Марфенин Н. Н.
Временная колония, несмотря на слабую физиологическую интеграцию, дает животному некоторые экологические преимущества, изучение которых позволяет яснее представить возможные причины возникновения колониальности. Колониальная организация увеличивает протяженность тела гидроида, способствуя образованию “клубка” — ассоциации нескольких колоний. Такое объединение помогает совместно удерживать и поедать относительно крупную добычу, контролировать значительное пространство. Молодые колонии, развивающиеся в клубке, имеют больше шансов выжить.
Гидроиды рода Moerisia занимают особое место в пределах отряда Leptolida из-за крайней примитивности колониальной организации. Moerisia можно назвать “морскими гидрами”, так как они по своей анатомии, способу вегетативного размножения и строению колонии похожи на пресноводную гидру.
Подробное изучение биологии таких животных должно помочь в познании происхождения колониальности.
Морфология полипоидной стадии М. maeotica, как и бесполое размножение гидроида, стала известна в основном благодаря работам М. В. Пальчиковой-Остроумовой (Paltschikowa-Ostroumova, 1925) и Г. Паспалева (Paspaleff, 1938). Важны также наблюдения Боулингира (Boulenger, 1908) на М. lyonsi, описавшего этот род. В то же время исследования роста и формирования колонии у гидроидов рода Moerisia, важные для понимания явления колониальности, мне не известны, так же как и работы по питанию гидрантов и степени физиологической интеграции колонии. Этим вопросам посвящена настоящая работа.
Материал и методикаMoerisia maeotica (Ostroumow, 1896) принадлежит к семейству Моеrisiidae подотряда Limnomedusae.
Полипоидное поколение имеет вид одиночных полипов или образует небольшие стелющиеся по субстрату колонии (Наумов, 1960). Сами полипы обладают полыми щупальцами, характерными для этого семейства. Видовая принадлежность определяется по размерам полипов, а также по порядку почкования: медузоидные почки закладываются на теле полипа среди щупалец, а латеральные почки дочерних полипов — ниже щупалец.
Вид распространен в Азовском, Черном и Каспийском морях, а также в солоновато-водных и значительно опресненных водоемах бассейнов Азовского, Черного, Средиземного и Эгейского морей (Наумов, 1968).
Живой материал, использованный в настоящем исследовании, был собран И. А. Косевичем во время отбора проб оброста с корпуса НИС “Эксперимент” в Каспийском море в августе 1981 г.
Гидроиды содержали в чашках Петри с каспийской водой без дополнительной аэрации, кормили науплиусами Artemia. Температура оставалась в пределах 18—20° С. Исследование проводили с января по май 1982 г.
РезультатыФормирование первичного полипа. Развитие колонии было прослежено со стадии латеральной почки, отделившейся от материнского полипа (рис. 1, 1). Начальные размеры продолговатой грушевидной почки: 0,3—0,4 мм в длину и примерно 0,1 мм в диаметре в самом широком месте.
Обычно сразу после отделения на проксимальном конце почки образуются три-четыре коротких щупальца и небольшой гипостом (рис. 1, /, 2). Длина такого полипа около 0,6 мм. Он уже способен заякоривать свежевылупившегося науплиуса, но проглотить его целиком еще не может.
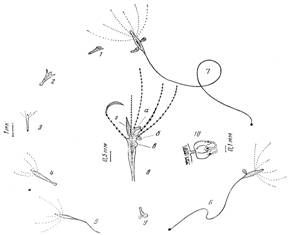
Рис. 1. Схема развития гидранта из латеральной почки. 1—латеральная почка после отделения от материнского полипа, 2 — формирование щупалец и гипостома, 3 — молодой гидрант после метаморфоза, 4 — выросший гидрант с несформировавшейся ножкой, 5—гидрант после образования ножки, 6—удлинение ножки и начало почкования: бугорок и миндалевидная стадия латеральной почки, 7—достижение окончательных размеров ножки; продолжение почкования: миндалевидная и грушевидная (завершающая) стадии латеральной почки, 8—тело гидранта при большем увеличении: хоботовидный гипостом (а), медузоидная (б) и латеральная (в) почки на ранних стадиях развития, расслабленные и сжатые (г) щупальца, 5 — основание ножки с прикрепительным диском (слева) и подоцистой (справа), 10—медузоидная почка при еще большем увеличении
Через 3 сут (при ежедневном кормлении) длина полипа становится около 2 мм, а его щупальца достигают 1,5 мм (рис. 1, 4). Затем наступает период его быстрого удлинения примерно вдвое — сильно вытягивается и становится тонкой ножка. Вероятно, это стадия формирования настоящей ножки (рис. 1, 5, 6). Одновременно число щупалец увеличивается до 5 или 6, они располагаются неправильным венчиком и способны в 10 раз удлиняться или сжиматься. Гипостом становится вытянутым, хоботовидным и подвижным, подошва гидранта слегка расширяется, желтеет и в ней формируется подоциста.
Дальнейший рост полипа происходит за счет постепенного удлинения ножки. Зона роста, очевидно, находится в основании тела гидранта, так что дистальная часть ножки оказывается наиболее молодой (рис. 1, 6). В это время она сохраняет способность продольно сжиматься в 3—4 раза, становясь временно толще в поперечнике.
После периода постепенного удлинения ножки гидранта, как правило, наступает этап (в данных наблюдениях на 12—14 сут) скачкообразного ее прироста. Менее чем за сутки она может удлиниться в 1,5—2 раза, достигнув 15—18 мм. При этом она становится еще тоньше и уже не может сокращаться так же сильно и быстро, как раньше (рис. 1, 7). Теперь перемещение гидранта осуществляется с помощью его щупалец— он вытягивает одно-два щупальца, прикрепляет их каким-то образом к субстрату, а затем, сокращая их, подтягивает тело. В дальнейшем удлинение ножки происходит незначительно.
Своих наибольших размеров полип достигает при длине ножки до 3D мм (обычно около 20 мм) и диаметре 0,05—0,1 мм. Ножка плавно переходит в веретеновидное тело 0,7—2,0 мм длиной. Щупалец, как правило, уже восемь, а их длина в расправленном состоянии до 6 мм. Ги-постом примерно 0,1 мм, в поперечнике длиной 0,3—0,5 мм (рис. 1, и). Данных размеров полип при благоприятных обстоятельствах может достичь при лабораторном культивировании за 12 сут.
Бесполое размножение осуществляется у М. maeotica несколькими способами: образованием латеральных почек, формированием медуз, образованием подоцист и фрагментацией колонии.
Образование латеральных почек начинается примерно на 10—15-е сут, т. е. уже у взрослого полипа. Обычно почка появляется на 0,3— 0,5 мм ниже венчика щупалец. Сначала она имеет вид полусферического бугорка (рис. 1,5), затем становится миндалевидной и, наконец, принимает грушевидную форму (рис. 1, 7), ориентированную узкой частью дистально (рис. 2, 7). Тонкая ножка, которой почка еще удерживается, судя по ее прозрачности, состоит только из эктодермы. На этой стадии развития почки полип льнет к субстрату, в результате чего она имеет возможность прикрепиться к нему своим дистальным утонченным концом. Все развитие почки занимает около 2 сут.
Одновременно на полипе могут развиваться одна-две латеральные почки. Их число и частота образования новых зависят от интенсивности питания. При хорошем питании новые почки могут появляться через каждые 2 сут.
Первые медузоидные почки появляются часто сразу после достижения полипом взрослых размеров, т. е. начиная примерно с 13 сут развития. Начальные размеры этих почек, шаровидных по форме, возникающих среди щупалец, 0,07 мм в диаметре. С самого начала они отличаются от латеральных почек не только своим местоположением, но также цветом (они прозрачны) и формой—у них сразу образуется ножка (рис. 1, 10). По-видимому, пища не может непосредственно поступать в медузоидную почку, так как никогда не приходилось видеть ее желтой или оранжевой, т. е. окрашенной в цвет пищи.
При благоприятных условиях развитие медузоидной почки завершается за 5—7 сут, после чего она отделяется. Колокол молодой медузки напоминает четыре соединенные друг с другом полусферы. Диаметр его около 0,6 мм. Медузка имеет четыре щупальца длиной по 1,2 мм. Щупальца сразу способны парализовать и удерживать науплиуса, однако заглатывания его наблюдать не удалось, да и представить заглатывание жертвы размером с науплиуса трудно, так как ротовой стебелек у медузки очень короткий.
Медузки малоактивны и подолгу лежат на дне на одном месте. Частота пульсаций, впрочем, достигает иногда 20 за 1 мин. Возможно, условия питания не соответствовали образу жизни медузок или отрицательно сказывался небольшой объем сосуда, так как проследить дальнейшее развитие их не удалось.
Подоцисты образуются в прикрепительных дисках гидрантов или рядом с ними (рис. 1, 9) при хорошем питании. Заметить их образование легко по ярко-желтой однородной окраске, которую приобретает прикрепительный диск. Клеточные слои и полость прижизненно в подоцисте не заметны.
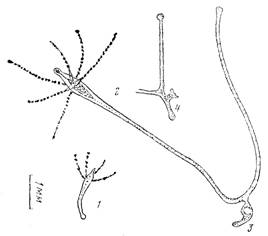
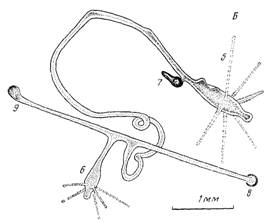
Рис. 2. Образование колонии. А:: 1— гидрант на 3-и сут развития; 2—гидрант на 11-е сут развития; 3—неотделившаяся почка до прикрепления; 4 — то же после прикрепления почки, образования столона и дочернего гидранта на нем на 12-е сут; Б—колония на 24-е сут развития; 5—первичный гидрант;6—второй гидрант; 7—грушевидная латеральная почка, которая может стать третьим гидрантом; 8—прикрепительный диск первого гидранта; 9 — прикрепительный диск второго гидранта
При дальнейшей жизни гидроида никаких изменений подоцист не происходит, и, вероятно, они могут переживать неблагоприятные условия после гибели организма (Paspaleff, 1938), но в данном случае мы это не изучали.
Четвертый способ бесполого размножения—фрагментация колонии— будет рассмотрен ниже.
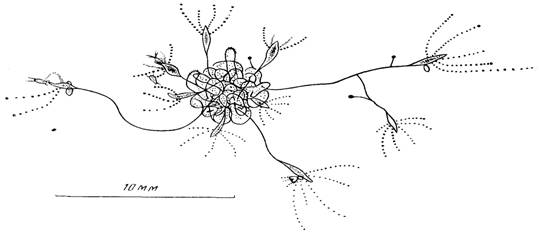
Рис. 3. Фрагментация колонии. Л — колония с тремя гидрантами и десятью прикрепительными дисками (черные точки) на 56-е сут развития. Б — та же колония после фрагментации на 74-е сут развития .
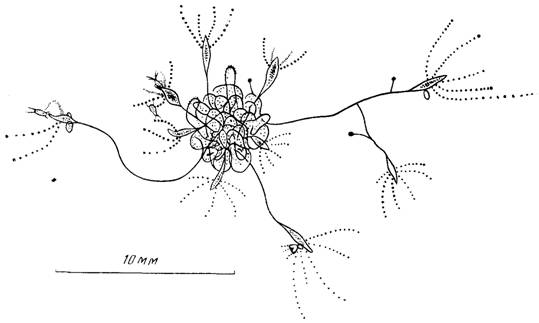
Рис. 4. Ассоциация нескольких колоний в виде “клубка”
Формирование колонии. Еще до достижения взрослых размеров у полипа может образоваться почковидное ответвление. Оно закладывается чаще всего в проксимальной части тела полипа, а по мере удлинения ножки оказывается все дальше от его тела (рис. 2, А, 3). Через несколько дней после возникновения вырост становится столоновидным и прикрепляется к субстрату. Затем в его средней части формируется гипостом со щупальцами (рис. 2, А, 4), однако связь с материнским гидрантом не прекращается. Дальнейшее развитие дочернего гидранта зависит от его питания и удаленности от материнского гидранта. При кормлении только материнского гидранта развитие дочернего явно задерживается вплоть до полной остановки. Торможение развития дочернего гидранта тем сильнее, чем длиннее становится ножка материнского гидранта, что, по-видимому, связано с неполноценным транспортом пищи в колонии от одного гидранта к другому.
От материнского полипа может произойти образование и второго дочернего полипа. В свою очередь от дочерних гидрантов продолжается рост колонии с помощью столонообразных выростов и формирование гидрантов третьего порядка. Удалось проследить развитие колонии до стадии четырех гидрантов, что заняло 70 сут.
По мере увеличения числа гидрантов растет и число прикреплений колонии к субстрату. Так, на стадии трех гидрантов (56 сут) в наблюдаемой колонии было зарегистрировано 10 прикрепительных дисков (рис.З,А).
Одновременно в колонии продолжается отпочковывание латеральных почек и рост из них дочерних колоний, которые вскоре переплетаются друг с другом, образуя настоящий клубок (рис. 4). Такая ассоциация колоний может быть полезной в их существовании. Обычно наиболее крупные гидранты вытягиваются из “клубка” на 1—1,5 см, что позволяет “клубку” контролировать площадь примерно в 3—7 см2. Гидрант, поймавший добычу, часто подтягивается благодаря сжатию ножки в направлении к “клубку”. Это приводит к тому, что крупная жертва становится добычей по крайней мере нескольких гидрантов из “клубка”. Такая поддержка должна быть жизненно важной для молодых гидрантов, малые размеры которых значительно уменьшают вероятность поимки ими добычи и сокращают продолжительность безболезненного голодания в отсутствие пищи. Кроме того, запутываясь в “клубке”, гидрант таким способом прикрепляется к нему, поэтому “клубок” для некоторых гидрантов является также субстратом.
Фрагментация колонии. В наших опытах взрослые отдельные колонии состояли не более чем из четырех гидрантов. По мере дальнейшего роста такие колонии обязательно распадались. Обычно этому предшествовало прикрепление одного или нескольких гидрантов к субстрату выростами-столонами. Через некоторое время ножка гидранта сокращалась и натягивалась как струна между новым прикреплением и старой частью колонии. Еще через несколько дней эта перемычка разрывалась (рис. 3, Б).
В другом варианте отдельный взрослый гидрант отрывается от колонии, не прикрепляясь перед этим к субстрату. Сначала ножка гидранта обвивается вокруг столона или ножки другого гидранта, а найдя опору, через некоторое время обрывается. Оторвавшийся гидрант не менее нескольких суток остается неприкрепленным на гладком дне чашки Петри или слабо удерживается за дно щупальцами. В естественных условиях он, вероятно, может прикрепляться к водорослям или другому нитевидному субстрату, обвивая его ножкой. Не исключена также возможность регенерации подошвы и прикрепления с ее помощью к субстрату, но это не было прослежено до конца. В обоих случаях фрагменты колонии жизнеспособны, а фрагментация колонии, следовательно, является настоящим бесполым размножением.
Питание. У голодных колоний гидранты вытягиваются и раскидывают щупальца, поджидая жертву. Ножки у полипов обычно удлиненные, часто образующие петли. Они обладают спонтанной активностью, медленно меняя свое положение.
Заякоривание (удержание, загарпунивание) науплиуса происходит моментально при соприкосновении его со щупальцами, которые сразу сокращаются, подтаскивая жертву к гипостому. Остальные щупальца обычно остаются в расправленном рабочем положении.
Гипостом сначала обследует добычу, изгибаясь к ней наподобие хобота и ощупывая ее. Найдя конец добычи (хвост или голову), гипостом начинает раскрываться, одновременно натягиваясь на добычу. Продолжительность проглатывания свежевылупившегося науплиуса Artemia от момента заякоривания до смыкания гипостома занимает чаще всего 4— 4,5 мин, но может длиться и до 8 мин.
Гидрант способен одновременно заякорить нескольких науплиусов, однако проглатывает он их по очереди. В наших опытах гидранты могли проглотить одного за другим не более четырех науплиусов. Продолжительность заглатывания второго, третьего и четвертого науплиусов остается примерно такой же, как и первого.
Установить продолжительность переваривания пищи в гидранте не удалось. Это просто сделать, если пища из гидранта после завершения ее переваривания выводится в ценосарк, как у большинства колониальных гидроидов; в данном же случае пища остается в гидранте надолго.
Во время переваривания пищи разбухания полипа из-за гидратации не происходит. Однако вскоре появляется латеральная почка, которая вбирает в себя, судя по интенсивности желто-оранжевого цвета энтодермы, значительную часть пищи. Возможно, образование латеральной почки компенсирует необходимое увеличение объема полипа при пищеварении.
Во время переваривания пищи тело гидранта пульсирует подобно описанным ранее пульсациям у других видов гидроидов (Марфенин, 1981). Волна поперечного сжатия (утончения) тела гидранта начинается сразу под гипостомом. Затем сжатие распространяется проксимальнее. Пища выдавливается в дистальную часть ножки, а когда сжатие доходит до ножки, она перегоняется снова в тело гидранта, которое к этому времени расслабляется. Пульсации имеют полутораминутный период при 20° С. Через четыре—семь периодов (т. е. 6—10 мин) у молодых гидрантов часто наблюдается продольное сжатие ножки гидранта, он как бы оседает, после чего ножка медленно вытягивается.
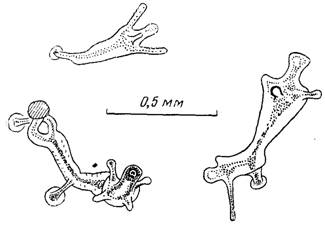
Рис. 5. Гидранты на 56-е сут голода
Сытые гидранты обычно бывают сжатыми: укорачиваются щупальца и ножка гидранта, тело его также становится короче и шире. В этом состоянии гидрант не теряет способности заякоривать и проглатывать добычу, но вероятность заяко-ривания значительно уменьшается.
В нескольких случаях удалось наблюдать экскрецию, которая происходила примерно через 7 ч после проглатывания науплиуса. Гипостом при этом выворачивается наизнанку, выбрасывая непереваренные остатки — шкурку науплиуса.
Наблюдения за состоянием гидроида при продолжительном голодании показали, что одиночные полипы небольших размеров (длина ножки в расслабленном состоянии 8 мм, щупалец 7 мм) выживают без пищи в течение более чем 40 сут. В первые дни полипы сильно вытягиваются, но уже к 20-м сут уменьшаются до 1 мм, после чего еще 20 сут остаются почти без изменений. Затем щупальца укорачиваются и рассасываются, хотя миниатюрные остатки полипов (около 0,2 мм) живут еще не менее одного месяца (рис. 5).
Транспорт пищи в колонии. Функционирование распределительной системы зависит от размеров колонии. У молодого первичного гидранта с ножкой менее 10 мм длиной перенос пищи в проксимальную часть ножки и педальный диск осуществляется при продольном сокращении ножки. Такие сокращения, приводящие как бы к оседанию полипа, происходят регулярно во время переваривания пищи. Благодаря им внутриполостное давление должно повышаться, что, вероятно, и приводит к расширению просвета ножки гидранта и выдавливанию пищи в проксимальном направлении. Так происходит заполнение пищей прозрачного педального диска, приводящее к образованию подоцисты.
В молодой колонии, состоящей из двух гидрантов, также зарегистрирован перенос пищи от одного гидранта к другому. Это происходит после первичного переваривания пищи, когда объем гидроплазмы увеличивается из-за поступления пищи и, возможно, из-за гидратации, связанной с возрастанием осмотического давления в гастральной полости. Перистальтические волны сжатия полипа, направленные проксимально, способствуют перемещению гидроплазмы в ножку гидранта. Во время сжатия тела гидранта избыток гидроплазмы вместе с частицами пищи выдавливается в ножку. Создается участок расширения или вздутия ножки, который иногда оказывается изолированным от тела гидранта участком сжатия. Проксимально распространяющаяся по ножке волна сжатия несколько продвигает перед собой вздутие и вскоре угасает. Следующая волна может способствовать при сильном сжатии продвижению порции гидроплазмы еще дальше. Так незначительные избытки пищи достигают дочернего гидранта.
При значительном избытке гидроплазмы, наблюдаемом лишь однажды, возможно расширение просвета на всем протяжении еще короткой ножки гидранта (менее 10 мм). В этом случае было видно, как частицы пищи двигаются то в одном, то в противоположном направлении, в зависимости от фазы пульсации тела гидранта.
У взрослых гидрантов с длинной тонкой ножкой транспорт пищи к подошве, очевидно, исключается. Проксимальная часть ножки у такого гидранта обречена на голодание.
Таким образом, интеграция колонии у М. maeotica посредством распределительной системы кратковременна, связана с периодами избытка пищи и осуществляется лишь на очень короткие расстояния (в несколько мм), а не по всей колонии. Гидранты одной, даже маленькой колонии чаще всего оказываются физиологически изолированными друг от друга.
Обсуждение результатовГидроид М. maeotica представляет пример временной колонии, существование которой во многом подчинено целям бесполого (вегетативного) размножения.
В отличие от более сложных постоянных колоний у временных:
1) рост колонии приводит к ее фрагментации, а не к значительному возрастанию числа связанных воедино гидрантов; 2) отдельные зооиды могут выходить из состава колонии и продолжать жить самостоятельно; 3) нет постоянных органов роста — специализированных верхушек роста; 4) физиологическая интеграция кратковременна и ограничена лишь малыми расстояниями; основное значение физиологической интеграции в поддержании существования на первом этапе развития молодых гидрантов.
Гидроиды из рода Moerisia и раньше привлекали внимание исследователей особой примитивностью колониальной организации. Так, Вылканов (Вълканов, 1950) считал, что дочерние гидранты в колонии просто недоразвиты и должны со временем отпочковаться, на основании чего он отрицал возможность существования колоний у этих гидроидов.
Теперь мы видим, что хотя в основном А. Вылканов оказался прав, вопрос о колониальной организации у М. maeotica более сложен, так как колонии с несколькими взрослыми гидрантами могут все же подолгу существовать. Этот простой факт меняет смысл всей аргументации. Необходимо понять значение наблюдаемой задержки в вегетативном размножении. Почему наряду с латеральными почками, отделяющимися еще до появления каких-либо щупалец, у гидроида есть и неотделяющиеся почки? Очевидно, образование временной колонии, наблюдаемое нами у М. maeotica, имеет некий больший смысл, чем простое вегетативное размножение.
Прежде всего колониальное строение позволяет поддержать на первом этапе существования молодых дочерних полипов, что особенно важно в условиях недостатка пищи. Мы видели, что ограниченный транспорт пищи от материнского гидранта к дочернему возможен.
Кроме того, образование даже небольших по числу гидрантов колоний способствует удлинению в 2—3 раза суммарной протяженности ножек гидрантов, которые, переплетаясь, образуют “клубок”—разновидность агрегации животных. Концентрация полипов должна быть для них важной при ловле и переваривании крупной добычи. Помимо этого, молодые гидранты, осевшие в клубке, имеют больше шансов прокормиться за счет остатков пищи, выловленной взрослыми гидрантами. Мне приходилось наблюдать, как молодые гидранты проглатывали науплиусов, притащенных на щупальцах взрослыми гидрантами.
Образование столонов, приводящее часто к возникновению дочерних гидрантов, помогает гидрантам лучше закрепиться или поменять место прикрепления. Последнее очень важно для сидячих животных, которые подвергаются постоянной опасности быть занесенными детритом или погребенными под другими эпибионтами. С помощью новых столонов при последующем обрыве старых прикреплений колония может медленно перемещаться по субстрату. В то же время она оставляет на местах былых прикреплений обрывки ножек с подоцистами. Возможно, создание запаса временно пассивных подоцист имеет положительное значение для вида. Размножение с помощью подоцист, к сожалению, пока что изучено очень мало (Иванова-Казас, 1977).
Сохранение примитивной организации колонии, по-видимому, было связано с особым развитием самих гидрантов. При относительно малых размерах гидрант обладает очень длинными щупальцами, способными в десять и более раз менять свою протяженность. Это становится возможным, вероятно, из-за того, что щупальца полые. В результате один гидрант контролирует достаточно большое пространство вокруг себя. Кроме того, длинная ножка позволяет перемещаться гидранту в радиусе 2 см, т. е. контролировать площадь около 10 см2.
Один и тот же результат—контроль возможно большего объема окружающего пространства—гидроиды достигают по крайней мере двумя различными путями: либо увеличением размеров ловчего аппарата гидранта, либо усовершенствованием колониальной организации.
Несмотря на то что колония у М. maeotica очень примитивна, не так просто даже на этом уровне колониальной организации выделить отдельные “особи” или, как еще говорят, “члены” колонии. Действительно, гидрант можно считать, безусловно, особью лишь на самом первом этапе, когда он еще один. При этом у него можно выделить несколько отделов: самое тело со щупальцами и гипостомом, ножку и подошву (прикрепительный диск, превращающийся со временем в подоцисту).
В дальнейшем от ножки начинает расти столон, а на столоне часто образуется дочерний гидрант. Кому принадлежит столон: первичному шдранту или дочернему? Более того, ножка первичного гидранта проксимальнее места ответвления столона с момента прикрепления его к субстрату также становится столоном, образуя одно целое с ним (рис. 2, Б), от которого отходят как первичный, так и дочерний гидранты. Получается, что часть зооида в течение его жизни становится общеколониальной. Это позволяет понять, что то, что мы называем “членом” колонии, представляет собой лишь часть организма и поэтому может называться особью лишь весьма условно.
Таким образом, даже примитивные временные колонии не могут быть рассматриваемы как агрегации особей. Признаки целостности колонии появляются с момента возникновения колониальной организации. Значение временной колонии не сводится к незавершенному вегетативному размножению, так как колониальность дает ряд функциональных и экологических преимуществ гидроиду.
Список литературыВълканов А. Съществуват ли колониални меризинди?—Тр. Морск. биол. ст. Сталин, 1950, т. 15, с. 187—192.
Иванова-Казас О. М. Бесполое размножение животных. Л.: Изд-во ЛГУ, 1977. 240 с.
Марфенин Н. Н. Некоторые особенности пищеварения в гидрантах у различных колониальных идроидов.—Ж. общ. биол., 1981, т. 42,№ 3, с. 399—408.
Наумов Д. В. Гидроиды и гидромедузы морских, солоноватоводных и пресноводных бассейнов СССР. М.—Л.: Изд-во АН СССР, 1960. 585 с.
Наумов Д. В. Тип кишечнополостные Coelenterata. — Атлас беспозвоночных Каспийского моря. М.: Пищевая пром-сть, 1968, с. 44—50.
Boulenger Ch. L. On Moerisia lyonsi, a new hydromedusan from Lake Qurun.— Quart. J. Microscop. Sci., 1908, v. 52, № 207, p. 357—378.
Paltschikowa-Ostroumowa M. W. Moerisia inkermanica n. sp.—Zool. Anz, 1925 v. 62, № 11/12, р. 273—284.
Paspaleff G. W. Die polypen Ostroumovia inkermanica (Pal. Ostr.) — Трудове на Черно-морската биологична станция въ Варна, 1938, № 7, с. 29
0 комментариев