Реферат выполнила студентка 12 группы II курса пед/ф-та Пасько С.П..
Саратовский Государственный Медицинский Университет
Кафедра биохимии
Саратов 2004
Промежуточный обмен аминокислот в тканях.
Промежуточный метаболизм аминокислот белковых молекул, как и других питательных веществ в организме, включает катаболические (распад до конечных продуктов) и анаболические (биосинтез аминокислот) процессы, а также ряд других специфических превращений, сопровождающихся образованием биологически активных веществ. Условно промежуточный метаболизм аминокислот можно разделить на общие пути обмена и индивидуальные превращения отдельных аминокислот.
Общие пути обмена аминокислот.
Общие пути превращения аминокислот включают реакции дезаминирования, трансаминирования, декарбоксилирования, биосинтеза и рацемизации. Реакции рацемизации характерны только для микроорганизмов, физиологическая роль которой заключается в синтезе D-изомеров аминокислот для построения клеточной оболочки.
Дезаминирование ( отщепление аминогруппы) – существует четыре типа реакций, катализируемых своими ферментами:
Восстановительное дезаминорование ( +2H+)
Гидролитическое дезаминированиие (+H2О)
Внутримолекулярное дезаминирование
Окислительное дезаминирование (+1/2 О2)
Во всех случаях NH2- группа аминокислоты высвобождается в виде аммиака. Помимо аммиака продуктами дезаминирования являются жирные кислоты, окикислоты и кетокислоты. Для животных тканей, растений и большинства микроорганизмов преобладающим типом реакций является окислительное дезаминирование аминокислот, за исключением гистидина, который подвергается внутримолекулярному дезаминированию.
Кроме перечисленных четырех типов реакций и катализирующих их ферментов в животных тканях и печени человека открыты также три специфических фермента (серин- и треониндегидратазы и цистатионин-γ- лиаза), катализирующих неокислительное дезаминирование серина, треонина и цистеина. Они требуют присутствия пиридоксаль-фосфата в качестве кофермента. Конечными продуктами реакции являются пируват и α- кетобутират, аммиак и сероводород.
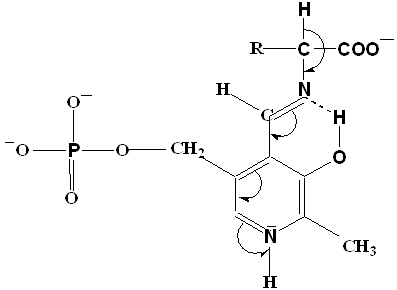
Трансаминирование – реакции межмолекулярного переноса аминогруппы (NH2) от аминокислоты на α-кетокислоту без промежуточного образования аммиака (глутамат+ пируват =
α-кетоглутарат + аланин). Впервые эти реакции были открыты в 1937г. А.Е. Браунштейном и М.Г. Крицман. Реакции трансаминирования являются обратимыми и универсальными для всех живых организмов, они протекают при участии специфических ферментов – аминотрансфераз (трансамниназ). Теоретически реакции возможны между любой амино- и кетокислотой, но наиболее интенсивно они протекают, если один из партнеров представлен дикарбоновой амино- или кетокислотой. В переносе амниогруппы активное участие принимает кофермет трансминаз – пиридоксальфосфат (производное витамина В6). Для реакций трансаминирования характерен общий механизм. Ферменты реакции катализируют перенос аминогруппы не на α -кетокислоту, а на кофермент; образовавшееся промежуточное соединение (шиффово основание) подвергается внутримолекулярным превращениям, приводящим к освобождению α-кетокислоты и пиридоксамнофосфата. Последний на втолрой стадии реагирует с любой другой α-кетокислотой, что через те же стадии приводит к синтезу новой аминокислоты и пиридоксальфосфата.
Декарбоксилирование - отщепление карбоксильной группы в виде СО2, образующиеся продукты реакции называются биогенными аминами, они оказывают сильное фармакологическое действие на множество функций. Эти реакции являются необратимыми, они катализируютя специфическими ферментами – декарбоксилазами аминокмлот- которые в качестве кофермента содержат пиридоксальфосфат ( кроме гистидиндекарбоксилазы и аденозилдекарбоксилазы – содержат остаток пировиноградной кислоты в качестве кофермента). В живых организмах открыты четыре типа декарбоксилирования аминокислот.
α-декарбоксилирование – характерно для тканей животных: от аминокислот отщепляется соседняя от α-углеродного атома карбоксильная группа.
ω-декарбоксилирование- свойственно микроорганизмам
декарбоксилирование, связанное с реакцией трансаминирования. Образуется альдегид и новая аминокислота, соответствующая исходной кетокислоте.
Декарбоксилирование, связанное с реакцией конденсацией двух молекул:
Обезвреживание аммиака в организме.
В организме человека подвергается распаду около 70г аминокислот в сутки: при этом освобождается большое количество аммиака, являющегося высокотоксичным соединением. Поэтому крнцентрация аммиака должна сохраняться на низком уровне (в норме уровень его не превышает 60 мкмоль/л). Концентрация аммиака 3 ммоль/л является летальной.
Одним из путей связывания и обезвреживания аммиака в мозге, сетчатке, почках и мышцах, является биосинтез глутамина( и, возможно, аспарагина). Поскольку глутамин и аспарагин с мочой выделяются в небольших количествах, было высказано предположение, что они выполняют скорее транспортную функцию переноса аммиака в нетоксичной форме.
Часть аммиака легко связывается с α-кетоглутаровой кислотой благодаря обратимости глутаматдегидрогеназной реакции; при синтезе глутамина связывается ещё 1 молекула, т.о. нейтрализуются две молекулы аммиака:
Орнитиновый цикл мочевинообразования.
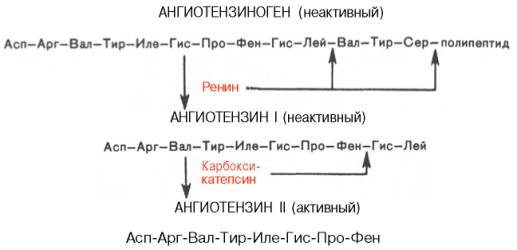
Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины (в основном,в печени).Она выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. На долю мочевины приходится до 80-85% всего азота мочи. Реакции синтеза мочевины, представлены в виде цикла, получившего название орнитинового цикла мочевинообразования Кребса.
На первом этапе синтезируется макроэргическое соединение карбамоилфосфат – это метаболически активная форма аммиака, используемая в качестве исходного продукта для синтеза ряда других азотистых соединений.
На втором этапе цикла мочевинообразования происходит конденсация карбамоилфосфата и орнитина с образованием
цитруллина; реакцию катализирует орнитинкарбамоилтрансфераза:
На следующей стадии цитруллин превращается в аргинин в результате двух последовательно протекающих реакций. Первая из них, энергозависимая, сводится к конденсации цитруллина и аспаргиновой кислоты с образованием аргининосукцината ( эту реакцию катализирует аргининосукцинат-синтетаза). Аргининсукцинат распадается во второй реакции на аргинин и фумарат поддействием аргининосукцинат-лиазы.
На последнем этапе аргинин расщепляется на мочевину и орнитин под действием аргиназы. Суммарная реакция синтеза мочевины без учёта промежуточных продуктов:
Это энергетически выгодная реакция, поэтому процесс всегда протекает в направлении синтеза мочевины.
В состоянии азотистого равновесия организм человека потребляет и соответственно выделяет примерно 15 г азота в сутки; из экскретируемого с мочой количества азота на долю мочевины приходится около 85% , креатинина-около 5%, аммонийных солей – 3%, мочевой кислоты-1% и на другие формы-около 6%.
Типы азотистого обмена. А м м о н и о т е л и ч е с к и й т и п , при котором главным конечным продуктом азотистого обмена является аммиак, свойствен рыбам. У р е о т е л и ч е с к и й т и п обмена - основным конечным продуктом обмена белков является мочевина, характерен для человека и животных. У р и к о т е л и ч е с к и й т и п - главным конечным продуктом обмена является мочевая кислота, характерен для птиц и рептилий.
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://www.monax.ru
Похожие работы
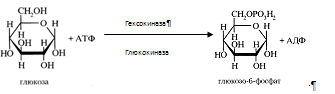
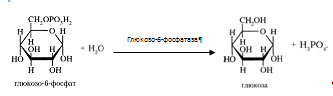
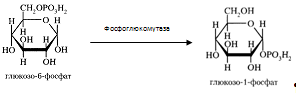

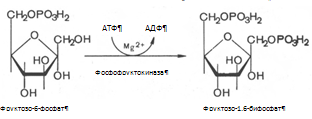
... 3) гепатодепрессии (печёночно-клеточной недостаточности, малой недостаточности печени, недостаточности синтетических процессов); 4) воспаления; 5) шунтирования печени; 6) регенерации и опухолевого роста. При подозрении на конкретную патологию учитываются основные биохимические синдромы, характерные для данного заболевания. За основу берётся стандартная программа функционального обследования, но ...
... активности клеток в кроветворных органах и тканях. Приобретенная аутоагрессивная гемолитическая анемия: в селезенке происходит разрушение и растворение (лизис) эритроцитов. Биохимия и патобиохимия печени. Рассмотрим основные представления о биохимических процессах, протекающих в печени. На пути между кишечником и внутренней средой организма – системой крови и лимфы – находится печень. В ...


... в дозе 2 мг/кг привело к увеличению активности АСТ в гомогенате мозга потомства почти в 2 раза (р <0,05) по отношению к контролю. При изучении токсического действия ионов кадмия на потомство белых крыс было обнаружено, что в гомогенате почек при дозе 0,5 мг/кг активность АСТ по сравнению с контролем достоверно не изменялась (р<0,68) (таблица 2, рисунок Д.2). При дозе 2 мг/кг активность АСТ ...

... Гензеляйт в 1932 г. вывели уравнения реакций синтеза мочевины, которые представлены в виде цикла, получившего в литературе название орнитинового цикла мочевинообразования Кребса. Следует указать, что в биохимии это была первая циклическая система метаболизма, описание которой почти на 5 лет опеределило открытие Г. Кребсом другого метаболического процесса – цикла трикарбоновых кислот. Дальнейшие ...
0 комментариев