Калашников Юрий Яковлевич
АннотацияГены не могут напрямую контролировать и взаимосвязывать сложные биохимические процессы живой клетки. Поэтому они вынуждены кодировать и программировать структурное построение и функциональное поведение молекулярных управляющих средств, к которым, в первую очередь, относятся ферменты и другие клеточные белки. Каким же образом биологические функции могут быть запрограммированы в структурной организации белковых макромолекул? Этот вопрос, видимо, наиболее актуален в современной молекулярной биологии. Предлагая идеи “молекулярной биохимической логики и информатики”, автор в своей статье приводит и конкретные обоснования информационной концепции действия белков и ферментов, которые, как он утверждает, являются молекулярными биологическими автоматами или манипуляторами с программной биохимической логикой управления.
Непревзойденная избирательность действия ферментов и других белков клетки является одной из самых жгучих и волнующих загадок современной биохимии и молекулярной биологии. С ней связаны поиск и нахождение новых подходов и путей к секретам управления обменом веществ и поведения биологических молекул в живых системах. Живая форма материи до сих пор остается одной из самых таинственных мировых проблем. Даже сегодня кажется невероятным, что в ничтожном пространстве одной клетки скрыты все основные характеристики живого, – стремление к четкой структурной и функциональной организации, неуемная жажда активности, размножения и распространения. Однако, несмотря на величайшее разнообразие и необычайную сложность живых форм, – все они имеют единую материальную элементную базу и дискретную молекулярную и клеточную организацию. Все живые клетки в своей основе имеют типовую молекулярно-биологическую систему управления и обладают генетической памятью с феноменальными информационными возможностями. А закономерности молекулярной биохимической логики и информатики, какими пользуются живые системы, по всей вероятности, и являются теми инструментами и механизмами, при помощи которых осуществляется не только структурная организация, но и управление биологической формой материи. Данная статья является продолжением весьма дискуссионной в биологии темы – “молекулярной информатики”, и посвящена информационным аспектам построения и функционального поведения белковых молекул, где попутно изложена и новая альтернативная концепция фермент-субстратных информационных взаимодействий.
1. Принципы и механизмы структурной организации белковых молекул. Программирование построения белковых молекул живой клетки, как известно, осуществляется универсальными клеточными аппаратными средствами транскрипции и трансляции, под руководством генетической информации. Логика структурного построения и функционального поведения белковых молекул определяется генами и природными свойствами стандартных био-логических элементов (аминокислот), которые входят в состав полипептидных цепей. А если учесть типовую дискретную организацию различных биологических молекул и структур, то в обобщенном виде можно сказать, – базовой основой организации биологической формы материи является генетическая информация и общий биохимический алфавит живой материи, который состоит более чем из 30 био-логических элементов (химических букв или символов) – нуклеотидов, аминокислот, простых сахаров, жирных кислот и других мономеров. По меньшей мере, существует два информационных уровня организации белковых (и других) макромолекул. На первом уровне построения – осуществляется последовательное ковалентное соединение соответствующих аминокислот в длинные полипептидные цепи. Так производится позиционное размещение аминокислот в “линейных” цепях, а, следовательно, и декодирование различного рода сигналов, информационных сообщений, инструкций и команд управления, передаваемых генами. Так осуществляется и запись нужного алгоритма структурного преобразования цепи, то есть – программирование трёхмерной организации белковой макромолекулы. При этом, загруженные в “линейную” цепь алгоритмы, – это воплощенные в последовательности аминокислот правила молекулярной биохимической логики, по которым, занесенные в цепь данные воспринимаются как элементарные сигналы, подлежащие исполнению. Здесь под алгоритмом следует понимать последовательность операций, которые выполняются аминокислотами в составе полипептидной цепи, сначала во время конформационного преобразования, а затем, и во время функционального поведения белковой молекулы. Таким образом, программирование структур и функций белковых молекул в клеточной среде осуществляется с помощью унифицированной системы био-логических элементов – аминокислот, которые являются химическими буквами белкового алфавита. А прохождение этих процессов осуществляется соответствующими аппаратными средствами транскрипции и трансляции под руководством генетической информации. Это стало возможным, благодаря тому, что все типовые аминокислоты обладают удивительными природными характеристиками. Все двадцать стандартных аминокислот оказались наделёнными такими химическими и физическими качествами и свойствами, сочетание которых позволяет им, в составе белковых молекул, одновременно выполнять буквально различные био-логические функции и операции. А именно: 1) служить в качестве строительных блоков, с помощью которых осуществляется физическое построение белковых молекул; 2) выполнять роль натуральных информационных единиц – химических букв, с помощью которых записывается молекулярная информация; 3) служить в качестве элементарных единиц аминокислотного кода, с помощью которого сначала идёт преобразование, а впоследствии, – воплощение и реализация генетической информации; 4) быть программными элементам, с помощью которых строятся алгоритмы структурного преобразования, а затем и программа функционального поведения ферментов и других белковых молекул; 5) обуславливать потенциальную и свободную химическую энергию белков и т. д. Здесь мы отметили лишь некоторые из основных направлений применения белкового алфавита. Однако, и из этих примеров ясно, что аминокислотная система действительно обладает уникальными многофункциональными природными качествами и свойствами, которые имеют фундаментальное значение в организации белковых молекул и их функций в любых живых клетках. Важно отметить, что указанные качества и свойства аминокислот существуют всегда и одновременно, и поэтому они, по своей сути, являются разными характеристиками одной и той же элементной базы. Только такое сочетание характеристик, позволяет аминокислотам обеспечивать в живой клетке и информационное построение белковых молекул, и программное управление их биологическими функциями. И всё это может осуществляться только на основе и за счет загруженной (аминокислотным кодом) в белковые макромолекулы структурной, программной и функциональной информации. Причем, каждая типовая аминокислота характеризуется наличием функциональных атомных групп (аминогруппы и карбоксильной группы), которые определяют её химические свойства и служат входными и выходными цепями, с помощью которых элементы могут ковалентно соединяться друг с другом в длинные полипептидные цепи. Кроме того, важно отметить, что каждая аминокислота имеет еще и свою, индивидуальную боковую атомную R-группу, которая в живой системе, как правило, используется в качестве – элементарного информационного химического сигнала! Поэтому, если в информационных технических системах наиболее широкое применение находят электрические сигналы, с переносчиком информации в виде импульсного тока или напряжения, то в молекулярно-биологических системах, в качестве элементарных сигналов, используются химические сигналы различных био-логических элементов общего алфавита – нуклеотидов, аминокислот, простых сахаров, жирных кислот и др., с переносчиком в виде их боковых атомных групп [1]. Наглядный пример: сообщение в цепи ДНК или РНК кодируется в виде последовательности нуклеотидов, а носителями генетической информации являются азотистые основания – “боковые” атомные группы нуклеотидов. Соответственно, и в полипептидной цепи белка это сообщение записывается в виде последовательности аминокислот, где носителями информации являются их боковые R-группы. “Линейную” “структурную основу любого пептида составляет зигзагообразный остов, образованный атомами углерода и азота. Направленные вовне по отношению к остову боковые R-группы любых соседних аминокислотных остатков ориентированы в противоположные стороны. На одном конце в молекуле пептида находится свободная аминогруппа, а на другом конце – свободная карбоксильная группа” [2]. Таким образом, ориентация соседних боковых атомных группировок в противоположные стороны позволяет полипептидной цепи в составе белка осуществлять сначала внутримолекулярные, а затем, и межмолекулярные информационные взаимодействия. Причем, если генетический код служит для переноса генетической программной информации на “линейную” структуру белка, то аминокислотный код является тем молекулярным кодом, с помощью которого осуществляется сначала преобразование, а затем, и, через деятельность белков, – воплощение и реализация генетической информации. Подробное изучение глобулярных и фибриллярных белков показало, что для каждого индивидуального белка характерна своя пространственная трёхмерная организация, которая зависит от его первичной структуры – то есть от информации, записанной линейным аминокислотным кодом. Заметим, что генетическим кодом кодируется только первичная – “линейная” структура полипептидной цепи. Однако “конкретная конфигурация (вторичная, третичная и четвертичная структуры) любого белка полностью определяется первичной структурой входящих в его состав полипептидных цепей и зависит от химических свойств боковых групп аминокислотных остатков” [2]. Следовательно, вторичная, третичная и четвертичная структуры белковых макромолекул кодируются и программируются уже другим молекулярным кодом – аминокислотным. Это ведёт к представлению, что только аминокислотный код обеспечивает трёхмерную структурную организацию белковой молекулы, а затем, и все её специфические свойства и функции. То есть, этот код, в первую очередь, предназначен для организации новой формы молекулярной информации – пространственной, стереохимической. Поэтому первый уровень организации белковых молекул характеризуется применением линейного аминокислотного кода, который служит основой преобразования линейной формы информации полипептидов в стереохимическую форму информации белковых молекул. Здесь следует обратить внимание на то, что различные аминокислоты полипептидной цепи, по всей вероятности, тоже организованны в виде отдельных комбинационных кодовых сигналов, определяющих (в клеточной среде) различные по своей биохимической характеристике зоны, участки и фрагменты, которые обуславливают соответствующие пути, порядок и последовательность информационной сборки белка. В полипептидных цепях белковых молекул кодируется разнообразнейшая информация. Поэтому важно знать, что любая полипептидная цепь всегда является тождественным эквивалентом соответствующего кодового послания генома, указывающего будущие характеристики белковой молекулы. Причем, каждое сообщение, при передаче информации в полипептидной цепи белка, по всей видимости, передаётся своим индивидуальным кодом (кодовыми комбинациями аминокислот). Поэтому информация в цепи может содержать как свою адресную и “операционную”, так и свою структурную и текстовую (информационную) части. Значит, различные информационные сообщения в полипептидных цепях могут быть представлены различными молекулярными кодами и кодовыми комбинациями аминокислотных остатков. Следовательно, в кодовых посылках структуры полипептидной цепи могут быть заключены: 1) адресные кодовые комбинации аминокислотных остатков, которые являются основой формирования адресных стереохимических кодов активного центра фермента (для коммуникативного взаимодействия с молекулами субстрата); 2) “операционная” кодовая комбинация аминокислот, – служит для формирования стереохимического кода операции активного центра, указывающего характер реакции; 3) структурная часть кодовой комбинации аминокислотных остатков, которая кодирует построение и одновременно осуществляет программное обеспечение исполнительных органов и механизмов белковых молекул; 4) текстовая (информационная) часть – кодирует и программирует средства информационной коммуникации белка с различными его молекулярными партнёрами. То есть, “информационная” часть полипептидной цепи предназначена для формирования различного рода локальных или поверхностных рельефных микроматриц, которые обычно располагаются на локальных или поверхностных участках белковой макромолекулы и состоят из многочисленных боковых R-групп аминокислотных остатков. Эффективность применения в живых системах молекулярных кодов обеспечивается многократным циклическим их повторением в структурах типовых биомолекул. Бесконечная череда длинных дискретных сообщений (в виде иРНК, полипептидных цепей и белковых молекул), по своей сути, и представляет собой, ничто иное, как те управляющие информационные потоки и сети, которые осуществляют циклическую передачу информации с целью управления, регулирования и контроля химических превращений и реализации различных молекулярных и других био-логических функций. Однако в живой клетке функционируют только трёхмерные биомолекулы и структуры, поэтому полипептидные цепи, с помощью аминокислотного кода, должны быть преобразованы из линейной формы в пространственную – стереохимическую. Поэтому второй информационный уровень организации белковых молекул – пространственный, осуществляется уже при помощи химических связей, значительно более слабых, чем ковалентные. Это происходит потому, что боковые R-группы тех аминокислот, которые в цепи связаны ковалентно, способны к слабым информационным взаимодействиям с другими боковыми R-группами, как в пределах одной макромолекулы, так и с боковыми группами и атомами близлежащих молекул. К таким взаимодействиям относятся слабые: водородные и ионные связи, ван-дер-ваальсовы силы, гидрофобные взаимодействия, которые в совокупности, благодаря их многочисленности и разнообразию, оказываются весьма сильными. Через посредство этих сил и связей идёт воплощение линейной молекулярной информации в пространственную структуру и стереохимическую форму информации белковых молекул. Cвязывание, взаимодействующих молекулярных структур, как правило, многоточечное. Оно осуществляется за счет участия многочисленных боковых атомных R-групп программных элементов, входящих в состав полипептидной цепи, то есть за счет информации. В результате таких преобразований “одномерная” молекулярная информация полипептидных цепей “сворачивается, пакуется и сжимается” в трёхмерную информацию белковых молекул, которая в таком виде становится пригодной для транспортировки, передачи по различным каналам и компартментам, а затем, и для непосредственного использования в различных биологических процессах. Заметим, что эти информационные силы и связи определяют не только степень прочности белковых макромолекул, но обуславливают и их функциональные возможности. Наличие в структурах белковых макромолекул как внутримолекулярных, так и внешних информационных сил и связей взаимодействия (обусловленных R-группами составляющих их элементов), которые сами по себе слабы, но мощны своей многочисленностью и разнообразием, позволяет говорить о том, что внутри и вокруг макромолекулы образуется специфическое силовое “информационное поле”. Это поле способно влиять как на структуру самого белка, так и на его микроокружение. Поэтому белковая макромолекула как бы стабилизируется самосогласованным сжимающим информационным полем, обусловленным кооперативными силами притяжения между боковыми атомными R-группами аминокислот. А лабильность самой полипептидной цепи в пространственной решетке, с её многочисленными боковыми R-группами элементов, позволяет осуществлять не только точную комплементарную “подгонку” внутримолекулярных структур, но и “подгонку” локальных или поверхностных структур, взаимодействующих друг с другом биомолекул. Трансформация линейных генетических сообщений в трёхмерную структуру и форму различных белковых молекул, – это важный этап перехода биологической информации из одной её молекулярной формы (линейной) в другую (стереохимическую). Линейный и пространственный элементарный состав белков определяется генами, а каждый био-логический элемент (аминокислота) в составе белковой молекулы тождественно может выполнять различные роли – как структурной, так и информационной единицы, как функционального, так и программного элемента. Поэтому все аппаратные средства живой клетки – белки, ферменты и другие клеточные компоненты обладают строго своей специфической структурной организацией, имеют своё информационное и функциональное назначение, а также своё индивидуальное энергетическое и программное обеспечение. И главное, – в результате стереохимических преобразований в структуре белковой молекулы формируются соответствующие молекулярные органы и исполнительные механизмы, а на локальных и поверхностных участках возникает такая пространственно-упорядоченная организация боковых атомных R-групп элементов, которая в живой системе играет роль стереохимических кодовых информационных сигналов. К таким сигналам могут относиться: стереохимические команды управления активного центра фермента (адресный код и код химической операции); различные сигнальные и регуляторные кодовые компоненты; коммуникативные локальные и поверхностные кодовые стереохимические матрицы (микроматрицы), служащие для информационного взаимодействия белковых молекул с их молекулярными партнёрами и т. д. При этом, сама программа функционирования белковой молекулы (благодаря программирующим свойствам элементов) коммутируется лабильными физико-химическими силами, связями и взаимодействиями между боковыми R-группами элементов (аминокислот) в составе её трёхмерной структуры. Поэтому природа взаимодействий боковых атомных групп, определяющих конформационные особенности и внутреннюю динамику белковой макромолекулы, имеет химическую основу и носит информационный характер. Нативная белковая макромолекула как бы стабилизируется самосогласованным сжимающим информационным полем, обусловленным силами притяжения между аминокислотами (программными элементами). А в результате преобразований каждый белок клетки получает своё индивидуальное структурное, информационное, энергетическое, функциональное и программное обеспечение. Поэтому, стереохимический принцип кодирования молекулярной биологической информации применяется живой природой для размещения в одной макромолекуле различных по своему назначению сигналов, сообщений, инструкций, команд управления, а также органов и механизмов их реализации [1]. Такая организация белковых молекул не обладает сильной структурной жесткостью, она всегда достаточно лабильна в тех пределах, которые необходимы для выполнения ими биологических функций. А функциональное поведение макромолекулы, при взаимодействии её с молекулярными партнёрами, определяется свободной энергией и результатом информационного взаимодействия как внутренних, так и внешних составляющих её элементов. Поэтому этот информационный уровень характеризуется уже взаимодействием биологических молекул друг с другом с помощью их локальных, рельефных или поверхностных микроматриц, в результате которых и возбуждаются их биологические функции. Как мы видим, в “молекулярной информатике” открывается большое поле деятельности для исследования информационных путей построения и программного поведения белковых молекул.
2. Адресный код и код операции активного центра фермента представляет собой био-логическую команду управления. Стереохимические коды и микроматрицы, представляющие собой управляющие или коммуникативные сигналы белковых (как, впрочем, и других) макромолекул, возникли в процессе эволюции живой материи и в настоящее время являются основой молекулярных информационных процессов в каждой живой клетке (организме). Все они образованы соответствующей пространственной организацией боковых атомных группировок био-логических элементов (химических букв или символов), входящих в состав кодовых сигналов. Все стереохимические коды и биохимические матрицы белков, образуются во время конформационных преобразований “линейных” полипептидных цепей в трёхмерную структуру и форму. Такое динамическое информационное взаимодействие элементов в составе биологических молекул, которое особенно характерно для белковых молекул, является основой динамического механизма их биологических функций. Биохимическая логика информационных взаимодействий, в частности, предопределяет и протекание химических реакций, так как она основана на явлениях стереохимического узнавания соответствующими ферментами различных био-логических элементов или их функциональных и боковых атомных групп и их химических связей, то есть различных химических букв, символов и знаков биологических молекул субстрата [1]. Стереохимические коды активных центров построены на основе аминокислотного кода, поэтому ферменты могут адресно взаимодействовать с молекулой субстрата и быстро находить нужную им химическую связь и связывающую группу. Кодовые компоненты активных центров ферментов могут комплементарно взаимодействовать с доступными для них функциональными или боковыми атомными группами и атомами молекулы субстрата. Поэтому все субстраты для своих ферментов являются сигнальными молекулами, несущими осведомляющую стереохимическую информацию. На этих принципах основана биохимическая логика информационных взаимодействий между ферментами и их субстратами. Субстраты – это тот химический и информационный материал, который обрабатывается управляющей системой клетки. При этом каждый фермент имеет необходимый и достаточный набор информационных, энергетических, программных и управляющих молекулярных средств, для того, чтобы работать в автоматическом режиме. Процедура управления химической реакции (превращение субстрата), с химической и информационной точек зрения, протекает как полифункциональный катализ, который детерминируется управляющим сигналом, – кодовой комбинацией различных аминокислотных остатков активного центра фермента. Основой стереохимического управляющего сигнала, как правило, служит пространственная кодовая комбинация различных функционально неоднородных амнокислотных остатков активного центра фермента. Причем, аминокислоты, образующие кодовые структуры сигнала, по длине полипептидной цепи обычно находятся далеко друг от друга и оказываются сближенными только при формировании свойственной данному ферменту трёхмерной конформации. Стереохимические коды активного центра обычно состоят из двух зон, имеющих определённое информационное и функциональное назначение. Та пространственная комбинация атомных группировок активного центра фермента, которая осуществляет поиск, узнавание и ориентацию молекулы субстрата, контактирует с непревращаемыми фрагментами субстрата и укрепляет его в активном центре, то есть производит поиск, приём и рецепцию его информации, – представляет собой функциональный стереохимический адресный код фермента. У разных ферментов этот код имеет различное, но строго своё определённое смысловое значение. Та пространственная комбинация атомных группировок активного центра фермента, которая принимает непосредственное участие в синтезе или расщеплении связи субстрата и входит в каталитическую зону, – является кодом каталитической операции, определяющим, в каждом конкретном случае, характер химической реакции [1]. Таким образом, у ферментов формат команды управления может состоять из двух полей: адресного кода, с помощью которого осуществляется динамический поиск и рецепция молекулы субстрата и кода каталитической операции, который определяет характер химической реакции. Во время информационного фермент-субстратного взаимодействия должны быть найдены и комплементарно соответствовать друг другу адресные и каталитические кодовые компоненты фермента и молекулы субстрата. Поэтому биохимическая логика информационных взаимодействий основана на матричных взаимодействиях кодовых компонентов различных биомолекул. По принципу взаимодополняемости локальные или поверхностные микроматрицы молекулярных партнёров должны комплементарно соответствовать друг другу. Значит, только по совпадению кодов в живой системе может осуществляться контроль передачи и приёма молекулярной биологической информации. После информационного этапа следует этап управляющих воздействий, когда вступают в действие электронно-конформационные механизмы фермент-субстратного комплекса и идёт управляемый акт химической реакции. Комплементарный стереохимический контакт управляющих и сигнальных фермент-субстратных кодовых компонентов является достаточной формой воздействия на исполнительные органы фермента. Этот контакт выполняет роль “электронного ключа”, который запускает электронно-конформационные механизмы аппарата химического катализа фермента. Таким способом осуществляется передача управляющей информации исполнительным органам макромолекул. По всей вероятности, это и есть те, пока недостающие и разыскиваемые фрагменты информационного управления, указывающие на единство процессов управления и информации в каждой живой клетке. Важно также подчеркнуть, что адресный код и код операции являются пространственными комбинационными кодами, поэтому в результате стереохимического кодирования у разных классов ферментов они принимают только своё смысловое значение. Этим, видимо, и объясняется высокая специфичность действия различных ферментов. Однако следует отметить, что более распространённым форматом команды управления являются не одноадресные, а двухадресные команды, когда активный центр фермента содержит код операции и адреса двух разных молекул субстрата, подлежащих преобразованию. К этому типу относится большинство известных управляемых химиче-ских реакций и, прежде всего, реакции переноса атомных химических групп. Имеются также трёхадресные и иные форматы команд управления, когда активный центр фермента содержит дополнительные адресные кодовые группы, например, для связывания с коферментом, с АТФ и т. д. На принципах различных вариантов адресации формируются и другие информационные сигналы, которые, как правило, размещены в углублениях или на поверхностных участках белковой молекулы. Эти сигналы обычно размещены в виде отдельных локальных или поверхностных кодовых биохимических матриц, которые служат для информационной коммуникации белка с другими биологическими молекулами клетки.
3. Информационная концепция фермент-субстратных взаимодействий. Длинная полипептидная цепь при построении любого белка оказывается как бы застёгнутой между отдельными аминокислотами и фрагментами цепи по матричному комплементарному типу. При этом относительно слабые многоточечные информационные взаимодействия, обусловленные многочисленнными боковыми R-группами аминокислот, становятся с одной стороны, достаточно прочными для стабилизации нативной конформации белка, а с другой – достаточно лабильными для участия их в формировании биологических функций. Оказавшиеся на поверхностных участках многочисленные боковые R-группы организуются в локальные или поверхностные биохимические матрицы, которые служат для информационной коммуникации белка с другими молекулами клетки. Белок строится на основе генетического кода, с использованием химических и стереохимических принципов записи информации, а это уже является достаточным условием для того, чтобы предложить и рассмотреть в данной статье информационную концепцию функционального поведения белковой молекулы. Ясно, что белки клетки, как носители информации в виде многочисленных элементарных сигналов боковых групп, являются уже не столько средствами хранения этой информации, сколько средствами её реализации и воплощения. В различных ситуациях связующим звеном между управляющей системой и управляемым процессом в живой клетке служат рецепторы информации – активные центры (или другие коммуникационные сигналы) и исполнительные органы и механизмы ферментов или других функциональных белков. Работа биологических рецепторов только в некоторой степени напоминает работу датчиков информации, которые используются в технических системах. Биологические рецепторы, например, ферментов сами осуществляют поиск, приём и рецепцию субстратной информации, что, по своей сути, является актом запрограммированного поиска объекта управления (молекулы субстрата), с “запросом” его информации. Нативная макромолекула белка вне информационного воздействия находится в исходном равновесном состоянии. Каталитический центр фермента становится активным и готовым к выполнению команды управления лишь с момента рецепции молекулы подлинного субстрата. Рецепция информации осуществляется активным центром фермента за счет полного соответствия его адресного и каталитического кодов химическим кодовым группам субстрата, и благодаря их комплементарным физико-химическим, стерическим и слабым энергетическим взаимодействиям – электростатическим, гидрофобным, водородным, вандерваальсовым и др. А для того, чтобы эти силы могли возникнуть и действовать необходимо, прежде всего, стерическое, пространственное соответствие. Как считают биологи, субстрат присоединяется к активному центру фермента, который геометрически и химически представляет собой как бы негативный отпечаток молекулы субстрата, то есть – комплементарен ей. А с информационной точки зрения – это процесс рецепции кодовых компонентов и проверка их на функциональное соответствие друг другу. Поэтому рецепция и приём осведомляющей кодовой информации субстрата заканчивается подключением его молекулы, через контакт “устройства комплементарного сопряжения” активного центра, к управляющим органам и механизмам фермента. В связи с этим, взаимодействие и контакт реагирующих белков и молекул в живой системе является событием информационным, генетически обусловленным, а не случайным как, например, при взаимодействии молекул в чисто химической реакции. Таким образом, фермент-субстратные взаимодействия можно представить в виде информационной модели, основанной на стереохимических принципах и правилах молекулярной биохимической логики. Ферменты обладают своей программой “осязательного” распознавания кодовых компонентов молекул субстрата, которые комплементарны по химическим и стерическим (геометрическим) характеристикам их активному центру. Адресный код и код операции каждого типового фермента имеет свой элементарный состав и индивидуальное пространственное расположение боковых атомных группировок в активном центре, поэтому изучение стереохимических кодов белковых молекул является одной из многих задач молекулярной биологической информатики. Процесс рецепции информации подлинного субстрата, осуществляемый активным центром фермента, вызывает конформационные изменения в фермент-субстратном комплексе, при которых кодовые химические группы фермента и молекулы субстрата занимают самое оптимальное положение для прохождения каталитической операции. Важно отметить, что подключение объекта управления (молекулы субстрата), через кодовый стереохимический контакт комплементарного сопряжения, ведёт к индукции электронно-конформационного возбуждения фермент-субстратного комплекса. Присоединение подлинного субстрата сначала ведёт к переброске электронов и протонов между ферментом и молекулой субстрата, усилению электронной перестройки вдоль сопряженной системы связей, что соответственно приводит к возбуждению фермент-субстратного комплекса и, как итог, благодаря подвижным водородным связям, ведёт к динамическим конформационным сдвигам и срабатыванию “силового молекулярного привода” аппарата химического катализа фермента. Эти механизмы обеспечивают ферменту не только химическую, но и динамическую реактивность и, как результат, – автоматический режим его работы. Возникшие конформационные изменения в фермент-субстратном комплексе сопровождаются разрывом или образованием химических связей субстрата, которые происходят с высвобождением или затратой энергии. В случае необходимости эти процессы поддерживаются химической энергией в форме АТФ. Быстрому протеканию ферментативной реакции способствует высокая химическая и динамическая реактивность фермента. Высокая химическая реактивность обеспечивается режимом полифункционального катализа, когда на превращаемую химическую связь субстрата одновременно действует стереохимическая комбинация различных каталитически активных химических группировок активного центра (код операции) фермента. Интересным фактом здесь является то, что белковые молекулы стереохимическим способом решает сразу две задачи, – информационной коммуникации и полифункционального катализа. Динамическая реактивность фермента, при взаимодействии фермента с субстратом, создаёт напряжение, то есть ориентирует и фиксирует взаимодействующие химические группы таким образом, что это создаёт механическую составляющую, которая снижает энергию активации и способствует эффективному прохождению реакции. Можно считать, что, в рамках сделанных допущений, информационная модель описывает процесс управления химической реакции, ведущий к образованию продуктов реакции. Образование продуктов реакции сопровождается нарушением их физико-химического соответствия управляющим кодовым компонентам фермента, а это приводит к возврату фермента в исходное состояние. Фермент, как взведённая пружина, возвращаясь в исходное состояние, способствует выбросу продуктов реакции из активного центра. Этап фермент-субстратного взаимодействия является заключительным фрагментом биокибернетического управления, указывающим на единство процессов управления и информации в живой системе. Заметим также, что клеточная система сразу же получает информацию о ходе управляемых процессов в виде стереохимических кодов продуктов реакции, которые становятся субстратами для других ферментов или выступают в роли молекул обратной связи. Сигнальная (осведомляющая) информация субстратов служит для информирования управляющей системы о состоянии управляемых объектов, о ходе реакций, об эффективности протекающих процессов и т. д. Отличительной способностью белков клетки является их способность адекватно и сходным образом отвечать на довольно слабые информационные воздействия, достаточно мощными обратимыми конформационными изменениями. В этом, видимо, и заключается основа и сущность их биологической активности. Способность белка индуцированно возбуждаться и адекватно отвечать на сигнальную информацию изменением своей конформации является специфической особенностью. Конформация фермента меняется при взаимодействии его с субстратом, молекула гемоглобина – при соединении с кислородом, конформационные изменения обеспечивают функционирование сократительных белков и т. д. Способность ферментов и других белков клетки автоматически отвечать на слабые информационные воздействия, довольно мощными обратимыми конформационными изменениями, используется клеткой практически для всех биологических функций.
4. Ферменты и белки – это молекулярные биологические автоматы с программным управлением. В живой клетке имеется множество локально рассредоточенных объектов управления (субстратов). Для эффективного управления ими все выходные управляющие аппараты биокибернетической системы клетки должны быть “механизированы и автоматизированы”. Они должны быть снабжены системой адресной доставки, а также обладать свойствами адаптивного управления, в зависимости от наличия объектов управления, регуляторных сигналов обратной связи или физико-химических воздействий окружающей среды. Всеми этими свойствами в достаточной мере обладают ферменты и другие функциональные белки живой клетки, которые являются материальными носителями не только программных средств, но и самих управляющих органов и механизмов. В связи с этим, в молекулярно-биологической системе клетки, в качестве выходного управляющего звена используются белки и ферменты, представляющие собой молекулярные биологические автоматы или манипуляторы с программным управлением [3]. В результате стереохимического кодирования и программирования каждый белок клетки получает своё, как внешнее, так и внутреннее структурно-функциональное и информационно-программное обеспечение. К внешнему обеспечению белков могут относиться: 1) средства информационной коммуникации, – представляющие собой адресные стереохимические коды активных центров, которые состоят из пространственной комбинации аминокислотных остатков с различными R-группами; при помощи таких кодов ферменты способны к адресному поиску, комплементарному взаимодействию и связыванию молекул субстрата; 2) зона химического катализа, представляющая собой код каталитической операции активного центра фермента, который определяет характер химической реакции и состоит из стереохимической комбинации различных боковых R-групп, обладающих высокой химической реактивностью; 3) к средствам коммуникативного “общения” белка с другими молекулами клетки могут также относиться разного рода и назначения локальные или рельефные поверхностные кодовые микроматрицы. Они образованы координатной мозаикой различных, иногда весьма многочисленных боковых R-групп, находящихся в углублениях или на поверхностных участках белковой молекулы. К внутреннему обеспечению белковых молекул могут относиться: 1) средства программного обеспечения, которые скорее неявно, чем в явной форме “загружены и заложены” в аминокислотной “линейной”, а затем, и трёхмерной организации белковой молекулы; 2) средства структурной организации исполнительных органов и механизмов белка, которые обладают высокой динамической реактивностью; 3) энергетические средства макромолекулы, заключенные в её химических ковалентных и нековалентных (слабых) связях, а при необходимости и в дополнительной энергии в форме АТФ. Стереохимические (пространственные) аминокислотные коды, находящиеся в белковых молекулах (и передаваемые по физическим каналам связи), по функциональному назначению могут разделяться на адресные коды, коды операций, регуляторные, информационные, структурные коды и др. Адресный код (или коды), – служит для комплементарной рецепции функционального адреса молекулы (или молекул) субстрата. При помощи адресных кодов определяется класс биохимических соединений (то есть вид молекулярной информации), тип информационного элемента или атомной группы для связывания его с активным центром. Код операции, – указывает характер химической операции (реакции) во время химических превращений. Регуляторные коды, – служат для принятия информационных сигналов обратных связей во время функционирования фермента. Информационные коды – это те локальные или поверхностные рельефные биохимические матрицы, которые служат для связывания белковой макромолекулы с её функциональными молекулярными партнёрами или партнёрами по агрегатированию. Структурные коды – это та кодовая организация химических букв в макромолекуле, которая определяет структурную организацию исполнительных органов и механизмов белковой молекулы. Такое стереохимическое кодовое разделение сигналов позволяет белку динамически и информационно взаимодействовать с различными молекулярными партнёрами: с транспортными молекулами, с коферментами, с мембранами клетки, с АТФ, с регуляторными молекулами, с партнёрами по агрегатированию и т. д. В связи с этим, процесс описания конкретного функционального алгоритма белковой молекулы на языке “стереохимических кодовых команд” можно было бы назвать – “программированием в стереохимических кодах”. Целью стереохимического кодирования белковых макромолекул является передача адресных информационных сообщений с кодовым разделением различных по своему назначению сигналов. Каждый функционально активный белок клетки, как молекулярный биологический программный объект, всегда состоит из данных, то есть, – функциональных биохимических программных элементов (аминокислот) и физико-химических алгоритмов, определяемых биохимической логикой их взаимодействия. При этом динамическая реактивность макромолекулы белка связана с кооперативным изменением сил притяжения и отталкивания, поэтому свободная энергия взаимодействия аминокислот в составе макромолекулы, при информационном контакте с молекулярными партнёрами, и определяет её функциональное поведение. При недостатке энергии белковые молекулы способны адресно (информационно) взаимодействовать с молекулами АТФ, которые в живой клетке выполняют роль аккумулятора химической энергии. Как мы видим, стереохимический язык живой формы материи является не только средством выражения информационных сообщений, но и средством “естественного общения” биологических молекул друг с другом. Основной целью стереохимического кодирования и программирования белковых молекул является: 1) передача в трёхмерных структурах белков различных сообщений со стереохимическим кодовым разделением сигналов; 2) программирование работы молекулярных органов и исполнительных механизмов, определяющих функции белковых молекул; 3) повышение помехоустойчивости информационных сообщений, путём применения комплементарных обратных связей, при взаимодействии биологических молекул друг с другом с помощью их биохимических матриц; 4) повышение достоверности передачи сообщений, так как ошибочное замещение одной аминокислоты на другую в любом стереохимическом коде, как правило, ведёт к “потере” биологического сигнала белковой молекулы; 5) возможность регуляторного воздействия на управляющие стереохимические коды макромолекулы фермента путем “разрешения или запрета” на прохождение управляющих команд (при помощи регуляторных молекул обратных связей); 6) экономное использование различных компартментов, каналов связи и т. д. Таким образом, стереохимический принцип кодирования и программирования функций белковых молекул – это, в первую очередь, и есть тот путь, который непосредственно ведёт от молекулярной информации к биологическим характеристикам живой формы материи. Нам до сих пор неясен и непонятен этот древнейший язык живой природы, который, по всей вероятности, является не только средством молекулярного “общения”, но и формой выражения биологической сущности живой материи. Только таким способом программируется весь путь и биологическая судьба любой активной макромолекулы живой клетки. Поэтому каждый фермент или другой белок клетки становится обладателем своей сложной биологической судьбы и начинает функционировать строго в соответствии с теми обстоятельствами, в которых он находится, и в соответствии с той программой, которая химическим и стереохимическим способом загружена в его линейную и трёхмерную структуру. Многие белки программируются таким образом, чтобы они могли реализовать не только свою управляющую информацию, но и специфически могли воспринимать и реагировать на осведомляющую информацию сигнальных и регуляторных молекул. Таким образом, трёхмерные структуры белков могут обладать своими “входными и выходными” средствами обмена информацией с другими молекулами клетки. К примеру, аллостерический фермент, благодаря выходным управляющим кодовым компонентам активного центра, всегда “знает” с каким объектом управления ему следует взаимодействовать, а благодаря набору входных кодовых компонентов, которые служат для обратной связи, он способен адекватно реагировать на информационные воздействия сигнальных или регуляторных молекул. Следовательно, для того, чтобы логический механизм фермента или другого белка клетки заработал и был способен точно и быстро выполнить все указания генов, в их трёхмерную структуру должны быть заложены как исполнительные органы и механизмы, так и их программное, функциональное, энергетическое и информационное обеспечение. Такое условное подразделение на отдельные средства структурного и информационного обеспечения макромолекул необходимо для понимания информационных принципов и механизмов, лежащих в основе функционального поведения как белковых, так и других биологически активных молекул. Теперь мы знаем, что все эти многоплановые цели и задачи могут выполняться белковыми молекулами только благодаря наличию в их структурах многофункциональных био-логических элементов – аминокислот, комбинационный состав которых, в каждом конкретном случае, определяется генами. Известно, что смысл действия информационных сигналов и сообщений, как правило, сводится к включению или выключению “силовых управляющих органов и механизмов”. В молекулярной биологической системе эти функции обычно выполняются ферментами или другими белками, но, заметим, – только на молекулярном уровне. Здесь управление химическими реакциями осуществляются не только за счет высокой химической реактивности ферментов, но и за счет их высокой реактивности динамической. При этом любая молекула субстрата воспринимается соответствующим ферментом как биологический объект управления, подлежащий химическому и динамическому (механическому) воздействию. А сам объект управления (субстрат), воспринимающий эти воздействия, является “нагрузкой”, как для аппарата химического катализа фермента, так и для его “силового молекулярного привода”. Таким образом, фермент действует на молекулу субстрата с помощью химических, динамических (механических) и информационных средств. А благодаря стереохимической форме представления информации ферменты способны в автоматическом режиме решать ряд биологических задач: 1) динамический поиск молекул субстрата (объектов управления) по их сигнальным информативным структурам; 2) приём осведомляющей информации молекул субстрата и подключение их, через матричный контакт устройства комплементарного сопряжения, к управляющим органам и механизмам фермента; 3) рецепцию кодов осведомляющей информации молекулы (или молекул) субстрата и проверку их на комплементарное соответствие управляющим сигналам – адресному коду и коду операции фермента; 4) запуск силовых молекулярных электронно-конформационных механизмов фермента, через контакт устройства “комплементарного сопряжения” фермента с субстратом. Стереохимический контакт управляющих и сигнальных кодовых компонентов фермента и субстрата является достаточной информационной формой воздействия на исполнительные органы и механизмы фермента. Сдвиги зарядов макромолекулы, во время взаимодействия её с молекулой субстрата, определяют динамическую реактивность фермента и ведут к снижению энергии активации и ускорению прохождения химической реакции, то есть к реализации кода каталитической операции. Таким образом, весь смысл прохождения генетической информации заключается в управлении ферментами различного рода химических реакций или в выполнении белками определённых биологических функций. Поэтому все генетически детерминированные функции управления на расстоянии в клеточной системе выполняются управляющими автоматами, то есть ферментами и белками. Динамическая организация белков включает в себя весь необходимый и достаточный набор информационных, управляющих, программных и энергетических средств, наличие которых указывает на несомненную принадлежность ферментов и других функциональных белков клетки к категории молекулярных биологических автоматов или манипуляторов с гибким программным управлением. Причем ключевые ферменты вполне можно отнести к категории полных автоматов с авторегулированием, так как после окончания рабочего цикла они не только начинают его вновь самостоятельно, но и могут регулировать прохождение химических реакций с помощью сигнальных или регуляторных молекул обратной связи. Известно также, что некоторые ферменты и белки, программно объединяются между собой или с молекулами РНК в агрегатированные автоматы и становятся способными к выполнению сложнейших биологических функций. К молекулярным агрегатам такого рода можно отнести ДНК и РНК-полимеразы, рибосомы, АТФ-синтетазу и т. д. Причем, каждый из этих, иногда довольно сложных аппаратных устройств, приспособлен выполнять определённую последовательность команд и био-логических операций, то есть, способен реализовать какие-то алгоритмы биологической деятельности. Поэтому и в данном случае имеются все основания говорить о программировании молекулярных биологических функций. Таким образом, живая клетка сама “проектирует”, создаёт и применяет для дистанционного управления высокоэффективные автоматические молекулярные средства с программным управлением. Только благодаря молекулярным биологическим автоматам, манипуляторам и агрегатам управление всеми клеточными процессами полностью “механизировано и автоматизировано”, информационно скоординировано и осуществляется в полном соответствии с теми генетическими программами, которые перенесены и загружены в их молекулярную структуру. Теперь уже не вызывает сомнений, что причиной упорядоченной организации живой материи является системная организация и высокая информационная насыщенность взаимодействующих биологических молекул, несущих как управляющую информацию – адресные и функциональные коды белков и ферментов, так и сигнальную осведомляющую – химические коды субстратов. Информационная молекулярно-биологическая система самоуправления клетки – это комплекс различных молекулярных управляющих устройств и средств, который, с одной стороны, осуществляет управление различными химическими процессами и биологическими функциями, а с другой – занимается реорганизацией и реконструкцией своих же биологических структур и компонентов. Поэтому ферменты и другие функциональные белки используются клеткой в качестве выходного управляющего звена её биокибернетической системы. И, действительно, только в клеточных условиях ферменты способны повышать “скорости катализируемых ими реакций в 108 – 1020 раз. А число оборотов наиболее активных ферментов достигает 36 000 000 в 1мин. Такое число молекул субстрата, претерпевает превращение за 1 минуту в расчете на одну молекулу фермента” [4]. Заметим, что такую непревзойденную производительность и избирательность, на наш взгляд, могут развивать и вырабатывать только лишь молекулярные биологические автоматы с программным управлением. Ясно, что подобные процессы не могут обеспечиваться химическими катализаторами, какими бы уникальными и замечательными свойствами они не обладали. Работу ферментов, как организаторов всех химических процессов живой клетки, нельзя определять только одним, хотя и существенным их свойством. Поэтому называть ферменты биокатализаторами химических процессов, с точки зрения сегодняшнего дня, более чем несовременно. Феномен био-логического управления по силам лишь молекулярным биологическим автоматам и манипуляторам. А полифункциональный катализ, используемый молекулярными биологическими автоматами (ферментами), применяется лишь как способ управления химическими превращениями. Однако избирательная химическая и динамическая реактивность фермента может осуществляться только информационным путём. В связи с этим, все белковые молекулы представляют собой не только потоки биоорганического вещества, но они же образуют и информационные потоки и сети, контролирующие различные биохимические и молекулярные функции живой клетки (организма). Программирование этих потоков и сетей обеспечивается экспрессией десятков и сотен различных генов, объединённых между собой скоординированными управляющими и регуляторными воздействиями. А если учесть, что различные ферментативные системы, состоящие порой из десятков и сотен ферментов, участвуют в организации множества различных последовательностей идущих друг за другом химических реакций, которые в совокупности составляют клеточный метаболизм, то можно констатировать, что управление химическими процессами и биологическими функциями клетки осуществляется молекулярными информационными потоками и сетями “автоматизированного” управления. Разные классы биомолекул выполняют различные специфические функции, которые основаны на применении своих биохимических элементов и своей структурно-функциональной информации. Так или иначе, генетическая информация, проникая в биологическую структуру через её элементарный состав, переносит туда и весь необходимый набор программных, энергетических и функциональных средств, на основе которых живая клетка достигает упорядоченности структур и процессов. В связи с этим все клеточные процессы управляются и взаимно координируются той программной информацией, которая в данное время перенесена и загружена в молекулярную структуру функциональных биологических молекул клетки. Радикально функции клеток могут меняться только при загрузке в её аппаратную часть новой молекулярной информации, то есть уже за счет других синтезированных биологических молекул, и в первую очередь, – белковых макромолекул, включаемых в состав различных молекулярных средств, структур и компонентов, например, в процессах деления или дифференцировки клеток.
Список литературы1. Ю. Я. Калашников. Основы молекулярной биологической информатики. – М., 2004. – 66с. – Депонир. в ВИНИТИ РАН 13.04.04, № 622 – В2004, УДК 577.217: 681.51
2. Ф. Айала, Дж. Кайгер. Современная генетика. Пер. с англ. в 3-х томах – М: Мир, 1988.
3. Ю. Я. Калашников. Ферменты и белки – это молекулярные биологические автоматы с программным управлением. – М., 2002. – 25с. – Депонир. в ВИНИТИ РАН 21.05.02, № 899 – В2002, УДК 577.217: 681.51
4. А. Ленинджер. Основы биохимии. Пер. с англ. в 3-х томах – М: Мир, 1
Похожие работы
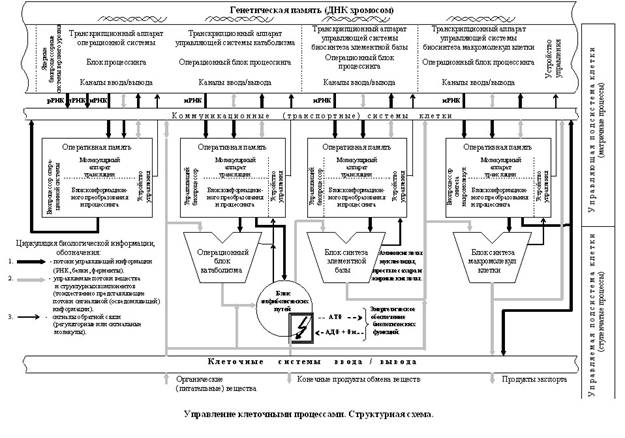
... её структурной и функциональной организации. Систематизируя отдельные информационные фрагменты и известные факты, в чем-то их переосмысливая, автор статьи сконструировал подобную схему и назвал её: “Информационное управление клеточными процессами. Структурная схема”. (Прилагается в конце статьи, с. 27). Структурная схема информационных процессов и сопряженных с ними процессов биохимических должна ...
... прохождения генетической информации и ее управляющий характер. Функции биопроцессорных единиц в управляющей биокибернетической системе клетки жизненно важны и требуют с информационной точки зрения более детального рассмотрения и изучения. По выполняемым функциям генетическая память, молекулярные биопроцессоры и их выходное звено – ферменты и белки являются центральными устройствами клетки, на ...
... зарождения. Это, по всей вероятности, и есть та диктатура информационной субстанции, которая определяет нашу биологическую сущность на самом фундаментальном – молекулярном уровне. 4. Информация правит нашим миром или информационная концепция эволюции. Информация, – она кажется нам нереальной и неопределимой. Необъятный мир её разнообразен и не изучен. Но она не только существует, но даже живёт ...
... полей космоса и окружающего нас мира. Изучение информационных полей живого вещества и сфер биологических макромолекул-кристаллоидов может дать дополнительные сведения о природе и принципах организации живой формы материи. 4. Единство вещества, энергии и информации – основной принцип существования живой формы материи. Вещество, энергия и информация являются важнейшими сущностями нашего мира и ...
0 комментариев