Чебкасов С.А, Ефимов В.Н., Ласков В.Н, Иванова Е.А.
НИИНК РГУ, Ростов-на- Дону, Россия
В опытах на 7 морских свинках, обездвиженных тубокурарином, показано, что миллисекундное (2мс интервал) совпадение экстремумов ЭКоГ в различных отведениях есть закономерное и регулярное явление для бодрствующего мозга.
Полученные результаты согласуются с концепцией дискретно-волнового механизма нервной интеграции и ее приложением к ЭЭГ. Эта концепция содержит представление об иерархии смысловых квантов ЭЭГ , по которому квазиритмические колебания - "волны" ЭЭГ есть ее энергетический компонент или же колебания возбудимости, тогда как информационным компонентом ЭЭГ являются дискретные "пики" (экстремумы), отображающие апериодические эпизоды формирования миллисекундных интегративных нейронных макроансамблей в коре мозга. В частности показано, что кроме периодических эпизодов миллисекундной синхронизации экстремумов ЭкоГ (ЭМС), которые могут быть разделены интервалами, соответствующими гамма, бета, сигма, альфа, дельта диапазонам ЭЭГ, ее "волнам", регулярно имеет место апериодическое следование ЭМС. Визуальное отслеживание записей ЭКоГ с должной временной разверткой показало миллисекундную синхронизацию именно "пиков", экстремумов ЭКоГ, но не ее "волн" - например, гамма осцилляций.
Показано, в соответствие с нашим концептуальным походом, что позитивных ЭМС , рассматриваемых по аналогии с волнами первичного ответа как тормозные, обнаруживается больше, чем негативных - возбудительных.
Показано, что внутрианализаторные взаимодействия реализуются достоверно большим числом ЭМС, чем внеанализаторные. Это соответствует нашему представлению об иерархии смысловых квантов ЭЭГ.
Проанализированы ближайшие проблемы исследования смысловых квантов ЭЭГ как метода адекватного шкалирования ЭЭГ с учетом нелинейности биологического времени.
Введение
Теоретически и на базе косвенных экспериментальных свидетельств была обоснована идея [4-14], что в ЭЭГ отображается иерархия циклически повторяющихся квантов функционирования полярных нервных модулей разного уровня сложности, которые образуют вложенную иерархию и вместе представляют собою мозг.
Интегрированное функционирование модулей порождает все более объемные временные функциональные ансамбли, что отображается в поэтапной миллисекундной синхронизации нейронных спайков. Такими временными макроансамблями нейронов кодируются все более сложные акты деятельности мозга.
Каждый временной макроансамбль служит для кодирования результатов нервной интеграции (на том или ином уровне), достигаемых в результате акта (той или иной сложности) функционирования соответствующей нервной структуры.
Синхронизация активности во временном макроансамбле нейронов порождает соответствующие колебания в ЭЭГ.
Обосновано [6,8,10,13,14], что каждый смысловой квант должен начинаться с кратковременного синхронного торможения соответствующих модулей. Это обусловлено необходимостью накопления энергии структурных напряжений для осуществления акта нервной интеграции, и также может отображать "обнуление" нервных устройств перед каждым актом деятельности. Далее развертывается процесс нервной интеграции в виде характерной последовательности эпизодов миллисекундной синхронизации нервных спайков в больших ансамблях нейронов, т.е. - эпизодов синхронизированного возбуждения, каждый из которых сопровождается эпизодом синхронизированного торможения. Это означает, что число тормозных эпизодов миллисекундной синхронизации должно превышать число возбудительных ЭМС .
Мы предположили, что колебания ЭЭГ, связанные с торможением и возбуждением должны быть разнонаправлены так же, как и в фазах первичного ответа: торможению соответствует позитивность, а возбуждению - негативность.
Отмеченные повторяющиеся последовательности колебаний мы обозначаем как смысловые кванты ЭЭГ.
Мы обосновали [5,6,8,11,13,14], что эти кванты организованы в иерархию: более сложным процессам интеграции соответствуют более длительные кванты (временной интервал от волны торможения до волны очередного торможения нарастает, как, видимо, должны нарастать и средние интервалы между экстремумами волн возбуждения). Скажем, если мы будем двумя электродами отводить ЭЭГ в пределах проекционных полей одного анализатора мы будем сталкиваться с ЭМС (эпизодами миллисекундной синхронизации экстремумов волн ЭЭГ) более часто, чем тогда, когда один электрод будет отводить из поля анализатора, а другой - например из ретросплениальной коры, или электроды будут расположены в нескольких зонах коры, поскольку в этих случаях в ЭЭГ будут отображаться смысловые кванты более старшего ранга. Еще больших интервалов между ЭМС мы можем ожидать, для далеко отстоящих электродов, когда в ЭЭГ отображаются смысловые кванты еще более высокого ранга и т.д.
Соответственно нашему подходу длительность каждого смыслового кванта даже для одного и того же модуля меняется в зависимости от текущего функционального состояния. Аналогично - относительно временных отношений в иерархии квантов. Это определяет нелинейность биологического времени [1-3] и создает большие трудности для адекватного анализа ЭЭГ.
Действительно, прогресс в области анализа ЭЭГ определятся тем, насколько адекватно мы можем судить по ней о функциях мозга. Но на сегодня ЭЭГ (в рассматриваемом ракурсе) - это запись без начала и без конца, квантованная на шкале физического времени, которое не соответствует времени биологическому, к тому же ЭЭГ - это запись, никак не привязанная к иерархии смысловых квантов.
Соответственно изложенному выше возникает ощущение насущной необходимости поиска и анализа смысловых квантов ЭЭГ, прежде всего - поиска реперных событий, позволяющих разметить ЭЭГ на шкале событий нервной интеграции.
Цель настоящей работы - поиск моментов миллисекундной синхронизации экстремумов, т.е. позитивных и негативных ЭМС, а также закономерностей их представленности и повторяемости в записях в зависимости от локализации отводящих электродов в тех или иных зонах коры.
Методика
Опыты проведены на 7 морских свинках обездвиженных тубокурарином и переведенных в режим искусственного дыхания. Ректальная температура поддерживалась в диапазоне 36-37 град. При проведении операции использовали локальную новокаинизацию и эфирный наркоз. Отведения ЭКоГ осуществляли спустя час после
операции, когда эфир полностью выветривался за счет легочной вентиляции. Отведения осуществляли эпидурально от правой ретросплениальной коры, от симметричных зон проекционной зрительной коры и от симметричных зон проекционной соматосенсорной коры, а также от премоторной коры правого полушария - всего 6 точек отведения. Исследовали фоновые записи.
Сигналы через усилители спаренной установки УФПТ-5 через АЦП с шагом квантования 2мс вводились на ПЭВМ и оцифровывались по программе "EEG-vision", разработанной в нашем институте. Длина записи составляла 30с.
Для автоматического анализа ЭЭГ мы составили программу "WavletEEG", которая находила локальные экстремумы волн на записях по каждому из 6 каналов, и выявляла моменты синхронизации таких экстремумов на 2мс интервалах в любой паре каналов. Регистрировались два вида событий - момент совпадения положительных экстремумов (максимумов) и момент совпадения отрицательных экстремумов (минимумов).
По алгоритму на основе формулы Байеса программа проводила вычисление коэффициентов "неслучайности" совпадений для каждой пары каналов.
Далее программа выполняла построение постимпульсной гистограммы по стандартному алгоритму, применяемому при анализе импульсной активности нейронов. Импульсом (событием) в данном случае являлся момент совпадений экстремумов. По сути дела такая гистограмма показывала вероятность наступления события на расстоянии (по времени) от данного события.
На гистограммах (см. рис.1А и Б) по горизонтали - время, по вертикали - вероятность события. Возможно четыре варианта таких гистограмм: "плюс-плюс", "минус-минус", "плюс-минус", "минус-плюс". Гистограмма "плюс-плюс" - это вероятность совпадения максимумов ("плюсов") после предшествующего момента совпадения максимумов ("плюсов"). Гистограмма "минус-минус" - это вероятность совпадения минимумов ("минусов") после предшествующего момента совпадения минимумов ("минусов"). И т.д. Такую гистограмму можно рассматривать как аналог функции автокорреляции (или кросскорреляции), но только для импульсных потоков.
Анализ записей выполнялся в два этапа. На первом этапе после автоматического выявления моментов совпадений экстремумов (ЭМС) и оценки их "неслучайности" проводился визуальный контроль записей, на которых уже были проставлены метки совпадений (см. рис. 2). При этом из списка совпадений исключались моменты артефактов, которые могли быть на записях. Затем выполнялся повторный анализ с построением гистограмм.
Описанный метод лежал в основе исследования закономерностей пространственно-временной организации эпизодов синхронизации экстремумов соответствующих волн.
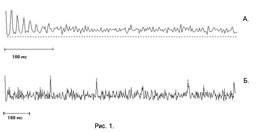
Постимпульсные гистограммы возбудительных ЭМС для отведений суммарной активности коры мозга морской свинки двумя электродами.
А - случай ритмического появления ЭМС с интервалом, соответствующим гамма-диапазону ЭКоГ.
Б - случай ритмического появления ЭМС с интервалом, соответствующим альфа-диапазону ЭКоГ.
Пояснения в тексте.
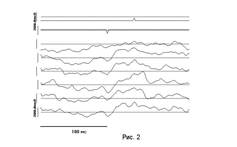
Выделение программными средствами отрицательных ЭМС - пик на канале 2 и положительных ЭМС - пик на канале 1.
З - канал отведения ЭКоГ премоторной коры, правое полушарие (ПП).
4 - то же - ретросплениальной коры, ПП.
5 - то же - зрительной, поле 17, ПП
6 - то же, левое полушарие (ЛП).
7 - канал отведения из SI, ЛП.
8 - то же, ПП.
Результаты
Вначале еще раз обратимся к рис.1А и Б на них видно, что эпизоды миллисекундной синхронизации (ЭМС) могут быть весьма закономерно организованы во времени, ложась на ритм гамма-волн (конкретно - около 80 Гц), но также на ритм волн иных диапазонов ЭКоГ: мы наблюдали альфа-диапазон -6-12Гц (рис 1. Б) и бета диапазон - до 30Гц, дельта диапазон -2-4Гц. Визуальный просмотр показал, что ЭМС характеризуют не столько миллисекундную синхронизацию "волн" ЭКоГ, но именно экстремумов - дискретных событий, разделенных временными интервалами. Как отмечалось - именно такие, дискретные и в общем случае апериодически следующие события в ЭЭГ мы рассматриваем в качестве информационного компонента ЭЭГ, но не "волны" (волн правильных в ЭЭГ нет вообще), последние на наш взгляд отражают динамику возбудимости нервных структур - модулей, того или иного уровня.
Действительно, проведенный анализ показал, что чаще всего ЭМС следовали апериодически, не вызывая соответствующих ритмически повторяющихся пиков на гистограммах (рис З). Это естественно, поскольку гистограмма построена в физическом времени, а биологическое время не линейно.
|
|
| Постимпульсные гистограммы тормозных ЭМС (вверху) и возбудительных (внизу) для отведений суммарной активности коры мозга морской свинки двумя электродами. Пример отсутствия ритмических осцилляций на шкале физического времени (особенно отчетливо - внизу) и пример (вверху) аритмичных ЭМС (дуплеты - смотри текст). |
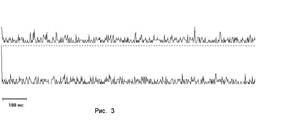
Особо отметим довольно обычные случаи отчетливых пиков ЭМС на гистограмме, которые не повторялись ритмически. Это соответствует ситуациям, когда, например, спаренные регулярным интервалом физического времени ЭМС - дуплеты - следуют друг за другом аритмично.
Поскольку длина смысловых квантов адекватно измеряется не в шкале физического времени, но в шкале событий, функциональных актов, которая существенно нелинейная [1-3] и может не соответствовать шкале физического времени, мы уделяли особое внимание другим способам анализа, кроме гистограммного.
В частности подсчитывалось абсолютное и процентное количество отрицательных - возбудительных и положительных - тормозных ЭМС экстремумов ЭКоГ попарно в пределах ипсилатеральных зон коры: премоторной, ретросплениальной, зрительной и соматосенсорной, а также количество ЭМС попарно в симметричных точках проекционных зон левого и правого полушарий, а также - для зон отведения из разных областей коры мозга.
Полученные результаты анализировали по соответствующим таблицам отдельно для каждого из семи опытов. Обнаружив, что картина распределения частоты проявления ЭМС существенно сходна в разных опытах (p< 0,05, критерий знаков) построили сводную таблицу.
Таблица. Пространственное распределение количества наблюдаемых эпизодов миллисекундной синхронизации возбудительных и тормозных экстремумов ЭКоГ (в %) по зонам коры мозга.
| Отведения ЭКоГ | 3 | 4 | 5 | 6 | 7 | 8 |
| 3 | - | 2,76 | 8,76 | 4,69 | 4,79 | 6,81 |
| 4 | 2,16 | - | 3,41 | 2,66 | 3,59 | 4,06 |
| 5 | 7,40 | 3,44 | - | 13,49 | 9,50 | 12,42 |
| 6 | 3,63 | 1,93 | 14,90 | - | 12,56 | 5,22 |
| 7 | 3,61 | 3,09 | 9,16 | 13,66 | - | 4,19 |
| 8 | 5,37 | 3,66 | 15,59 | 7,07 | 3,20 | - |
(+) n = 961
(-) n = 1033
Зоны отведения ЭКоГ: правое полушарие - 3,4,5,8; левое полушарие - 6,7;
3 - премоторная кора; 4 - ретросплениальная кора; 5 и 6 - зрительные проекционные зоны, 7 и 8 - соматосенсорные проекционные поля.
В таблице жирным шрифтом выделены достоверно отличающиеся значения (критерий Стьюдента для долей выборок[15]).
Представлены усредненные результаты по семи опытам.
Из таблицы прежде всего видно, что согласно ожиданиям (см. введение) число ЭМС тормозных экстремумов ЭКоГ превышало число возбудительных.
Далее - число ЭМС оказалось достоверно выше для случаев отведения внутри зрительного анализатора, чем для ЭМС между зрительным проекционным полем и ретросплениальной корой. При этом разница была особенно велика (p< 0,001, t-критерий для долей выборок) для контралатеральной зрительной зоны и ретросплениальной коры.
Несколько иная, чем для зрительной коры картина наблюдалась в симметричных полях соматосенсорной коры. Это, однако, можно объяснить ее сравнительно низким тонусом из-за обездвиживания тубокурарином.
То есть в целом представление о том, что для отведений внутри анализатора должны быть характерны более высокочастотные и короткие смысловые кванты ЭЭГ, чем для отведений из удаленных зон коры с их более длительными и низкочастотными смысловыми квантами (отображающими ансамбли старшего ранга), находится в разумном соотношении с результатами эксперимента.
Межанализаторные ЭМС для ипсиотведений оказались представлены даже большим числом, чем внутри анализаторные. Это неожиданно. Надо учесть, однако, низкую из-за тубокурарина активность соматосенсорной коры. Возможно, что у необездвиженного животного результаты будут другими. В общем это вопрос для дальнейших исследований.
Итак, данные таблицы показывают как минимум, что представленность и соответственно - частота следования ЭМС имеет функциональный смысл, в целом подтверждая разумность представления об иерархии смысловых квантов ЭЭГ.
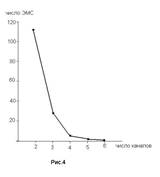
Наконец, мы сравнили число ЭМС в зависимости от числа одновременно анализируемых зон отведения. Полученная зависимость иллюстрируется рис.4. Эта зависимость вполне согласуется с предсказанной для иерархии смысловых квантов ЭЭГ (см. введение).
|
|
| Распределение числа наблюдаемых ЭМС , наблюдаемых с ростом числа одновременно анализируемых каналов. |
Заключение
Наш подход к вопросу о роли миллисекундной синхронизации различных ЭЭГ отведений принципиально отличается от сложившегося со времени открытия синхронизации гамма-осцилляций Сингером с коллегами, например [16] и последующего бума исследований и осмысления полученных результатов. Перечислять их нет смысла - их "тьмы и тьмы". Среди недавних отечественных обзоров с ясно выраженной оригинальной позицией отметим [17] - прежде всего потому, что он базируется на альтернативном нашему концептуальном подходе. И эта последняя работа в центр внимания ставит миллисекундную синхронизации прежде всего гамма осцилляций.
В противовес наш концептуальный подход не связывает образование ни микро , ни макро нейронных ансамблей, образующихся как код того или иного этапа нервной интеграции на миллисекундных временных интервалах исключительно с гамма осцилляциями, да и вообще - с наличием каких либо специфических осцилляций как непременного условия. И экспериментальные данные дают подтверждение именно этому взгляду на ЭЭГ феномены в миллисекундном диапазоне, подходу, учитывающему как нелинейность отображаемых в ЭЭГ процессов, так и их многоуровневость, а также их функциональную неоднозначность - наличие энергетического и информационного компонентов.
С другой стороны цель нашей работы, которая принесла лишь первые результаты - это создание метода адекватного шкалирования ЭЭГ. Такое шкалирование требует тщательного обоснования критериев. Разумное совпадение наших концептуально обоснованных предсказаний с фактами эксперимента убеждает в правильности выбранного направления работ, несмотря на их нетрадиционность. При этом остается впереди решение задачи идентификации начального (перед смысловым квантом ЭЭГ), тормозного колебания, которое будет "репером" смыслового кванта. Проблема состоит в том, чтобы отличить его от прочих тормозных ЭМС.
Подведем итоги.
Полученные результаты согласуются с концепцией дискретно-волнового механизма нервной интеграции и ее приложением к ЭЭГ - представлением об иерархии смысловых квантов ЭЭГ, по которому квазиритмические колебания - "волны" ЭЭГ есть ее энергетический компонент, колебания возбудимости, тогда как информационным компонентом ЭЭГ являются дискретные "пики" или экстремумы, отображающие апериодически следующие эпизоды формирования в коре мозга миллисекундных интегративных нейронных ансамблей разной сложности. Показано в частности, что кроме периодических эпизодов миллисекундной синхронизации (ЭМС) экстремумов ЭКоГ, которые могут быть разделены интервалами, соответствующими гамма, бета, сигма, альфа, дельта диапазонам ЭЭГ, ее "волнам", регулярно имеет место апериодическое следование ЭМС экстремумов, пиков ЭКоГ. Визуальное отслеживание записей ЭКоГ с должной временной разверткой показало миллисекундную синхронизацию именно "пиков", экстремумов ЭКоГ, но не ее "волн" - например, гамма осцилляций.
Показано, в соответствие с нашим концептуальным походом, что позитивных ЭМС, рассматриваемых по аналогии с волнами первичного ответа как тормозные, обнаруживаются в большем количестве, чем негативных ЭМС - возбудительных.
Показано, что "внеанализаторные" отведения характеризуются достоверно меньшим
числом ЭМС, чем внутрианализаторные; - это соответствует нашему
представлению об иерархии смысловых квантов ЭЭГ.
Проанализированы ближайшие проблемы исследования смысловых квантов ЭЭГ как метода адекватного шкалирования ЭЭГ с учетом нелинейности биологического времени.
Список литературы
Вернадский В.И. Пространство и время в неживой и живой природе - Размышления натуралиста -М.: Наука, 1975.
Владимирский Б.М. Информационные воздействия на нейрорегуляцию и поведение человека. - Научная мысль Кавказа, 1997, N 3, - С.45-53.
Владимирский Б.М. Собственное физиологическое время и неопределенность в нервной системе. - http://krinc.ru\Krinc\public\Articles
Чебкасов С.А. Конструкция интегративных модулей неокортекса и трансуровненвые принципы организации естественных информационно-управляющих систем // Ростов-на-Дону, НИИНК, 1980, Изд-во ЦВВР, 2003, - 158 c. ISBN 5-94153-059-5.
Чебкасов С.А. Материалы и заключения о функционировании простых декодирующих модулей зрительной коры. - Ростов-на-Дону,1991. Деп. ВИНИТИ N 3113 - В91. - 76 с.
Чебкасов С.А. Основные подходы и новые пути к сенсорной коррекции сна. - Отчет по теме N ГР 01910044497. Этап 1992г. - С. 33-89.
Chebkasov S.A. Desinchibition and stepwise synchronization of neural activity as mechanism of producing and encoding orientation signals within the functional modules in the visual cortex. - Proc. IJCNN-93, Nogaya, Japan, 1993. - V.1 - P. 113.
Чебкасов С.А. О возможных "смысловых" квантах ЭЭГ. - Успехи физиол. наук, 1994, N 4 С. 117.
Чебкасов С.А. Проблема сенсорного управления функциональным состоянием мозга с позиций концепции дополнительности. - Ростов-на-Дону, 1994. - Деп. ВИНИТИ N 1644 - В94.- 91 с.
Чебкасов С.А. Предстимульное торможение. - Физиол. журн. СССР, 1995, Т.81, N 5. С.119.
Chebkasov S.A. The phenomena of neuronal spike-s synchronization in the milliseconds range: purpose, mechanism, basic units. - The complementary approach - ICAUTO-95. Paper #110.
Chebkasov S.A. New attack on the memory-s problem-- Abstr. 4-th IBRO World Congress of Neuroscience. 1995, Kyoto, Japan. Publ. by rapid com. Oxford, New York, P.369, N D9.3.
Чебкасов С.А. Взаимоотношение специфической и неспецифической систем мозга. Дополнительный подход. - Деп. ВИНИТИ. - N 1938 - В97. - 34 с.
Чебкасов С.А. Концепция информационно-структурного дополнения: существо, нейрофизиологическое обоснование, перспективы. - Ростов-на- Дону, Изд-во ЦВВР, 2003. - 44 с. ISBN 5-94153-064-1.
Плохинский Н.А. Биометрия. - М: Изд-во МГУ, 1970, 368 с.
Friewald WA, Kreiter AK, Singer W. Synchronization and assembly formation in the visual cortex. - Prog Brain Res 2001; 130-40.
Кирой В.Н., О.Г. Чораян Нейронные ансамбли мозга. - Успехи физиол. наук, 2000, том 31,N3, с.23-38.
0 комментариев