Владимир Николаевич Анисимов, д.м.н., проф., рук. отд. канцерогенеза и онкогеронтологии НИИ онкологии им.Н.Н.Петрова, СПб
Перспективы биологии: от ложного знания - к истинному невежеству.
В.Я.Александров
За последние 160 лет ожидаемая продолжительность жизни в экономически развитых странах постоянно увеличивалась со средней скоростью 3 мес/год, и нет никаких оснований полагать, что эта тенденция изменится в ближайшее время [1]. Более того, стремительно растет число лиц, проживших 100 и более лет. В некоторых странах Европы их количество удваивается каждые 10 лет. Растет число долгожителей и в России. Например, в Санкт-Петербурге в 1979 г. было 92 жителя, перешагнувших 100-летний рубеж, в 1996 г. - 150, а в 2001 г. - уже 369 [2]. По данным ООН, наиболее быстро растущий сегмент старческой популяции - лица 80 лет и старше. Такое глобальное постарение населения сопряжено с проблемой качества жизни в пожилом возрасте. Все это определяет возросший интерес к геронтологии, и прежде всего - к изучению первичных механизмов старения организмов и популяций, а также факторов, определяющих продолжительность жизни.
До настоящего времени обсуждается вопрос, можно ли разделить нормальное, или физиологическое старение (без болезней) и патологическое старение, непосредственно связанное с такими заболеваниями, как рак, болезни сердца и сосудов, остеопороз, остеоартрит, сахарный диабет и некоторыми нейродегенеративными заболеваниями (болезнью Альцгеймера, болезнью Паркинсона и др.) *.
* Термины, используемые для описания старения, довольно неточны, поскольку нет общепринятого определения самого процесса. В англоязычной литературе имеются два термина - aging и senescence, и хотя оба переводятся на русский язык одним словом старение, но в отечественной литературе aging обычно соответствует нормальному, или физиологическому старению, а senescence определяют как патологическое старение.
Этой проблемой уже более 40 лет занимается Балтиморский проект по старению (Baltimore Longitudinal Study on Aging, BCLA) Национального института старения США, в рамках которого ведутся наблюдения за группами здоровых людей на протяжении их жизни. Поскольку возрастная динамика показателей весьма варьирует от человека к человеку, практически невозможно оценить степень старения отдельного индивидуума, основываясь на измерении одной или нескольких биохимических, физиологических или физических характеристик. Поэтому до сих пор и нет универсальных тестов для определения биологического возраста, сопоставимого по надежности с хронологическим.
Благодаря работе BCLA выяснилось, что большинство измеряемых показателей меняется с возрастом постепенно, тогда как скачкообразные изменения скорее характерны для ассоциированной с возрастом патологии. Различают заболевания, ассоциированные со старением (aging-dependent), или с возрастом (age-dependent). Так, некоторые генетически детерминированные заболевания (например, болезнь Хантингтона) зависят от возраста, поскольку проявляются в предсказуемые годы. Изменения, связанные с нормальным старением, могут играть существенную роль в развитии той или иной патологии. Поэтому весьма важно отличать непатологические возрастные изменения (например, поседение волос) от способствующих развитию одного или нескольких патологических процессов (например, накоплению оксидативных повреждений) и вызывающих или указывающих на заболевание (например, образование амилоидных бляшек в мозге как фактор риска болезни Альцгеймера). Такое разграничение необходимо для выбора профилактических мер против преждевременного старения и возрастной патологии.
В короткой журнальной статье невозможно осветить даже бегло основные направления исследований современной фундаментальной геронтологии, поэтому отошлем читателя к соответствующей литературе [3, 4]. Мы коснемся лишь наиболее “горячих точек” в этой области, которая, по мнению журнала "Science", станет в ближайшие 25 лет одной из наиболее приоритетных научных дисциплин.
Поиск генов старения и долголетия
| Как сумел ты так долго прожить, борода? Как ты дожил до этаких лет? Льюис Кэррол. Алиса в Зазеркалье |
Пpодолжительность жизни животных и человека существенно pазличается у pазных видов. Этот показатель варьирует в 1 млн раз среди всех видов и от 10 до 50 раз внутри групп с одинаковым уровнем организации. Cреди млекопитающих рекорд долгожительства принадлежит одной из пород китов (свыше 200 лет), лабораторные грызуны живут не более двух-трех лет, а многие другие грызуны такого же размера - до 5-10 лет и более. Поскольку видовая максимальная продолжительность жизни коррелирует со скоростью развития, есть основания полагать, что имеются сходные механизмы развития и старения, которые видоспецифичны и генетически запрограммированы. Генетический код человека и шимпанзе сходен на 98%, однако человек живет до 122 лет, а шимпанзе до 56 лет. Очень небольшое число генов может определять различия между ними в стратегии развития и скорости физиологического старения [5].
Оценить наследуемость долгожительства у человека можно, наблюдая за членами одной семьи, включая пpиемных детей (для учета pоли условий сpеды). Результаты большинства исследований долгожительства у близнецов показывают, что наследуемость продолжительности жизни у человека не превышает 50%. По данным шведских геронтологов, изучавших большую группу близнецов, воспитывавшихся в разных семьях, одна треть изменчивости в общей смертности обусловлена генетическими факторами. На пpодолжительность жизни близнецов могут влиять специфические гены, напpимеp опpеделяющие пpедpасположенность к ожиpению или атеpосклеpозу. Вместе с тем получены данные, свидетельствующие о большей наследуемости долгожительства. Так, потомки столетних имели в четыре раза большую вероятность прожить 85 лет и более, чем дети тех, кто умер до 73 лет [4].
Сегодня довольно распространена точка зрения, согласно которой генетическая программа развития исчерпывается репродуктивным успехом (т.е. рождением потомства), а выживание организма после завершения репродуктивной функции если и опосредовано геномом, то весьма косвенно [4]. Ряд недавних публикаций, касающихся связи между возрастом рождения детей и продолжительностью жизни родителей, привлек к этой проблеме пристальное внимание. Показано, что женщины, прожившие 100 и более лет, в четыре раза чаще рожали детей после 40 лет, чем прожившие не более 73 лет [6]. По мнению авторов, поздняя менопауза может быть фактором, способствующим долголетию.
Анализ данных о количестве детей и возрасте их родителей в семьях британских аристократов выявил, что эти показатели коррелируют с продолжительностью жизни [7]. Среди умерших в молодом возрасте (до 20 лет) две из каждых трех женщин были бездетными, а среди проживших более 80 лет таких было менее трети. Ранние роды и большое число детей негативно сказывались на продолжительности жизни женщины. Возраст первых родов был наименьшим у умерших рано и наибольшим у проживших более 80 лет. Больше шансов дожить до 100 лет имели родившие первенца после 40 лет. Интересно, что и мужья жили дольше, если число произведенных ими детей было не слишком велико (а дрозофилы жили дольше, если им не позволяли размножаться). Продолжительность жизни дочерей больше связана с продолжительностью жизни матери, чем отца; у сыновей эта зависимость значительно менее выражена и не определяется полом родителей.
В последние годы существенный прогресс в изучении генетики старения обусловлен работами на беспозвоночных. Популярной моделью стала нематода (Caenorhabditis еlegans), которая живет около 20 дней. Черви размножаются гермафродитическим самооплодотворением, что обеспечивает формирование генетически однотипных популяций. C помощью химических мутагенов или методов генной инженерии получили червей с большей на 50% средней и в два раза большей максимальной продолжительностью жизни, чем у их предшественников. Наличие таких популяций нематод убедительно свидетельствует о генетическом контроле темпа старения. Идентифицирован ген age-1, модификация или супрессия продукта которого увеличивает продолжительность жизни [4]. Этот ген контролирует целую группу генов, имеющих отношение к продолжительности жизни (daf-2, daf-23, spe-26, clk-1). Так, уменьшение активности гена daf-2 (гомолога гена рецептора инсулина) в два раза увеличивает продолжительность жизни нематод.
Другая хорошая модель для геронтологов - плодовая мушка Drosophila melanogaster. Как и в случае с нематодой, получены мутанты дрозофилы, имеющие разную продолжительность жизни. Особи с увеличенным числом копий генов ферментов супероксиддисмутазы (sod1) и каталазы жили на 20-37% дольше обычных, а мухи с избыточными копиями генов лишь одного из этих ферментов антиокислительной защиты таким эффектом не обладали [8]. Трансгенные дрозофилы с избыточной экспрессией гена sod1 в мотонейронах жили на 40% дольше и были значительно устойчивее к окислительному стрессу, чем мухи, не имевшие этого гена [9].
У дрозофил, как и у нематод, система передачи сигнала от инсулина и инсулинподобного фактора-1 (IGF-1) также регулирует размеры тела и продолжительность жизни (рис.1). Так, мутации в регуляторном участке гена гомолога рецептора инсулина (chico) увеличивали продолжительность жизни мух на 45%. Мухи, гетерозиготные по двум различным мутациям в этом гене, жили на 85% дольше, чем дрозофилы дикого типа [10]. Оба типа мутаций в этом гене также сопровождались усилением активности фермента супероксиддисмутазы.
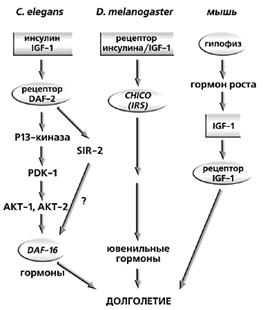
Рис. 1. Схемы передачи сигнала инсулина у нематоды, плодовых мух и млекопитающих.
Хотя параллели между системами регуляции инсулина у нематод и плодовых мух весьма отчетливы, но есть и определенные различия. Так, линия дрозофил c мутацией, названная mth (“мафусаилова” от Methuselach - Мафусаил) не только увеличивала их продолжительность жизни на 35%, но и повышала устойчивость к стрессу, вызванному самыми различными факторами: голоданием, нагреванием или добавлением в корм параквата - генератора активных форм кислорода [11].
С помощью методов генной инженерии получены линии мышей с увеличенной продолжительностью жизни. Гомозиготные мыши с “выключенным” геном рецептора гормона роста (ghr-/-) живут значительно дольше, чем гетерозиготные (ghr+/-) или дикого типа (ghr+/+). У мышей ghr-/-, пропорциональных карликов, замедлен рост, уменьшены длина костей и содержание костных минералов, нет рецептора гормона роста и белка, связывающего его, снижено содержание в крови инсулин-подобного фактора роста I и связывающего его белка-3, а в сыворотке крови увеличена концентрация гормона роста [12].
Карликовые мыши Эймса (мутанты, несущие единичные точечные мутации в Prophet pit-1 гене) живут на 50-64% дольше, чем особи дикого типа. Эта модель - один из первых примеров способности единичного гена продлевать жизнь млекопитающим. У мышей Эйсма снижен уровень пролактина, тиреоидстимулирующего гормона, гормона роста, инсулин-подобного фактора-1 и инсулина в крови, повышена чувствительность к инсулину и понижена температура тела [13]. Как самцы, так и самки карликовых мышей бесплодны и иммунодепрессивны.
У самок мышей Igf1r+/- с частично нокаутированным геном рецептора IGF-1 средняя продолжительность жизни увеличена на 33% по сравнению с самками дикого типа (p < 0.001), а у самцов - лишь на 16%. У этих мышей нет карликовости, а основной обмен, температура тела, потребление корма, физическая активность и фертильность не отличались от контроля, но они более устойчивы к окислительному стрессу по сравнению с контролем (Igf1r+/+) [14].
В течение последних лет ведется интенсивный поиск кандидатов на роль генов смерти и долголетия у человека. В настоящее время принято считать, что для долгожительства человека важен только один ген аполипопротеина Е (AпоЕ). У проживших 100 лет преобладает аллель AпоЕ-E2. Напротив, усиление аллеля E4 располагает к гиперхолестеринемии, коронарной болезни сердца и болезни Альцгеймера (но не к раку или диабету). У лиц старше 90 лет риск болезни Альцгеймера, связанный с аллелем E4, достигает плато. Полагают, что AпоЕ должен рассматриваться скорее как ген “хилости” (frailty), а не как ген долголетия.
Это направление - одно из самых перспективных в современной геронтологии, но оно требует сложного оборудования и больших средств для изучения генетических маркеров старения и ассоциированных с возрастом заболеваний. Однако накопленные к настоящему времени данные позволяют полагать, что нет и, наверное, не будет обнаружен единственный ген, определяющий старение и долголетие как индивидуума, так и вида.
Свободные радикалы и старение
| Ваша теория и солидна, и остроумна. Впрочем, все теории стоят одна другой. М.А.Булгаков. Мастер и Маргарита |
В последние годы наиболее популярна свободноpадикальная теоpия стаpения. Практически одновременно выдвинутая Д.Харманом (1956) и Н.М.Эмануэлем (1958), она объясняет не только механизм старения, но и широкий круг связанных с ним патологических процессов (сердечно-сосудистых заболеваний, возрастной иммунодепрессии, дисфункции мозга, катаракты, рака и некоторых других). Согласно этой теоpии, пpодуциpуемые главным образом в митохондриях активные формы кислорода (О2-., Н2О2, НО·, О2) повpеждают клеточные макpомолекулы. Подсчитано, что за 70 лет жизни человека организм производит около тонны радикалов кислорода, хотя только 2-5% вдыхаемого с воздухом кислорода превращается в его токсические радикалы.
В клетке крысы за день может образоваться до 104 вызванных активными формами кислорода повреждений ДНК, и до 10% молекул белка могут иметь карбонильные модификации. Подавляющее большинство из них нейтрализуется еще до того, как успеют повредить те или иные компоненты клетки. Так, из каждого миллиона супероксидных радикалов от ферментной защиты ускользает не более четырех.
Сегодня показано, что видовая продолжительность жизни коррелирует с активностью фермента супероксиддисмутазы (СОД), содержанием -каротина, -токоферола и мочевой кислоты в сыворотке крови. Так, у долгоживущих линий D.melanogaster активность антиоксидантных ферментов (СОД, каталазы, глютатионредуктазы и ксантиндегидрогеназы) достоверно выше, чем у короткоживующих линий мух [14].
В пользу свободнорадикальной теории старения говорят и эксперименты, в которых трансгенные линии D.melanogaster с дополнительными копиями генов, обеспечивающих избыточную активность СОД и каталазы, жили на 20-37% дольше контрольных мух. Мухи с избыточными копиями генов одного из этих ферментов таким свойством не обладали [8]. Витамин Е, мелатонин, хелатные агенты и некоторые синтетические антиоксиданты увеличивали продолжительность жизни не только дрозофил, но и лабораторных мышей и крыс. Поскольку продукты взаимодействия активных форм кислорода с макромолекулами постоянно обнаруживаются в органах и тканях организма, это означает, что системы антиоксидантной защиты недостаточно эффективны и клетки постоянно испытывают окислительный стресс. Противодействие ему может играть существенную роль в механизме геропротекторного действия эндогенных и экзогенных антиоксидантов.
Меньше калорий - длиннее жизнь
| Диета, которая обильна, опасна, и также опасно избыточное насыщение. Те, кто от природы жирны, ближе к смерти, чем те, которые тощи. Гиппократ. 477-460 гг. до н.э. |
Еще в 30-е годы ХХ в. установили, что диета с существенным (на 40%) ограничением калорий увеличивает на 30-50% максимальную и среднюю продолжительность жизни крыс и мышей. Ограничение пищи увеличивало продолжительность жизни также у рыб, амфибий, дафний, насекомых и других беспозвоночных. В трех больших исследованиях на приматах (главным образом на макаках резус) показано, что некоторые физиологические эффекты низкокалорийной диеты, наблюдаемые у грызунов, воспроизводятся и у обезьян [4]. У них уменьшается уровень глюкозы и инсулина в крови, снижаются температура тела и энергозатраты. Установлено, что геропротекторный эффект голодания определяет общее снижение потребления калорий, а не какого-либо ингредиента пищи.
Подсчитано, что у грызунов при низкокалорийном рационе 80-90% из различных изученных параметров (поведение и обучаемость, иммунный ответ, экспрессия генов, активность ферментов и действие гормонов, толерантность к глюкозе, эффективность репарации ДНК, скорость синтеза белка) проявляло признаки замедленного старения. Такая диета стимулировала апоптоз, который выбраковывает пренеопластические клетки в тканях организма, замедляет накопление мутаций и развитие возрастной патологии.
Пожалуй, основное действие низкокалорийной диеты состоит в ослаблении интенсивности свободнорадикальных процессов. У грызунов при таком содержании замедляется скорость генерации супероксида и Н2О2, уменьшаются окислительные повреждения и падение вязкости мембран; голодание снижает чувствительность тканей in vitro к острому окислительному стрессу. Наибольший защитный в отношении окислительного стресса эффект низкокалорийного питания проявляется в постмитотических клетках головного мозга, сердца и скелетных мышц.
Группа специалистов из США и Италии [15] оценивала эффективность ограниченного питания у людей, соблюдавших низкокалорийную диету в течение шести лет, в сравнении со здоровыми людьми, придерживавшимися обычной американской диеты. 18 испытуемых (возраст от 35 до 82 лет) не курили, не имели хронических заболеваний, не принимали гиполипидемических, антигипертензивных или других лекарств и ежедневно потребляли от 1112 до 1958 кКал. В их рацион входили фрукты, овощи, орехи, злаки, белки и мясо (сладкие напитки, закуски и десерт исключались). 26% калорий они получали из белков, 28% из жиров и 46% из углеводов. Лица контрольной группы (18 человек) потребляли 1976-3537 кКал в день без ограничений в сладком. У испытуемых были меньшая масса тела, уровень общего холестерина, липопротеинов низкой плотности, триглицеридов, глюкозы, инсулина, С-реактивного белка, тромбоцитарного фактора роста АВ (PDGF-AB), систолического и диастолического артериального давления и выше уровень липопротеинов высокой плотности, чем в контрольной группе. Авторы полагают, что низкокалорийная диета существенно снизила риск развития атеросклероза, что подтверждалось уменьшением на 40% толщины интимы медиа артерий. У испытуемых отмечена также меньшая частота воспалительных процессов по сравнению с контрольной группой.
Однако обычный человек вряд ли сумеет на протяжении многих лет строго соблюдать подобную диету. Наиболее известный случай такого эксперимента на себе (при этом не вполне успешного) - опыт известного американского геронтолога и иммунолога Роя Уолфорда. В самые последние годы весьма “горячей точкой” стал поиск миметиков (имитаторов) ограниченной диеты. Предложенный американскими специалистами заменитель сахара 2-дезоксиглюкоза увеличивал продолжительность жизни грызунов, но обладал такими побочными эффектами, что от его использования в качестве геропротектора пришлось отказаться.
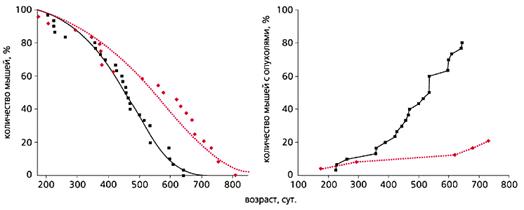
Рис. 2. Влияние фенформина (цветная кривая) на продолжительность жизни
и развитие спонтанных опухолей (справа) у самок мышей С3H/Sn [3].
Еще в 80-годы прошлого века с помощью некоторых фармакологических средств, в частности антидиабетических бигуанидов (повышают чувствительность тканей к инсулину, улучшают толерантность к углеводам, снижают уровень липидов и устраняют явления метаболической иммунодепрессии), также удается продлить жизнь мышей и крыс и снизить у них частоту спонтанных и индуцированных химическими канцерогенами или ионизирующей радиацией новообразований (рис.2). В нашей лаборатории антидиабетический бигуанид метформин увеличивал продолжительность жизни и тормозил развитие рака молочной железы у трансгенных мышей HER-2/neu (рис.3). А в клинике применение метформина снижает риск развития злокачественных новообразований у пациентов, страдающих сахарным диабетом типа 2.
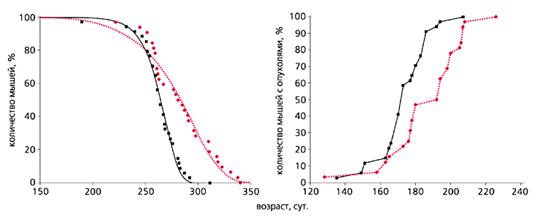
Рис. 3. Влияние метформина (цветная кривая) на продолжительность жизни
и развитие спонтанных опухолей (справа) у самок трансгенных мышей HER-2/neu [18].
Стареющие клетки: хорошие граждане, но плохие соседи
| Не обольщайся призраком покоя: Бывает жизнь обманчива на вид. Настанет час, и утро роковое Твои мечты, сверкая, ослепит. Николай Заболоцкий |
В 1971 г. научный cотрудник Института биохимической физики РАН А.М.Оловников, используя данные о принципах синтеза ДНК в клетках, предложил гипотезу маргинотомии, объясняющую механизм работы счетчика клеточных делений. Согласно его гипотезе, ДНК-полимераза при матричном синтезе полинуклеотидов полностью не воспроизводит линейную матрицу, и реплика всегда получается короче в ее начальной части. Таким образом, при каждом делении клетки ее ДНК укорачивается, что ограничивает пролиферативный потенциал клеток и, очевидно, служит “счетчиком” числа делений и, соответственно, продолжительности жизни клетки в культуре. Открытие в 1985 г. фермента теломеразы, достраивающего укороченную теломеру в половых клетках и клетках опухолей, обеспечивая их бессмертие, стало блестящим подтверждением гипотезы Оловникова.
В последние годы достигнут значительный прогресс в изучении роли теломер в старении. Дисфункция теломер (связана ли с укорочением, прямым повреждением или ассоциированным с теломерой дефектным белком) может приводить к трем последствиям: старению клетки, ее гибели (факторы, подавляющие неопластический процесс) или нестабильности генома, которое может способствовать злокачественной трансформации клетки.
Старение клетки рассматривают как один из ее защитных механизмов при дисфункции теломер, поскольку оно останавливает пролиферацию, тем самым блокируя канцерогенез (“хорошие граждане”) [16]. В течение жизни в организме накапливаются соматические мутации, некоторые из них могут инактивировать гены, участвующие в клеточном старении. Кроме того, потеря гетерозиготности и мутации в генах-супрессорах (р53 и Rb) и онкогенах (например, ras) встречаются даже в нормальных клетках. Другой защитный механизм клетки с дисфункцией теломер, в который вовлечен интактный р53, - апоптоз. В клетках, где накапливаются мутации в гене р53 или в компонентах его регуляции, развивается нестабильность генома, повышающая вероятность злокачественной трансформации. Потеря дистальных областей теломер связана с уменьшением пролиферативной жизни клеток как in vitro, так и in vivo. Анализ данных по укорочению теломер в 15 различных тканях человека показал, что в год теломеры теряют от 20 до 60 пар оснований.
В настоящее время активно развивается гипотеза о важной роли укорочения теломер и соответственно реактивации теломеразы при старении и канцерогенезе. В опухолях человека появление бессмертных клеток (иммортализация) почти всегда обусловлено подавлением гена каталитической субъединицы теломеразы (hTERT). Множественность механизмов, подавляющих или регулирующих активность этого фермента, может объяснить исключительную редкость спонтанной иммортализации нормальных клеток человека. С другой стороны, введение субъединицы hTERT стимулирует рост различных типов клеток человека без последующего развития опухолевого роста. Вероятно, основная функция супрессорного гена p53 состоит в остановке роста в ответ на потерю теломер в старых клетках. Этой гипотезе не противоречат данные о поведении большинства опухолей, в которых этот ген мутирован, и объясняют характеристики редких типов опухолей, в которых функция p53 сохранена.
Недавно показано, что in vitro старые фибробласты человека стимулируют пролиферацию предраковых и злокачественных эпителиальных клеток, которые способны образовывать опухоли при прививке голым мышам (“плохие соседи”) [17]. У фибробластов на более ранних пассажах (пресенильных) эта способность менее выражена. Несмотря на довольно убедительные аргументы в пользу связи между старением клеток, биологией теломеры и раком человека, данные о роли клеточного старения в старении человека довольно противоречивы и требуют дальнейших исследований.
Увеличение продолжительности жизни и риск рака
| Все равно, по какой дороге идти. Куда-нибудь ты наверняка попадешь, если будешь идти достаточно долго. Льюис Кэррол. Приключения Алисы в Стране чудес |
Расчеты показывают, что у человека с продолжительностью жизни 70 лет, крысы (2.5-3 года) и мыши (2-2.5 года) частота новообразований одинакова и составляет 30% [3, 19]. Однако при нормализации по количеству клеток в организме оказывается, что мышь более склонна к развитию опухолей, чем человек. Этот феномен склонны объяснять тем, что в соматических клетках мыши теломераза более активна, чем в клетках человека, чему соответствует значительно большая длина теломер у мыши по сравнению с человеком. Вместе с тем видовая продолжительность жизни млекопитающих соотносится с эффективностью репарации ДНК и устойчивостью их клеток к окислительному стрессу. Так, в ДНК человека восстановление гуанина, алкилированного канцерогенными нитрососоединениями, в сотни раз выше, чем у мыши, что связано с большей резистентностью человека к этим агентам. Оказалось, что эффективность репарации вызываемых канцерогеном повреждений в ДНК различных органов связана с продолжительностью жизни мышей. Анализ данных о частоте рака у генетически модифицированных животных с увеличенной продолжительностью жизни говорит о снижении у них частоты злокачественных новообразований [3].
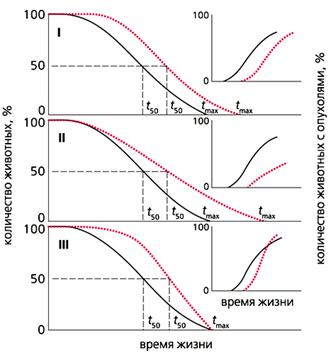
Рис. 4. Влияние трех типов геропротекторов (цветные кривые) на продолжительность жизни животных
и частоту спонтанных опухолей (на врезке).
I - продолжительность жизни всех особей увеличивается, скорость старения не меняется, новообразования развиваются позднее, чем в контрольной группе, но с такой же частотой;
II - скорость старения снижается, частота новообразований уменьшается;
III - продолжительность жизни не меняется, а скорость старения и количество новообразований растет.
tmax - максимальная продолжительность жизни, t50 - время полужизни.
Показано, что различные геропротекторы по-разному влияют на развитие новообразований, что определяется в основном типом замедления старения в популяции (рис.4). Наши наблюдения свидетельствуют, что более “прямоугольный” характер кривых выживания ассоциирован с увеличенной скоростью развития фатальных опухолей у крыс.
С конца ХVIII в. и вплоть до середины ХХ в. в экономически развитых странах постепенно увеличивалась частота раковых заболеваний. Частота всех злокачественных новообразований в США и многих других странах, включая Россию, заметно росла с 1960 по 1990 г. и у мужчин, и у женщин, что совпадало с увеличением продолжительности жизни. С 90-х годов ХХ в. в наиболее экономически развитых странах, таких как США, Швеция и Дания, частота рака стала несколько снижаться (рис.5). И именно во второй половине ХХ в. в наиболее развитых странах изменился характер траекторий смертности, что сопровождалось уменьшением смертности в самых старших возрастах (рис.6). В то же время в других странах, например в России, частота злокачественных новообразований продолжает расти. Очевидно, что увеличение продолжительности жизни и характера траектории смертности по типу параллельного сдвига или даже увеличения ее наклона будет способствовать снижению частоты злокачественных новообразований.
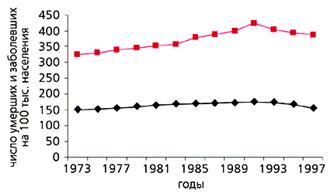
Рис. 5. Динамика заболеваемости (цветная кривая) и смертности от рака в США в XX в. [19].
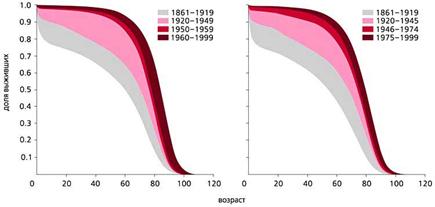
Рис. 6. Динамика изменений выживаемости женщин (слева) и мужчин в Швеции с 1861 по 1999 г. [20].
* * *
Наш великий соотечественник И.И.Мечников - основоположник современной геронтологии и автор самого термина “геронтология” - весной 1907 г. в своем предисловии к первому изданию “Этюдов оптимизма” писал: “Наука в России переживает продолжительный и тяжелый кризис. На науку не только нет спроса, но она находится в полнейшем загоне”. К сожалению, сегодня в России ситуация мало чем отличается от той, что была 100 лет назад. Бюджет всей российской науки не превышает бюджета Национального института старения США - одного из самых “маленьких” в системе Национального института здоровья, и существенно меньше, чем средства, выделяемые государством на содержание чиновников. Однако стареют и они.
И к ним, а также нашим депутатам, мы адресуем Открытое письмо, опубликованное недавно в Интернете (http://cureaging.com; http://ieet.org/index.php/IEET/ more /scientistsantiaging/; http://www.longevity-science.blogspot.com/ и др.), подписанное 54 ведущими учеными-геронтологами мира. Ниже приводится перевод этого письма на русский язык:
“Всем, кого это касается.
На моделях многих сильно отличающихся друг от друга видов лабораторных животных (нематоды, дрозофилы, мыши и т.п.) удается достигнуть замедления старения и продления активной жизни. Поэтому, исходя из общности фундаментальных механизмов старения, есть основания полагать возможным замедление старения у людей.
Расширение наших знаний о старении позволит лучше противостоять таким истощающим организм патологиям, связанным со старением, как рак, сердечно-сосудистые заболевания, диабет II типа и болезнь Альцгеймера. Терапия, основанная на знании фундаментальных механизмов старения, будет способствовать лучшему противодействию этим возрастным патологиям.
Именно поэтому наше письмо является призывом к увеличению финансирования этой области, что крайне необходимо для интенсификации как исследований фундаментальных механизмов старения, так и поиска способов его замедления. Все это может привести к намного большим дивидендам, чем в случае, если бы эти же средства и усилия вкладывались в непосредственное противостояние возрастным патологиям. Поскольку механизмы старения становятся все более и более понятными, могут быть разработаны эффективные средства вмешательства в этот процесс. Это позволит значительному количеству людей продлить здоровую и продуктивную жизнь”.
Список литературы1. Oeppen J., Vaupel J.W. // Science. 2002. V.296. P.1029-1031.
2. Старение населения Санкт-Петербурга: социально-демографические аспекты / Ред. Г.Л.Сафарова. СПб., 2006.
3. Анисимов В.Н. Молекулярные и физиологические механизмы старения. СПб., 2003.
4. Handbook of the Biology of Aging. Sixth Edition / Eds E.J.Masoro, S.N.Austad. Amsterdam, 2006.
5. Cutler Mattson // Aging Cell. 2006. V.6.
6. Perls T., Levenson R., Regan M., Puca A. // Mech. Ageing Dev. 2002. V.123. P.231-242.
7. Westendorp R.G., Kirkwood T.B.L. // Nature.1998. V.396. P.743-746.
8. Orr W.C., Sohal R.S. // Science. 1994. V.263. P.1128-1130.
9. Parkes T.L., Elia A.J., Dickinson D. et al. // Nature Genetics. 1998. V.19. P.171-174.
10. Tatar M., Bartke A., Antebi A. // Science. 2003. V.299. P.1346-1351.
11. Lin Y.J., Seroude L., Benzer S. // Science.1998. V.282. P.943-946.
12. Coschigano K.T., Clemmons D., Bellush L.L., Kopchick J.J. // Endocrinology. 2000. V.141. P.2608-2613.
13. Bartke A., Coschigano K., Kopchick J. et al. // J. Gerontol. Biol. Sci. 2001. V.56A. P.B340-B349.
14. Holzenberger M., Dupond J., Ducos B. et al. // Nature. 2003. V.421. P.182-187.
15. Fontana L., Meyer T.E., Klein S., Holloszy J.O. // Proc. Natl. Acad. Sci. USA. 2004. V.101. P.6659-6663.
16. Anisimov V.N., Berstein L.M., Egormin P.A. et al. // Exp. Gerontol. 2005. V.40. P.685-693.
17. Krtolica A., Campisi J. // Успехи геронтол. 2003. Т.11. С.109-116.
18. Anisimov V.N., Ukraintseva S.V., Yashin A.I. // Nature Rev. Cancer. 2005. V.5. P.807-819.
19. Ries L.A.G., Eisner M.P., Kosary C.L. et al. // SEER Cancer Statistics Review 1973-1998. Nat. Can. Institute. Bethesola. MD. 2001.
20. Vashin A.I., Begun S.S., Boikos I. et al. // Mech. Ageing Dev. 2002. V.123. P.637-6
Похожие работы
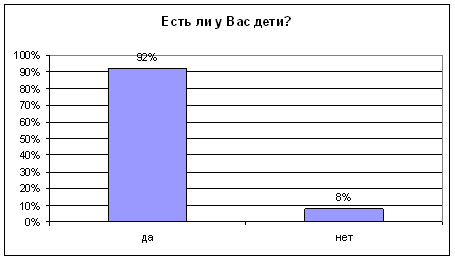
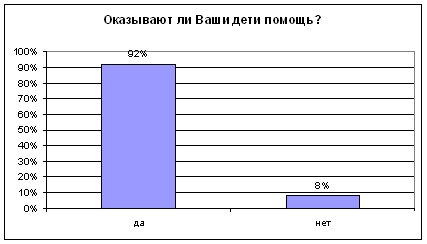
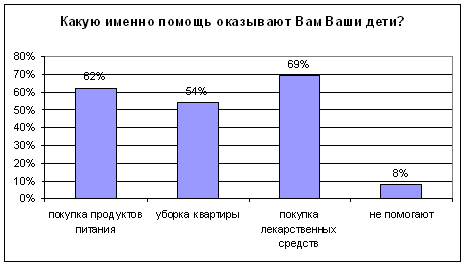
... было не однозначным: от полного отказа в помощи старикам до создания относительно комфортных условий жизни. Во второй главе "Практические подходы к организации социальной помощи пожилым людям" рассмотрено оказание социальной помощи на примере Пенсионного Фонда Российской Федерации. Приведены нормативно-правовые акты, на основании которых назначаются пенсии и другие социальные выплаты. Описан ...
... , находимые там и датируемые 6-4 тысячелетием до нашей эры показывают нам богатый животный мир Африки. На фресках изображены буйволы, антилопы, бегемоты. Как показывают исследования опустынивание саванны на территории современной Сахары началось около 500 000 лет назад, но обвальный характер процесс принял с 3 т. до н. э. Характер жизни кочевых племен Юга Сахары, образ жизни, которых не слишком ...
... . В-третьих, это круг вопросов, связанных с решением проблем ослабленного здоровья пожилых людей. В-четвёртых, немаловажным является решение проблемы досуга, культурно-нравственной востребованности пожилых людей, рекреации, адаптации пожилого человека, ещё недавно принадлежащего к трудоспособной возрастной когорте, к новой социальной роли. Разумеется, в разных странах, в различных культурах все ...
... : 1 – НЕЗАВИСИМОСТЬ; 2 – УЧАСТИЕ; 3 – УХОД; 4 – РЕАЛИЗАЦИЯ ВНУТРЕННЕГО ПОТЕНЦИАЛА; 5 – ДОСТОИНСТВО. ГЛАВА 2. ТЕХНОЛОГИИ СОЦИАЛЬНОЙ РЕАБИЛИТАЦИИ ЛИЦ ПОЖИЛОГО ВОЗРАСТА 2.1. Медико – социальная реабилитация пожилых людей В социальной работе с пожилыми людьми используются разно образные формы и методы. Это и социальное обслуживание на дому, и срочная социальная помощь, и ...
0 комментариев