторное сгибание ноги у собаки. (По Wahlsten, Cole, 1972)
Макинтош (Mackintosh, 1974[13]) указывает, что разница между классическим и инструментальным подходами к выработке реакции избегания может быть связана с вопросом о том, сигнализируют ли внешние стимулы об ударе током или о безопасности и образуют ли реакции на эти стимулы адекватные обратные связи у животного. В тех случаях, когда животное избегает тока, выпрыгивая из ящика или убегая от опасного стимула, само избегание отлично от других форм поведения животного и переносит его в иную внешнюю стимульную ситуацию. Впрочем, для непрерывно плавающего карася уплывание в определенное место (для избегания тока) не очень отличается от обычного поведения, В то же время для неподвижно сидящей крысы убегание туда, где она не получит удара током, представляет собой особую реакцию с ее очевидными последствиями. Поэтому не следует удивляться тому, что крысе легче научиться реакции избегания такого типа, чем нажиму на рычаг.
Крысы, которые могут нажать на рычаг, чтобы избежать удара током и попасть в отделение ящика, где тока никогда не бывает, научаются нажимам быстрее, чем крысы, которые могут нажимать на рычаг, но при этом не получают доступа в такое отделение (Masterton, 1970[14]). Простой нажим на рычаг оставляет животное в более или менее той же внешней стимульной ситуации, что и раньше, а убегание в другое отделение удаляет его от стимула, связанного с током. Научение избеганию идет гораздо эффективнее в перегороженном ящике с одним “электрическим” отделением, чем в таком же ящике с поочередным включением тока в обеих его половинах, вероятно, потому, что в последнем случае нет участка, ассоциирующегося у крыс с безопасностью.
Научение типа инсайтаИдея об участии в научении животного когнитивного процесса имеет долгую историю. Она нашла ясное выражение в работах школы гештальт-психологов, которые считали, что животные способны к инсайту, т.е. что животные способны решать проблемы посредством инсайта (“озарения”) благодаря врожденной тенденции воспринимать ситуацию как целое.
Классическая серия опытов была проведена Кёлером между 1913 и 1917 гг. Во время первой мировой войны Кёлер был интернирован на острове Тенерифе. Там он занялся исследованием шимпанзе на опытной станции по изучению человекообразных обезьян и опубликовал результаты своей работы в книге “"The Mentality of Apes" (“Мыслительные способности высших обезьян”), вышедшей в 1925 г. В опытах Кёлера шимпанзе для получения вознаграждения (корма) должны были пользоваться орудиями. Так, в одном случае шимпанзе нужно было доставать палкой еду, находящуюся вне клетки. Кёлер считал, что его задачи отличались от задач Торндайка очень важной деталью. Животные Торндайка не могли понять, как действует замок, открывающий дверцу их клетки, потому что он находился снаружи и был скрыт от глаз. Они могли решать поставленную задачу только путем проб и ошибок. А в опытах Кёлера все составляющие, необходимые для решения задачи, находятся в пределах видимости животного.
Одной из шимпанзе в опытах Кёлера давали две бамбуковые палки, ни одна из которых не была достаточно длинной для того, чтобы достать плод, лежащий вне клетки. Но палки можно было соединить и получить из них одну, более длинную. После многих безуспешных попыток дотянуться до плода одной из коротких палок шимпанзе прекратила попытки, принялась играть с палками и случайно соединила их, всунув более тонкую в полый конец второй. Тогда обезьяна подпрыгнула и немедленно побежала к прутьям клетки, чтобы достать плод длинной палкой. Кёлер истолковал это, как пример инсайта. В другом опыте плод был подвешен высоко под потолком клетки. Его можно было достать, поставив друг на друга ящики. Некоторые шимпанзе научались решать эту задачу.
Объясняя результаты своих опытов, Кёлер пришел к выводу, что его животные проявляли инсайт. Этот термин обозначает понимание отношений между стимулами или событиями. Научение по принципу инсайта отличается от научения методом проб и ошибок, потому что полагает внезапное возникновение новой реакции. Философа Бертрана Рассела позабавила разница между взглядами Торндайка и Кёлера. Он писал: “У всех тщательно наблюдавшихся животных обнаружились национальные черты наблюдателя. Животные, которых изучали американцы, бешено прыгают с невероятными напором и темпераментом, и, наконец, случайно получают желаемый результат. А животные, которых наблюдали немцы, сидят спокойно, думают и, в конце концов, выдают решение из своего внутреннего сознания”.
Толкование решения задачи животным с точки зрения гештальт - психологии подвергнуто критике по многим причинам. Опытами предполагается определить, ведут ли себя животные на основе инсайта в условиях, которые, как считается, требуют именно такого поведения. Независимые доказательства того, что задача действительно требует инсайта, отсутствуют, если животному удается решить ее, значит явление инсайта доказано. Не все принимают такой ход рассуждений. Еще одна проблема связана с тем, что очень трудно выяснить, является ли реакция, предположительно основанная на инсайте, подлинно новой. Действительно, шимпанзе совершают большое количество посторонних действий, играют и безуспешно пытаются получить еду. Не означает ли это, что они приходят к решению задачи в результате накопления проб и ошибок?
Этот вопрос был изучен Шиллером (Schiller, 1952[15]), который систематически исследовал врожденные компоненты поведения шимпанзе при решении задачи. Например, в одной работе он дал 48 новым шимпанзе, соединяющиеся попарно палки, не поставив перед ними никакой задачи. В течение часа 32 обезьяны соединили палки, причем 19 из 20 взрослых животных проделали это за пять минут. На основании этих и других исследований возникло сомнение в правильности гештальт - психологического подхода. Очевидно, очень важен предыдущий опыт животных. Умение обращаться с палками и ящиками очень сильно сказывается на способе решения задачи. Шимпанзе, которым дают играть с этими предметами, узнают об их свойствах. Способность использовать имеющиеся орудия так, как это нужно для решения задачи, во многом зависит от зрелости обезьяны. Обнаружив, что с предметами можно совершать определенные действия, животное будет повторять их снова и снова. Некоторые манипуляции просто слишком сложны для молодых шимпанзе, но после включения в поведенческий репертуар могут совершаться в самых различных ситуациях.
Основные различия между инсайтом и другими формами научения, по-видимому, состоят в способности более умных животных пользоваться опытом, приобретенным в других условиях. Но это необязательно значит, что инсайт должен рассматриваться отдельно от научения. Способности животных решать задачи трудно исследовать, так как человек может плохо представлять себе, как животное видит данную ситуацию. Совершенно очевидно, что не следует задавать животному задачу, которая лежит за пределами его манипуляционных возможностей или противоречит его естественным склонностям. В то же время каждый вид хорошо приспособлен природой к выполнению некоторых на первый взгляд “умных” действий, считать которые свидетельством “инсайта” или “интеллекта” неверно. В действительности эти термины не более чем условные обозначения для явлений, еще требующих объяснения.
Ассоциативное научениеДля рассмотрения гипотезы о том, что животные обладают механизмами обнаружения и узнавания причинных отношений, следует дать определение природы таких отношений. Существуют два основных типа причинных связей, и вряд ли можно сомневаться в том, что оба их животное может научиться различать (Dickinson, 1980[16]). Одно событие (причина) может вызвать другое событие (следствие) или не вызвать его (отсутствие следствия). Первое событие не обязательно должно быть непосредственной причиной следствия или его отсутствия, но оно может быть определенным звеном причинно-следственной цепи. В самом деле, событие, замеченное животным, может быть не частью причинной цепи, а просто указанием на то, что произошло вызванное данной причиной событие. Именно кажущаяся причина важна для животного.
Способность научиться узнаванию обоих типов причинных отношений можно показать на простом опыте. Голодных голубей сажают в скиннеровский ящик с двумя световыми ключами и механизмом подачи корма. Одной группе голубей демонстрируют связь свет-корм (причина-следствие), а другой - свет -• отсутствие корма (причина без следствия). Третьей группе предъявляют только свет. В первом случае один из ключевых дисков освещается на 10с с неправильными интервалами, и пища подается, как только выключается свет. Во втором случае свет и пищу предъявляют столько же раз, что и в первом, но внимательно следят за тем, чтобы корм никогда не появлялся вслед за включением света. В третьем случае совсем не дают пищи. В первом случае свет сигнализирует о появлении корма, а во втором - о его отсутствии. Затем свет можно предъявить каждой группе голубей отдельно от пищи и отметить, приблизится к нему птица или, наоборот, удалится. Результаты такого опыта ясно показывают, что голуби, наблюдавшие связь свет-корм, стремятся приблизиться к свету, как и можно, ожидать на основе нормальной классической выработки условного рефлекса. Голуби, которым предъявлялось отношение свет-отсутствие корма, также не оставались безразличными к свету, а определенно избегали его (Wasserman et al., 1974[17]). Лишь голуби третьей группы были, по-видимому, безразличны к свету (рис. 4)
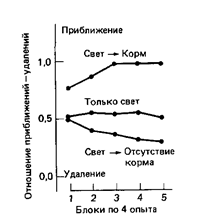
Рис. 4. Приобретение голубями, которым предъявляются разные сочетания освещения ключа и подачи корма, тенденции приближаться к освещенному ключу или отдаляться от него. В условиях “свет корм” освещение ключа сочеталось с кормом, и голуби вырабатывали реакцию приближения. В условиях применения только света голуби никогда не получали корма и никак не реагировали на ключ. В условиях “свет-отсутствие корма”, они получали корм, но ни во время, ни вскоре после освещения ключа; у этих голубей возникала реакция удаления от ключа. (По Dickinson. 1980.[18])
Анализируя результаты этого опыта, надо помнить о ряде важных моментов. Во-первых, мы не можем считать, что голубь научился устанавливать причинную связь света с пищей или ее отсутствием. То, чему животные научаются, сталкиваясь с причинными отношениями, можно рассматривать только с позиций теорий о внутренних изменениях, происходящих в результате приобретенного опыта. Во-вторых, тот факт, что в одном случае голуби приближаются к свету, а в другом удаляются от него, интересен, но не относится непосредственно к обсуждаемой проблеме. Многие различия в поведении, вызванные предъявлением стимулов, можно было бы счесть указанием на произошедшее научение, однако говорить о значении конкретных действий животного логично только при анализе того, чему именно оно научилось. В-третьих, имеет смысл считать свет кажущейся причиной непредъявления пищи, только если пища предъявляется в его отсутствие. Нельзя ожидать, что голубь научится принимать свет за причину отсутствия пищи, если он никогда не получал ее в данной ситуации, как это было в третьей опытной группе. Животное имеет возможность усвоить ассоциацию “причина-отсутствие следствия”, только когда почему-либо ожидает, что определенное следствие произойдет (Dickinson, 1980[19]).
Хотя иногда можно сделать вывод, что изменение поведения говорит о произошедшем научении, отсутствие таких изменений нельзя считать указанием на полное отсутствие научения. Дикинсон называет это проблемой молчания поведения. Так, например, в случае описанного выше опыта мы не можем считать, что голуби, которым предъявлялся только свет, ничему не научились. Напротив, имеются данные о том, что крысы научаются игнорировать стимулы, не предсказывающие никакого изменения в ответ на их действия (Mackintosh, 1973[20]). Если в ходе опыта крысам вначале предъявляют два стимула независимо один от другого, то потом они научаются связывать их медленнее, чем животные, которым эти стимулы предварительно не предъявлялись. Следовательно, крысы научаются тому, что определенные стимулы незначимы, и это мешает последующему основанному на этих сигналах научению.
Существуют и другие формы научения, при которых наблюдается молчание поведения. Животные могут научиться тому, что два события не связаны между собой, т. е. следствие не связано либо с данной причиной, либо с целым классом причинных событий. Если этот класс действий входит в поведенческий репертуар животного, то такую форму научения называют “приобретенной беспомощностью” (Maier, Seligman, 1976[21]), т.е. животные научаются тому, что они ничего не могут сделать для улучшения ситуации. Такая выученная беспомощность замедляет будущее научение в аналогичных условиях.
Для того чтобы животное научилось различать простую причинную связь - между двумя событиями должна быть полная положительная корреляция (Dickinson, 1980[22]). С точки зрения животного, всегда имеется множество возможных причин события помимо той, которую применил экспериментатор. Между двумя событиями должна существовать достаточно тесная связь, для того чтобы одно из них было принято за причину второго. Для выяснения значения фоновых, или ситуационных, сигналов Макинтош (Mackintosh. 1976[23]) обучал крыс нажимать на рычаг для получения корма, а затем вводил в опыт различные стимулы. Одной группе при каждом нажиме предъявляли световой сигнал, другим - сложный, состоявший из света и шума. Для одних групп использовался слабый шум (50 дБ), для других - сильный (85 дБ). Во всех случаях животное сразу после предъявления каждого стимула получали легкий удар током. В конце опыта всем группам предъявляли один только свет, чтобы узнать, насколько, животные научились связывать его с током.
Полученные результаты показывают, что свет подавлял нажимы на рычаг в разной степени в разных группах. Если предъявлялся только свет или свет со слабым шумом, подавление было значительным, а когда свет сопровождался ильным шумом, оно было гораздо слабее. Таким образом, наличие мощного второго стимула ослабляло связь света с током даже несмотря на то, что между ними была полная корреляция. Это явление называется затенением. Степень затенения зависит от сравнительной силы затеняемого и затеняющего стимулов, поэтому слабый шум производил незначительное затеняющее действие.
Животные научаются связывать два события, только если вначале сочетание их было неожиданным или внезапным (Mackintosh, 1974[24]). В обычном опыте по условным рефлексам такая внезапность создается подкреплением. Так, если стимул сопровождается ударом тока, причем ни сам этот стимул, ни фоновые сигналы в начале эксперимента не предвещали включения тока, такое подкрепление будет внезапным. Но предположим, что животное уже испытало раньше удар тока в присутствии стимула А; тогда, если с током коррелируют стимулы А и Б, присутствие первого из них будет блокировать выработку реакции на второй. Это явление, впервые открытое Кеймином (Kamin, 1969[25]), называют блокадой. В одном из своих экспериментов Рескорла (Rescorla, 1971[26]) продемонстрировал этот эффект и показал также, что чем неожиданнее подкрепление, тем лучше животное научается
В природе временная связь между какими-либо событиями не всегда является причинной связью. Например, если кошка прыгает на яблоню, когда лает собака, а затем на землю падает яблоко, то мы скорее подумаем, что кошка, а не лай собаки, была причиной его падения. Как прыжок кошки, так и лай собаки одинаково связаны во времени с падением яблока, но другие детали этих событий вводят нас к заключению, что его причиной была все же кошка. Точно так же можем показать, что животные легче образуют ассоциации между одними типами стимулов, чем между другими. Например, крысы легко связывают вкус со следующей болезнью, но нелегко научиваются связывать с ней тон или свет. Показано, что крысы быстрее ассоциируют между собой два события, воспринимаемые одной сенсорной модальностью (Rescorla, Furrow, 1977[27]) или локализованные в одном месте. Наконец, для того чтобы животное связало между собой два события, обычно нужно, чтобы они были очень близки во времени. Временные отношения между двумя событиями изучались во многих опытах по научению (Dickinson, 1980[28]), показавших, что оно наиболее эффективно, когда одно событие (причина) происходит незадолго до второго (следствия). Однако некоторые данные говорят в пользу того, что такие отношения объясняются не прямым действием временного интервала на процесс научения, а различной степенью затенения первого события фоновыми сигналами в зависимости от временного интервала (Dickinson, 1980[29]). А это означает, что влияние временного интервала на научение должно зависеть от значимости фоновых сигналов. В частности, следует ожидать, что фоновые сигналы будут гораздо менее важны при выработке ассоциации вкуса с болезнью, чем при сочетании тона с током, потому что присутствующие в экспериментальной ситуации фоновые стимулы обычно не существенны для научения избеганию определенного вкуса.
Ревуски (Revusky, 1971[30]) первый предположил, что отсутствием затенения, возможно, объясняется выработка вкусового отвращения, которая происходит несмотря на очень длинные интервалы времени между едой и последующим заболеванием. Он показал, что эффективный интервал можно сократить, введя фон из значимых (вкусовых) стимулов.
Импринтинг как научениеЛоренц (Lorenz, 1935[31]) считал, что импринтинг принципиально отличается от форм научения; однако в настоящее время эта точка зрения стала неполной. Импринтинг - сужение предсуществующих предпочтений (a narrowing of pre-existing preferences) - это процесс, который имеет много общего с другими формами перцептивного научения. Вместе с тем Импринтинг это и научение организма реагировать особым образом на импринтированного ситуацию.
Перцептивное научение-понятие весьма неоднозначное, поскольку часто трудно определить, до какой степени животное научается организовывать свой перцептивный мир, а до какой степени его восприятие внешних стимулов является ценным. В многочисленных экспериментах было показано, что животные, которые на ранних этапах своего развития были лишены перцептивного опыта, различали стимулы хуже, чем нормальные животные (см. Hinde, 1970[32]). Однако интерпретация экспериментов с сенсорной депривацией в аспекте перцептивного научения вызывает много возражений. Подвергшиеся депривации животные могут плохо выполнять тесты по различению сигналов, во-первых, потому что они научились полагаться на другие органы чувств, во-вторых, потому что предъявление новых стимулов вызывает у них эмоциональный дискомфорт и, наконец, в-третьих, потому что у них ухудшаются механизмы сенсорного восприятия. Здесь нужно другое. Хорошее доказательство в пользу перцептивного научения могут представить эксперименты, в которых обеспечивалось бы получение специфического опыта с помощью особых стимулов. Для развития полной песни зяблика необходимо, чтобы птица, с одной стороны, прослушала ее в особый период своего раннего развития, а с другой - имела бы возможность практиковаться в исполнении этой песни на более поздней стадии развития. Первую фазу освоения песни, во время которой птица запоминает “образ” полной песни, можно рассматривать как перцептивное научение. Точно так же зебровые амадины, которые были воспитаны бронзовой амадиной и на которых они через несколько лет изоляции реагировали как на половых партнеров, дают нам хороший пример перцептивного научения.
В лаборатории Гибсона был проведен эксперимент, ставший уже классическим. Крысят содержали в клетках, на стенках которых были прикреплены различные металлические фигуры. Затем оценивалась способность крыс приближаться к одной из двух таких фигур, чтобы получить пищевое подкрепление. Авторы обнаружили, что крысы, ранее видевшие одну из таких фигур, значительно быстрее научались решать данную задачу, чем контрольные животные (Gibson, Walk, 1956[33]). Эти результаты сходны с данными Бейтсона (1964)[34] о том, что цыплята гораздо легче запечатлевают те объекты, с которыми они уже встречались в своем “домашнем” окружении. Все это свидетельствует о большой роли перцептивного научения в процессе импринтирования.
По всей вероятности, импринтинг также имеет отношение и к инструментальному научению. Если утятам на следующий день после вылупления показать игрушечный поезд, то позже их можно научить клевать столбики, если сразу после клевания мимо них будет пробегать эта игрушка. Если утятам показывать поезд на более поздних стадиях их развития, то они уже не могут научиться выполнять этот тест. Бейтсон и Риз (Bateson, Reese, 1969[35]) обнаружили, что утята кряквы и цыплята могут научиться нажимать на педаль, чтобы вызвать включение мелькающего света (к которому у них не было предварительного импринтирования), если такое обучение, проводить в период, чувствительный к импринтированию. Таким образом, представляется вероятным, что цыпленок научается воспринимать свою мать посредством перцептивного научения, а с помощью инструментального обусловливания он научается тем реакциям, которые приводят его к ней. Бейтсон и Уэйнрайт (Bateson, Wainwright, 1972[36]) показали, что по мере ознакомления цыпленка с заимпринтированным стимулом он начинает предпочитать слегка отличающиеся от него стимулы. Способность узнавать родителей “со всех точек зрения” развивается только после того, как птенец построит комплексную картину из всех сенсорных характеристик своего родителя (Bateson, 1973[37]).
Для некоторых типов научения подкрепление не всегда бывает обязательно, однако с помощью подкрепления такое научение может быть модифицировано. Более того, животные способны выполнять даже инструментальные поведенческие навыки, чтобы получить подкрепление в форме песни своего биологического вида или образа полового партнера. Роль подкрепления при импринтировании как раз и относится к этой категории, и поэтому утверждение, что импринтинг - это особая форма научения, не может получить должной поддержки. С функциональной точки зрения импринтинг можно классифицировать как вид предпрограммированного научения. Однако этот термин также приложим и ко многим другим аспектам научения, например научению пению, научению узнавать место и, наконец, латентному научению.
ЗаключениеПодводя итог, можно сказать, что животные научаются ассоциировать два события, если отношение между ними соответствует тому, что обычно называют причинно-следственной связью. Таким образом, они способны усвоить, что одно событие (причина) предвещает второе (следствие) или говорит о его отсутствии в будущем (в отсутствие следствия). Животные могут также научиться связывать некоторые стимулы с отсутствием последствий в данной ситуации или не придавать причинного значения определенному классу стимулов (включая собственное поведение животного). Условия для такого рода ассоциативного научения существуют, если исходить из гипотезы, что животные приспособлены для приобретения знаний о причинных связях в окружающей их среде. Итак, животное должно уметь отличать потенциальные причины от фоновых стимулов, а для этого должно произойти что-то неожиданное, что привлечет внимание животного к данным событиям, или же сами события должны быть (врожденно) значимыми в отношении определенных последствий. Если эти условия не выполняются, фоновые сигналы могут затенить потенциальные причинные события или же научение может быть блокировано предыдущей связью с незначимым в данный момент стимулом. Таким образом, условия ассоциативного научения соответствуют нашим опирающимся на здравый смысл взглядам на природу причинности. Они не согласуются с традиционной точкой зрения на научение как на автоматическую связь между стимулом и реакцией, феномен усваивания информации при молчании поведения наводит на мысль о необходимости некой когнитивной интерпретации научения животного. Но это не должно приводить нас к поспешным выводам о когнитивных способностях животных или о природе их ума.
Список использованной литературы:
1. Д. Мак-Фарленд. Поведение животных. М., Мир. 1988г.
2. Н. Грин, У. Стаут, Д. Дейлор. Биология. В 3-х т. Т. 2, с. 311-312. М., Мир. 1996.
3. Н. Грин, У. Стаут, Д. Дейлор. Биология. В 3-х т. Т. 3, с. 389. М., Мир. 1996.
4. Дикинсон А. Современная теория изучения животных. М., Наука. 1980.
5. Лоренс Ф. Поведение животных. М. 1986.
[1] Thorndeike E.L., 1898, Animal intelligence: an experemental study of the assotiative processes in animals.
[2] Thorndeike E.L.,1913, The psychology of Learning.
[3] Thorndeike E.L, 1911, Animal intelligence.
[4] R. Во1les, 1970, Species-species defense reactions and avoidance learning.
[5] Mackintosh, 1974, The Psychology of Animal Learning
[6] Logan, Boice, 1969, Aggressive behaviours of paired rodents in an avoidance context.
[7] Biederman et al., 1964, Facilitation of discriminated avoidence learning by dissociation of CS and manipulandum/
[8] Grossen, Kelley, 1972, Species-specific behaviour and acquisiton of avoidance behaviour in rats.
[9] Mackintosh, 1974, The Psychology of Animal Learning
[10] Mackintosh, 1974, The Psychology of Animal Learning
[11] Brogden et al., 1938, The role of incentive in conditioning and extinction.
[12] Miller, 1948, Studies of fear as an asquirable drive.
[13] Mackintosh, 1974, The Psychology of Animal Learning
[14] Masterson, 1970, Is termination of a warning signal an effective reward for a rat?
[15] Schiller, 1952, Innate constituents of complex responses in primates.
[16] Dickinson, 1980, Contemporare Animal Learning.
[17] Wasserman et al., 1974, Pavlovian appetitive contingencies and approach versus withdrawal to conditioned stimuli in pigeons.
[18] Dickinson, 1980, Contemporare Animal Learning.
[19] Dickinson, 1980, Contemporare Animal Learning
[20] Mackintosh, 1974, The Psychology of Animal Learning
[21] Maier, Seligman, 1976, Learned helplesseness.
[22] Dickinson, 1980, Contemporare Animal Learning.
[23] Mackintosh. 1976, Overshadowing and stimulus intensity.
[24] Mackintosh, 1974, The Psychology of Animal Learning
[25] Kamin, 1969, Preditstability, supreise, attention and conditioning.
[26] Rescorla, 1971, Variations in the effectiveness of reinforcement and nonreinforcement following prior inhibitory conditioning.
[27] Rescorla, Furrow, 1977, Stimulus similarity as a determinant of Pavlovian conditioning.
[28] Dickinson, 1980, Contemporare Animal Learning
[29] Dickinson, 1980, Contemporare Animal Learning
[30] Revusky, 1971, The role of interference in association over delay.
[31] Lorenz, 1935, Der Kumpan in der Umwelt des Vogels.
[32] Hinde, 1970, Animal behavior (2nd edition).
[33] Gibson, Walk, 1956, The effect of prolonged exposure to visually presented patterns on learning to discriminate them.
[34] Bateson P.P.G., 1964, An effect imprinting on the perceptual development of domestic chics.
[35] Bateson, Reese, 1969, The reinforcing properties of conspicuous stimuli in the imprinting situation.
[36] Bateson, Wainwright, 1972, The effects of prior exposure to light on the imprinting process in domestic chicks.
[37] Bateson, 1973, Internal influences on early learning in birds.
Похожие работы
... . В результате в дополнение к врожденным, видовым программам поведения в центральной нервной системе постоянно формируются новые, благоприобретаемые, индивидуальные программы, на которых зиждутся процессы научения. Физиологические механизмы этого непрерывного программирования поведения были особенно детально проанализированы П.К.Анохиным и получили свое отражение в его концепции «акцептора ре ...
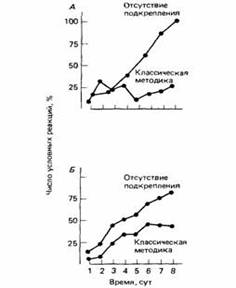
... последствие независимым от питания способом. После гамма-облучения у крыс возникало отвращение к ранее предпочитавшейся воде с сахарином. Гарсия и др. указали на значение эффекта такого типа для психологии научения животных и привели девять подтверждающих его опытов. Однако прошло еще 10 лет, прежде чем весь смысл этого феномена получил должную оценку и ученые поняли, что потребность в ...

... поведения животного. Что бы животное ни делало, ход событий установлен либо экспериментатором, либо аппаратурой, действующей по определенной программе. В отличие от этого при инструментальном научении поведение животного служит одним из важных факторов, определяющих ход событий. Обычно процедура инструментального обучения состоит в том, что подкрепление или наказание подаются или устраняются при ...
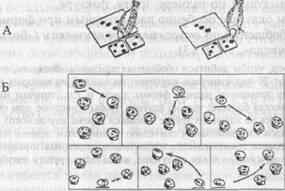
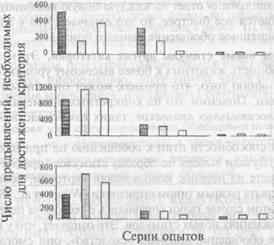
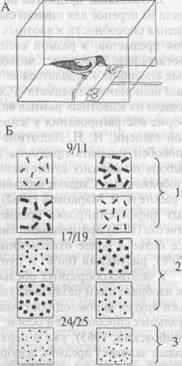
... (формы, размера и др.). Способность ориентироваться на отвлеченные (обобщенные) признаки предметов имеет первостепенное значение для характеристики высших когнитивных функций животных. Рассмотрим подробнее, что известно в целом о способности животных к обобщению и абстрагированию указанных признаков и каких степеней обобщения могут достигать в том или ином случае животные с разным уровнем ...
0 комментариев