Г.Н. Лысенко (Нежинский госуниверситет, Нежин)
Один из наиболее известных примеров восстановительных смен степной растительно сти - трансформация растительного покрова отделения Украинского степного природного заповедника «Михайловская целина» (Сумская область). Особенно ярко сукцессия проходила на участке с абсолютно заповедным режимом. За более чем полувековую историю наблюдений этот эталон луговых степей полностью лишился сообществ с ярко выраженной эдификаторной ролью типичных степных ценозообразователей - ковылей и типчака (Stipa, Festuca), а также осоки низкой (Carex humilis).
Растительный покров в дозаповедный период был представлен достаточно однообраз ными дигрессивными сообществами с доминированием типчака (овсяницы валисской, Festuca valesiaca), осоки низкой и полевицы виноградниковой (Agrostis vinealis), со значительной примесью степного разнотравья. На местах интенсивных сбоев были широко распространены мхи туидиум и тортула (Thuidium abietinum, Tortula ruralis). В то время наиболее распространенной была разнотрав но-низкоосоково-типчаковая ассоциация (Festuca valesiaca + Carex humilis + разнотравье). На этом фоне выделялись сообщества с доминированием ковылей волосистого (Stipa capillata) и перистого (S. pennata). Высокое ценотическое значение имели мятлик узколистый, овсец опушенный, тимофеевка степная (Poa angustifolia, Helictotrichon pubescens, Phleum phleoides). Красочность степи придавали многочисленные виды разнотравья - шалфей луговой, подмаренник русский, лабазник обыкновенный, мытник хохлатый, клевер горный (Salvia pratensis, Galium ruthenicum, Filipendula vulgaris, Pedicularis comosa, Trifolium montanum) и многие другие. Немногочисленные корневищные злаки - пырей ползучий, вейник наземный и кострец безостый (Elytrigia repens, Calamagrostis epigeios, Bromopsis inermis) - формировали сообщества лишь по днищам балок и ложбин стока. В списке флоры насчитывалось 262 вида сосудистых растений. Именно такой нашли и описали целинную степь, находившую ся в ведении Михайловского конного завода, Е.М. Лавренко и И.Г. Зоз, посетив участок в конце 1920-х гг. (Лавренко, Зоз, 1928).
В 1928 г. «Михайловская целина» была провозглашена заповедником местного значения, а в 1951 г. передана в ведение Академии наук Украины (АН УССР), что стало началом периода эффективной охраны. Между 1928 и 1951 годами хозяйственное использование (выпас и сенокошение в различных комбинациях) не прекращалось, но его интенсивность существенно снизилась. В это время растительные сообщества плакорных участков и пологих склонов приобрели типичные черты богаторазнотравно-типчаково-ковыльных луговых степей.
Вместе с тем, результаты обследования заповедной степи Г.И. Билыком в 1957 г. свидетельствуют о значительном , более чем наполовину, уменьшении площадей под сообществами с доминированием типчака, перистого ковыля (Stipa pennata s. str.) и осочки (Carex humilis), при одновременном широком распространении волосисто-ковыльных сообществ (с доминированием Stipa capillata) (Бiлик, 1957). Однако дерниннозлаковые сообщества все еще преобладали в заповедной степи. Корневищно-злаковые формации с доминированием костреца (Bromopsis inermis) и вейника (Calamagrostis epigeios) уже занимали около четверти территории «Михайловс кой целины» и имели тенденцию к дальнейше му распространению.
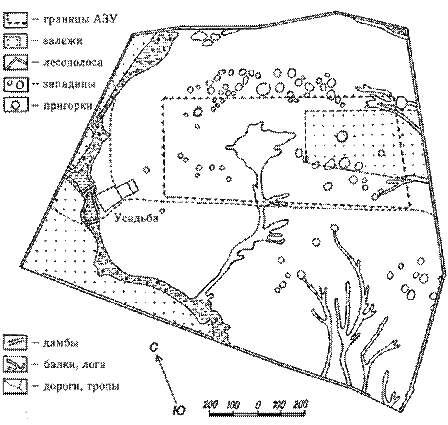
Последнее подтверждается материала ми геоботанического картографирования , проведенного Г.И. Билыком и В.С. Ткаченко в 1971 г. (Бiлик, Ткаченко, 1972, 1973). Тогда растительность заповедника была еще близка к эталонному варианту северных луговых степей. Однако существенное преобладание корневищнозлаковых формаций над дерновиннозлаковым и (соответственно, 61 % и лишь 16 % площади заповедного участка), по мнению Г.И. Билыка, свидетельствовало о продолжающейся мезофитизации растительности.
Отмеченная тенденция саморазвития фитоценозов «Михайловской целины» сохранилась и в последующее десятилетие. Крупномасштабная геоботаническая съемка, осуществленная В.С. Ткаченко в 1981 г. (Ткаченко, 1984), подтвердила выводы Г.И. Билыка. Площади под сообществами с доминированием дерновинных злаков сократились до минимума (до нескольких гектаров), тогда как корневищно-злаковые сообщества начали трансформироваться в кустарниковые степи с эдификаторной ролью ракитника русского (Chamaecytisus ruthenicus), ранее не наблюдавшиеся на территории заповедной степи. Следует отметить, что указанные процессы развивались на фоне вначале четырехлетней, а затем трехлетней сенокосной ротации (то есть каждый участок выкашивался раз в 4 или 3 года). Как оказалось, ракитник весьма отзывчив на удаление надземной биомассы: оно провоцирует у этого кустарника мощный всплеск побегообразования, как правило, влекущий за собой ускорение его экспансии. К сожалению, попытки регулировать сообщества с помощью сенокошения оказались неэффективными - они не помогли сохранить типичные виды зональных луговых степей, для чего, собственно, и был организован этот заповедник.
Очередное обследование «Михайловской целины», проведенное еще десятилетие спустя (Ткаченко и др., 1993), свидетельствовало об углублении процесса мезофитизации растительного покрова. Этот процесс продолжался, несмотря на усиление режима сенокошения (произошел переход на двухлетнюю ротацию).
Тогда же мы отметили специфику развития растительности абсолютно заповедного участка (АЗУ). Какие-либо воздействия на этот участок были полностью исключены с 1956 г. К 1991 г. значительные площади АЗУ заняли разнотравные сообщества с доминированием крапивы (Urtica dioica) и бодяка (Cirsium arvense), приуроченные к обширному понижению, занимающему центральную часть абсолютно заповедного участка. Одновременно с вышеописанными процессами продолжалось разрастание степных кустарников, в основном представленных зарослями терна (Prunus stepposa), которые за время полного невмешательства образовали мощные куртины. Кроме того, на территории АЗУ, впрочем как и в косимой степи, значительно увеличилось число одиночно стоящих деревьев дикой груши и яблони (Pyrus communis, Malus sylvestris) при стабильном количестве сосны (Pinus sylvestris), появившейся на территории заповедника в 1956 г.
Вследствие ежегодного сенокошения площадь под кустарниковой степью сократилась, так как оказалось, что такая частота скашивания сильно угнетает развитие ракитника русского. Однако уплотнение сенокосной ротации (кошение через год) не остановило углубление процесса мезофитизации. Поэтому площади под корневищнозлаковыми формациями продолжали увеличиваться, тогда как дерновиннозлаковые фитоценозы практически полностью исчезли с территории заповедника.
Ценотическое значение растительных сообществ с типчаком ограничивается их содоминированием с мятликом (Poa angustifolia) и райграсом высоким (Arrhenatherum elatius), поскольку такие, некогда типичные содоминанты, как тырса и осочка, стали очень редкими компонентами сообществ. Значительное распространение луговых сообществ с доминированием райграса вывело луга в «Михайловской целине» на лидирующие позиции.
Мы разделяем мнение А.М. Семеновой-Тян-Шанс кой (1977), что подобные изменения растительного покрова луговой степи спровоцированы существующим режимом заповедания. В результате наших исследований оказалось, что показатели влажности и кислотности почв, а также содержания в почве минерального азота на косимом участке и АЗУ статистичес ки достоверно отличаются. Величины этих параметров, характеризующие экотопы АЗУ, оказались значитель но ближе к лесным местообитаниям, чем к степным или лугово-степным, тогда как косимые участки степи еще сохраняют эталонные качества луговых степей. Углубляющиеся процессы мезофитизации травостоя заповедного участка требовали коррекции сенокосно го режима. Поэтому, основываясь на экспериментальных исследованиях влияния различных сроков кошения на состав и структуру фитоценозов, применявшаяся с 1989 г. двухлетняя сенокосная ротация в 1998 г. была заменена на ежегодное сенокошение с пропуском каждого пятого года.
Однако результаты очередного обследования «Михайловской целины» в 2001 г. (Ткаченко и др., 2003) свидетельствовали о дальнейшем углублении процессов олуговения травостоя. Господствующими оставались корневищно-злаковые сообщества. Но теперь они были представлены преимущественно луговыми и лугово-степными фитоценозами, среди которых доминировали райграсовые сообщества формации Arrhenathereta elatii, распространившейся из посевов многолетних трав, характерных для охранной зоны заповедника. Сообщества дерновинных злаков, особенно с доминированием типчака, внешне незаметны на территории заповедного участка. Лишь редкие дернины ковылей перистого и волосатика выделялись на фоне косимой степи во время цветения. На АЗУ продолжалась экспансия крапивы (ею занято уже более четверти площади абсолютно заповедного участка), фитоценозы которой приурочены не только к депрессиям рельефа (днище водосборной котловины, находящейся в центре АЗУ), но и к плакорным участкам, которые ранее были заняты сообществами с доминированием пырея, костреца и вейника (формации Elytrigieta repentis, Bromopsideta inermis, Calamagrostideta epigeioris).
Типичные степные виды подзоны луговых степей, особенно эдификаторы, практически исчезли с территории заповедника, что, в свою очередь, привело к изменению структуры травостоев. Вместе со сменой доминантов в травостоях виды лугово-степного и степного разнотравья (Adonis vernalis, Galium ruthenicum, Linum austriacum, Filipendula vulgaris, Salvia nutans и др.) заменились на типичные луговые, опушечные и даже лесные виды (Potentilla alba, Polygonatum multiflorum, Ophioglossum vulgatum и многие другие).
Изменилась и вертикальная структура травостоев. В большинстве растительных группировок теперь сложно идентифицировать ярусы, характерные для степной растительности. Е.М. Лавренко и И.Г. Зоз (см. выше) упоминают о моховом покрове с примесью слоевищ сине-зеленой водоросли Stratonostoc, свойственной более южным степям и полупустыням. Но в настоящее время моховой покров встречается здесь лишь изредка и, как правило, приурочен к дорогам, тропам, выпасаемым участкам, площадь которых не превышает 5 га. Не выражен и ярус дерновинных злаков и осочки с сопутствующими видами (Carex praecox, Trifolium montanum, Fragaria viridis, Plantago lanceolata, P. stepposa, Ajuga genevensis, Viola hirta, Thymus marschallianus и др.), хотя он был характерным до введения заповедного режима.
Размывание ярусной структуры объясняется наличием толстого слоя ветоши - он не только изменяет гидротермический режим почвы, но и выступает механическим препятствием, мешающим возобновлению степных видов. К тому же нынешние доминанты (ракитник, крапива и луговые травы - райграс высокий, пырей ползучий, вейник наземный) не только определяют физиономические черты сообществ, но и формируют свойственный только им микроклимат, особенно освещенность под пологом травостоя, делая условия непригодными для типичных степных видов. В связи с этим уменьшается видовое богатство растительных сообществ, что наиболее ярко иллюстрирует формация крапивы (Urticeta dioici). Хотя, по мере распространения данной формации, ее видовое богатство с 1991 по 2001 г. увеличилось с 88 до 100 видов, видовая насыщенность (число видов на единицу площади) уменьшилась с 18 до 16 видов (Ткаченко и др., 2003). Причем нередки практически монодоминантные заросли крапивы (не более 5-6 сопутствующих видов) с общим проективным покрытием не менее 80-100 %.
Возник дисбаланс в сезонном развитии травостоев выпасаемых, выкашиваемых и абсолютно заповедного участков. Ранней весной наличие мощного слоя ветоши на АЗУ тормозит развитие эфемеров и эфемероидов. Проведенные нами измерения температуры почвы на участках с различными режимами позволили объяснить 12-ти дневную разницу в зацветании брандушки и стародубки (Bulbocodium versicolor и Adonis vernalis). Средняя температура корнеобитаемого слоя почвы на косимом участке была выше, чем на АЗУ - в зависимости от глубины, для слоя 30-40 см на 0,6°С, для слоя 0-10 см на 3,8°С. Отличия статистически достоверны.
Таким образом, существующие режимы охраны, а вернее режимы использования, не позволяют противостоять направленному изменению основных ценотических структур растительности заповедника. Сенокошение, даже ежегодное, не позволяет удалить весь объем биомассы растений, который в конце вегетационного сезона трансформируется в массу отмершей органики, а влияние последней на степь было отмечено еще в классической работе А.М. Семеновой-Тян-Шанской (1977).
В большинстве случаев неэффективность сенокошения объясняется несовершенством используемой сельскохозяйственной техники. Проблема в том, что вследствие неровного микрорельефа - наличия микропонижений, слепышин, кротовин и т.п. - сложно регулировать высоту скашивания. Поэтому, как правило, рабочие части косилки настраивают несколько выше требуемой оптимальной высоты скашивания, что значительно уменьшает эффект данного регуляционного мероприятия. Кроме того, в степных заповедниках запрещено кошение отавы. Использование этого агротехнического приема значительно усилило бы влияние сенокошения, особенно в лесостепной зоне.
Наблюдавши йся нами слой ветоши (местами на АЗУ он достигает 15-20 см) препятствует семенному возобновлению многих видов, давая преимущество вегетативно подвижным растениям - в данном случае, длиннокорневищным злакам (пыреи, кострецы, вейники) и крапиве двудомной. Изъятие излишков биомассы растений могли бы осуществлять дикие копытные, но в современных условиях об их роли говорить не приходится. Миниатюрная площадь «Михайловской целины» (202,4 га) еще каким-то образом может прокормить небольшое стадо копытных, но не в состоянии предоставить убежище и обеспечить свободу сезонных миграций мало-мальс ки крупной группе животных. За последние полтора десятилетия наблюдений на территории заповедника несколько сезонов находилась пара косуль с приплодом и отмечались одиночные особи лося в период весенне-осен них миграций.
Впрочем, судя по рассказам старожилов и немногочисленным литературным источникам (Ширяев, 1907; Залесский, 1914), дикие копытные не играли существенной роли в регуляции лугово-степных фитоценозов в течение, по крайней мере, последних ста лет. Район «Михайловской целины» очень интенсивно использовался в сельскохозяйственном отношении. На протяжении вегетационного периода практиковался выпас лошадей, крупного рогатого скота, основной июньский сенокос и покос отавы в конце августа. Зимой нередко осуществляли тебеневку. В случае, если таковой не проводили, практиковали ранневесен ний пал. Вследствие такой антропогенной нагрузки фитоценозы «Михайловской целины» перед заповеданием представляли собой достаточно сильные сбои с преобладанием дигрессивных типчатников, очень стойких к стравливанию.
Часть растительной биомассы изымается насекомыми-фитофагами. Но об их роли в трансформации первичной продукции степных экосистем Украины существуют лишь отрывочные сведения. К огромному сожалению, наши энтомологи продолжают заниматься инвентаризацией энтомофауны, не уделяя достаточного внимания изучению роли насекомых, особенно фитофагов, в степных экосистемах. Кроме того, отсутствуют сведения о специфике почвообразовательных процессов на участках с различными режимами заповедания, особенно на АЗУ, где решающее значение могут иметь особенности функционирова ния детритной пищевой цепи. Слабо изучены процессы минерализации органического вещества и роль редуцентов в завершении биогеохимических циклов даже для основных биогенных элементов.
Все вышеизложенное прямо указывает на неэффективность существующих режимов охраны и воспроизводства степных экосистем в зоне лесостепи. А ведь именно здесь существует реальная угроза полной утраты лугово-степных ландшафтных комплексов, сформированных на водораздельных пространствах. Об этом, в свое время, упоминал еще В.В. Осычнюк (Осичнюк, 1979). Как оказалось, режим абсолютного заповедания полезен лишь на первых этапах восстановительной сукцессии, когда действительно требуется полностью исключить все виды антропогенного влияния. Однако по мере хода сукцессии из-за неполноты комплекса биоценотических структур нарушается динамическое равновесие между функциональными блоками биоценозов. И в этот период абсолютно заповедный режим, вопреки ожиданиям, из блага превращается в дестабилизирующий фактор.
По мнению В.С. Ткаченко (2004), «руководствуясь принципом эволюционной адаптации степных экосистем к комплексному воздействию ряда экзогенных факторов, профилирующих «типичные» зональные фитоценоструктуры в прошлом (выпас, палы, сенокошения), необходимо именно их включать в регуляционный комплекс, … который базируется на постоянном контроле состояния экосистем, анализах длительных и временных изменений биоценотического разнообразия и прогнозах. Обязательной составляющей такого контроля должен быть фитоценотический моноторинг».
По нашему глубокому убеждению, лугово-степные экосистемы можно сохранить, лишь разработав индивидуальную «технологическую» схему для каждого конкретного степного участка. Такая схема должна быть основана на результатах комплексных исследований и экспериментов по сенокошению, выпасу, палам и их комбинациям.
Промедление может привести не только к исчезновению зональных степных и лугово-степных видов и связанных с ними фитоценотических структур. Существует риск критического изменения значений ряда лимитирующих экологических факторов, прежде всего - эдафических, что сделает возврат лугово-степных экосистем к эталонному «типичному» состоянию невозможным даже теоретически. В этом случае заповедник уже не сможет решать стоящие перед ним задачи по сохранению видового и ценотического разнообразия, будет утрачен единственный участок целинной луговой степи в Украине.
К огромному сожалению, проведение упомянутых выше экспериментов начато лишь в последнее время. Однако уже получены предварительные результаты по влиянию выпаса лошадей на изменение растительного покрова разнотравно-типчаково-ковыльной степи (отделение Украинского степного природного заповедника «Хомутовская степь», Донецкая область). Расширяются эксперименты по контролируемым палам. Но решить проблему сохранения степных экосистем на современном этапе невозможно без согласования действий не только ученых, представляющих различные отрасли науки, но и представителей природоохранных служб, главнейшей задачей которых является координация действий всех структур, принимающих участие в природоохран ном процессе.
Список литературыБiлик Г.I. Рослиннiсть заповiдника «Михайлiвська цiлина» та її змiни пiд впливом господарської дiяльностi людини // Укр. ботан. журн., 1957. 14, № 4. С. 26-39.
Бiлик Г.I., Ткаченко В.С. Сучасний стан рослинного покриву заповiдника «Михайлiвська цiлина» на Сумщинi // Укр. ботан. журн., 1972. 29, № 6. С. 696-702.
Бiлик Г.I., Ткаченко В.С. Змiни рослинного покриву степу Михайлiвська цiлина на Сумщинi залежно вiд режиму заповiдностi // Укр. ботан. журн., 1973. 30, № 1. С. 89-95.
Залесский К.М. Первые сведения о флоре Сумского уезда Харьковской губернии: Список растений, собранных и наблюдаемых в Сумском, отчасти Лебединском и Ахтырском уездах // Тр. Об-ва испытателей природы Харьковского ун-та. Харьков, 1914. 47, № 1. С. 101-147.
Лавренко Є., Зоз I. Рослиннiсть цiлини Михайлiвського кiнного заводу (кол. Капнiста) Сумської округи // Охорона п'яток природи на Українi. Харкiв, 1928. С. 23-36.
Осичнюк В.В. Деякi особливостi заповiдного режиму у вiддiленнях Українського державного степового заповiдника // Укр. ботан. журн., 1979. 36, № 4. С. 347-352.
Семенова-Тян-Шанская А.М. Накопление и роль подстилки в травяных сообществах. Л.: Наука, 1977. 191 с.
Ткаченко В.С. О природе луговой степи заповедника «Михайловской целины» и прогноз развития ее в условиях заповедности // Ботан. журн., 1984. 69, № 4. С. 448-457.
Ткаченко В.С. Фiтоценотичний монiторинг резерватних сукцесiй в Українському степовому природному заповiднику. Киев: Фiтосоцiоцентр, 2004. 184 с.
Ткаченко В.С., Генов А.П., Лисенко Г.М. Структура рослинностi заповiдного степу «Михайлiвська цiлина» за даними крупно масштабного картування у 1991 р. // Укр. ботан. журн., 1993. 50, № 4. С. 5-15.
Ткаченко В.С., Генов А.П., Лисенко Г.М. Структурнi змiни в рослинному покривi заповiдного лучного степу «Михайлiвська цiлина» за даними великомасштаб ного картування у 2001 р. // Вiстi Бiосферного заповiдника «Асканiя-Нова», 2003. Т. 5. С. 7-17.
Ширяев Г.И. Материалы для флоры Лебединского уезда Харьковской губ. // Тр. Об-ва испытателей природы при Харьковском ун-те. Харьков, (1905-1906) 1907. 40, № 2. С. 233-2
Похожие работы
... : – консолидация предприятий, организаций и учреждений, занятых обслуживанием туристов; – расширение спектра регионального турпродукта; – организация акций и мероприятий, содействующих развитию туризма в Курской области (выпуск проспектов, буклетов, появление информации о регионе в Интернете и туристских газетах и журналах; различные ярмарки и концертные программы); ...
... калия повышенное или высокое составляет от 20,0-30,0 мг на 100 г почвы (средняя обеспеченность) и 30,0-40,0 мг на 100 г почвы (повышенная обеспеченность). 5. Обоснование выбора почвенных участков, пригодных для орошения Пригодность территории под орошение определяется совокупностью целого ряда местных условий, из которых важнейшим является почвенно-мелиоаративная обстановка. В целом при ...
... и ''ядом замедленного действия". Причина тому проста - находящиеся в ней вредные химические элементы очень хорошо усваиваются человеческим организмом. Из года в год в области остается острой проблема качества питьевой воды. Курская область входит в число 32 территорий России, хозяйственно-питьевое водоснабжение в которых осуществляется за счет запасов подземных вод с помощью водозаборных скважин, ...
... сортов яблони, 80 - груши, 35 - сливы, 40 сортов вишни. В конце 1966 года начата закладка карликового сада. [28] Глава 4. Современное состояние особо охраняемых природных территорий г. Ставрополя Много лет Ставрополь держал марку самого зеленого города в стране. Скверы, бульвары, озелененные с двух сторон улицы, сады на приусадебных участках, широкие зеленые газоны и масса цветочных клумб ...
0 комментариев