Содержание
Оболочки головного мозга
Венозные синусы головного мозга
Артерии головного мозга
Общая характеристика строения головного мозга
Общая характеристика развития головного мозга
Концевой мозг- telencephalon
Развитие концевого мозга
Промежуточный мозг-diencephalon
Развитие промежуточного мозга
Средний мозг-mesencephalon
Развитие среднего мозга
Ромбовидный мозг-rhombencephalon
Развитие ромбовидного мозга
Список используемой литературы:
Оболочки головного мозга
Твердая оболочка головного мозга-dura mater encephali- самая наружная. В отличие от твердой оболочки спинного мозга она срастается с надкостницей черепных костей, поэтому эпидуральное пространство отсутствует. Между надкостницей и твердой мозговой оболочкой проходят лишь вены, образующие две системы венозных синусов - дорсальную и вентральную.
В субдуральное пространство от твердой мозговой оболочки отходят две складки: серповидная и перепончатый мозжечковый намет. Серповидная складка-falx cerebri- проходит сагиттально от петушьего гребня решетчатой кости до мозжечкового намета. Она лежит в продольной щели между полушариями большого мозга. Наиболее сильно серповидная складка выражена у собаки и лошади- она почти достигает мозолистого тела; она меньше у свиньи и коровы и особенно у мелких жвачных. Перепончатый мозжечковый намет-tentorium cerebella membranaceum- отходит оn костного намета или теменной кости, а также от гребней каменистых костей и располагается в поперечной щели между большим мозгом и мозжечком.
Паутинная оболочка головного мозга-arachnoidea encephali- на извилинах мозга прочно срастается с мягкой оболочкой. Подпаутинное пространство сохраняется только в щелях и бороздах между извилинами и на базальной поверхности мозга, где она образует расширения - вентральные цистерны продолговатого мозга, цистерну моста, червячка (и др.). Под паутинной оболочкой вдоль серповидной складки отходят в субдуральное пространство ворсинки, принимающие иногда форму узелков - пахионовы гранулы –granulationes arachnoidales. Пахионовы гранулы внедряются в сагиттальный синус и усиливают отток спинномозговой жидкости в вены.
Мягкая, или сосудистая, оболочка головного мозга-pia mater encephali- очень прочно срастается с мозгом, она заходит во все щели и углубления и вместе с сосудами проникает в вещество мозга. Вворачиваясь в полости мозга, она участвует в формировании сосудистых покрышек-tela chorioidea- в которых заложены сосудистые сплетения-plexus chorioideus. Такие образования находятся в желудочках мозга: боковых, третьем и четвертом.
Спинномозговая жидкость-liquor cerebrospinalis- заполняется субдуральное и субарахноидальное пространства головного и спинного мозга и через парные отверстия-apertura lateralis ventricularis quarti- каудально от боковых ножек мозжечка- и не парное отверстие-apertura mediana ventriculi quarti- сообщается с полостями желудочков и каналов мозга. Спинномозговая жидкость выделяется клетками эпендимы и сосудистых сплетений мозга. Она течет в субарахноидальном пространстве в сторону головного мозга, то есть краниально, а в центральном канале спинного мозга- каудально. Отток цереброспинальной жидкости происходит из субдурального пространства в венозную систему, а из субарахноидального пространства в органы лимфообращения.
Венозные синусы головного мозга
Дорсальная система синусов состоит из непарных сагиттального и прямого синуса и парных поперченных, затылочных и дорсальных каменистых; в них впадают вены мозга.
Сагиттальный синус-sinus sagittalis- находится в серповидной складке; назально он начинается из оболочечных вен, у лошади также из решетчатых и мозговых вен. В него впадают:
1)дорсальные вены мозга-vv.cerebri dorsales;
2)оболочечные вены из твердой мозговой оболочки;
3) костные вены-vv. diploicae;
4) короткий прямой синус-sinus rectus.
Последний образуется каудально от валика мозолистого тела, слиянием вены мозолистого тела с большой веной мозга.
Вена мозолистого тела-v. corporis callosi - выносит кровь из передней половины мозга.
Большая вены мозга-v. cerebri magna-принимает в себя глубокие вены мозга-vv. cerebri profundi- выносящие кровь из мозга и его сосудистых сплетений.
Каудально от впадения прямого синуса, сагиттальный синус делится на правый и левый поперечные синусы,-sinus transversus - каждый из которых направляется в височный канал и переходит в дорсальную мозговую вену-v. cerebri dorsalis. Последняя впадает в височную поверхностную вену. В поперечный синус, до его погружения в височный канал, вливается каменистый синус-sinus petrosus- который выносит кровь из базальных частей мозга.
Между поперечными синусами находится соединительный синус-sinus communicans- он лежит в основании мозжечкового намета. В него впадают затылочные синусы-sinus occipitales- помещающиеся в бороздах по обе стороны червячка мозжечка.
Отток крови из дорсальной системы синусов, помимо мозговых дорсальных вен, происходит так же и через эмиссарии-emissarium- в височные глубокие вены. Эмиссарии- это непостоянные отводящие каналы, прободающие стенку черепной полости.
Вентральная, или базилярная, система синусов включает синусы циркулярные и базилярные.
Циркулярный синус-sinus circularis-окружает гипофиз. Он образован правым и левым кавернозными синусами, соединяющимися межкавернозными синусами-sinus cavernosus et intercavernosus.
Каждый кавернозный синус назально переходит в глазничную вену мозга каудально- в базилярный синус. Глазничная вена мозга-v. cerebralis orbitalis- впадает через глазничную щель в глубокую лицевую вену.
Базилярный синус-sinus basilaris- вливается вентральный позвоночный синус. В области рваного отверстия он отдает вентральную и мозговую вену-v. cerebralis ventralis- которая впадает в затылочную вену. Близ затылочного сустава базилярные синусы, анастомозируя между собой, образуют вентральный затылочный синус-sinus occipitalis ventralis. В базилярную систему синусов открываются вентральные вены мозга.
Артерии головного мозга
1.Внутренняя сонная артерия-a. carotis interna -проникает в черепную полость через сонное или рваное отверстие и делится на назальную и каудальную соединительные ветви. С одноименными ветвями другой стороны они формируют вокруг гипофиза артериальное кольцо-circulus arteriosis cerebri. Назально из кольца выходит назальная артерия-a. cerebri nasalis- она направляется к мозолистому телу и питает передние отделы полушарий.
От назальной соединительной ветви ответвляются 4 сосуда:
а) назальная артерия мозговой оболочки-a. meningea nasalis;
б) средняя мозговая артерия-a. cerebri media-идет на боковую поверхность мозга;
в) назальная артерия сосудистого сплетения-a. chorioidea nasalis- по зрительному тракту следует в сосудистое сплетение бокового желудочка;
г) внутренняя глазничная артерия-a. ophthalmica interna- по зрительному нерву выходит в глазницу.
От каудальной соединительной ветви отходят:
а) каудальная мозговая артерия-a. cerebri caudalis- для задних отделов полушарий и четверохолмия
б) каудальная артерия сосудистого сплетения-a. chorioidea caudalis-в сосудистое сплетение боковых желудочков.
2. От затылочной артерии отходит спинномозговая артерия-a. cerebrospinalis- через межпозвоночное отверстие атланта она проникает в позвоночные канал и делится на краниальную и каудальную ветви, соединяющиеся с одноименными ветвями другой стороны. От краниальных ветвей в головной мозг отделяется основная артерия мозга-a. basilaris cerebri- она впадает в артериальное кольцо, а на своем пути отдает: позади моста- каудальную артерия мозжечка, впереди моста- краниальную артерию мозжечка-a. cerebella caudalis et cranialis- а к слуховому нерву- внутреннюю слуховую артерию-a. auditiva interna. Каудальная ветвь спинномозговой артерии анастомозирует со спинномозговой вентральной артерией-a. spinalis ventralis. Сосуды мозга иннервируются симпатической нервной системой.
Общая характеристика строения головного мозга
Головной мозг- encephalon- с дорсальной поверхности разделяется поперечной щелью-fissura transversa cerebri- на большой и ромбовидный мозг.
Большой мозг-cerebrum-состоит из двух полушарий, промежуточного и среднего мозга. Правое и левое полушария большого мозга-hemisphaerum cerebri dextrum et sinistrum- дорсально разграничиваются глубокой продольной щелью-fissura longitudinalis cerebri. Промежуточный и средний мозг дорсально прикрыты полушариями.
Ромбовидный мозг-rhombencephalon-состоит из мозжечка, мозгового моста, объединяемых в задний мозг, и продолговатого мозга.
Мозжечок-cerebellum- располагается дорсально от продолговатого мозга и сзади от полушарий большого мозга.
Продолговатый мозг-medulla oblongata- является непосредственным продолжением спинного мозга. На переднем конце его расположен мозговой мост-pons cerebri.
Впереди моста находятся ножки большого моста. Спереди ножки простираются до зрительных трактов-tractus opticus- и перекреста зрительных нервов-chiasma opticum. Между ножками большого мозга, позади зрительного перекреста, лежат: сосцевидное тело-corpus mamillare- и серый бугор с воронкой и гипофизом. Впереди зрительных трактов и с боков от ножек большого мозга выступает обонятельный мозг-rhinencephalon- орально он заканчивается двумя обонятельными луковицами-bulbi olfactorii. Обонятельный мозг представляет вентральный отдел полушарий большого мозга, а покрытый бороздами и извилинами плащ-pallium- дорсо-латеральный их отдел.
Полушария относятся к концевому мозгу-telencephalon- а зрительный тракты, серый бугор, воронка, гипофиз и сосцевидное тело - к вентральным отделам промежуточного мозга-diencephalon.
Ножки большого мозга являются вентральной частью среднего мозга-mesenecephalon.
Мозжечок и мост входят в состав заднего мозга-metencephalon. Промежуточный, продолговатый и средний мозг объединяются в ствол большого мозга-truncus cerebri.
Общая характеристика развития головного мозга
Головной мозг развивается в связи с возникновением и дальнейшим развитием различных высших нервных центров: рецепторных, моторных и ассоциативных, которые обеспечивают согласованную функцию всех органов - соматических, висцеральных, сосудистых.
На ранних стадиях онтогенеза и филогенеза головной мозг представляет собой расширенный конец мозговой трубки, который лежит впереди хорды, поэтому он называется прехордальным или первичным мозговым пузырем. Развитие его связано с органом обоняния, и его обонятельная функция сохраняется у всех животных, включая млекопитающих.
Несколько позже на переднем конце эпихордального мозга обособляется прехордальный, или вторичный, мозговой пузырь. Развитие его обусловлено: возникновением первичного ассоциативного и комиссурального центра; наличием жаберного аппарата и органов боковой линии водных животных; дифференцировкой внутренних органов.
От передней части эпихордального мозгового пузыря обособляется средний мозговой пузырь, который, исключая млекопитающих, выполняет ведущую роль, поскольку в него поступают импульсы решительно от всех рецепторных аппаратов и прежде всего зрительных.
С дорсальной поверхности все три отдела мозга довольно резко отделяются друг от друга поперечными комиссурами из нервных волокон, впереди и позади среднего мозга.
В дальнейшем и прехордальный мозг разделяется на концевой мозг в виде парного пузыря и промежуточный мозг.
Концевой мозг наивысшей стадии своего развития достигает у млекопитающих животных с наличием новой коры большого мозга, которая становится высшим нервным центром.
В промежуточном мозге у низших животных в связи с ведущей ролью обонятельного анализатора первое место занимают гипоталамус и эпиталамус. Они являются промежуточными обонятельными центрами, посылающими импульсы в средний мозг, а гипоталамус, кроме того, посылает импульсы в гипофиз и в автономные центры среднего и продолговатого мозга. В силу этого гипоталамус становится высшим подкорковым вегетативным центром. Эту функцию он охраняет и у млекопитающих животных.
Лишь с формированием коры большого мозга у млекопитающих усиленно начинает развиваться средний участок промежуточного мозга - таламус, через который кора получает решительно все рецепторные импульсы с периферии.
Таким образом, исторически гипоталамус более связан с эпихордальным мозгом, а таламус с прехордальным.
Почти одновременно с делением прехордального мозга дифференцируется и эпихордальный мозг позади среднего мозгового пузыря на задний мозг и продолговатый мозг. Задний мозг до млекопитающих представлен одним лишь мозжечком, который является подкорковым центром корреляции мышечных движений для сохранения равновесия. Лишь у млекопитающих к мозжечку вследствие развития новой коры большого мозга добавляется мозговой мост, а сам мозжечок усиливается двумя полушариями мозжечка.
Полости первичных мозговых пузырей становятся в развитом мозге мозговыми желудочками. Из полости прехордального мозга в концевом мозге возникают парные боковые желудочки, а в промежуточном мозге- третий мозговой желудочек. Все три желудочка соединяются межжелудочковым отверстием. Желудочек среднего мозгового пузыря вследствие разрастания стенок последнего превращения в мозговой водопровод, а полость ромбовидного мозга становится четвертым мозговым желудочком, который сообщается с центральным спинномозговым каналом.
Концевой мозг- telencephalon
Состоит из двух полушарий большого мозга-hemisphaerum dextrum et sinistrum-, разделенных с дорсальной поверхности глубокой продольной щелью-fissura longitudinalis cerebri.
В каждом полушарии рассматривают плащ, обонятельный мозг, полосатые тела и боковые желудочки мозга. Плащ расположен в полушарии дорсо-лательно от желудочка, обонятельный мозг – вентро-медиально. Границей между плащом и обонятельным мозгом на вентральной поверхности мозга служит базальная пограничная борозда. Полосатое тело лежит в вентральной стенке полушария дорсально от частей обонятельного мозга.
А) Плащ -pallium
Состоит из серого и белого мозгового вещества. Серое мозговое вещество-substantia grisea- образует кору большого мозга-cortex cerebri- на его поверхности. На ней различают извилины мозга-girus - разделенные бороздами и щелями. На латеральной поверхности плаща видны три дугообразные извилины. Они окружают латеральную сильвиеву борозду-sulcus lateralis. Дугообразные извилины четко выражены у собаки, а у других животных каждая дуговая извилина подразделяется бороздами на части. На медиальной поверхности плаща в основном заметны две дугообразные извилины вокруг мозолистого тела, причем ближайшая к нему поясная извилина-girus cinguli- функционально связана с обонятельным мозгом.
Белое мозговое вещество плаща-substantia alba- располагается под корой плаща. Оно состоит из проводящих путей - ассоциативных, комиссуральных и проекционных.
а) ассоциативные волокна соединяют отдельные участки коры в пределах каждого полушария. Они разделяются на короткие волокна и длинные.
б) комиссуральные волокна соединяют участки, принадлежащие разным полушариям. Они формируют мозолистое тело-corpus callosum- самую крупную комиссуру головного мозга. Оно помещается между полушариями в глубине продольной щели. Различают ствол мозолистого тела-truncus corporis callosi-и два конца - передний и задний. Передний конец называется коленом мозолистого тела-genu corporis callosi- он загибается вентрально. Задний конец, или валик, мозолистого тела-splenium corporis callosi- срастается со сводом.
Комиссуральные волокна, выходящие из ствола мозолистого тела, образуют лучистость спайки-radiatio corporis callosi- она формирует дорсальную стенку бокового желудочка мозга и расходится в передний, боковой и задний отделы коры плаща.
в) проекционные волокна - соединяют кору плаща, как с отдельными частями ствола головного мозга, так и со спинным мозгом. Они образуют в полосатом теле внутреннюю капсулу. Функционально проекционные пути разделяются на эфферентные и афферентные.
Эфферентные проводящие пути выносят импульсы из коры полушарий большого мозга в разные отделы ствола мозга: большого, ромбовидного и спинного.
Афферентные проводящие пути приносят импульсы в кору полушарий из спинного и ромбовидного мозга. Они образуют заднюю часть внутренней капсулы.
В противоположность коре больших полушарий все серое вещество остальных отделов всей центральной нервной системы объединяется понятием «подкорка». Импульсы со всех частей тела сначала идут в разные отделы подкорки, включая зрительный бугор, а из последнего поступают в кору полушарий. Кора постоянно разлагает, расчленяет возбуждения, возникающие в рецепторных приборах, и в то же время объединяет, синтезирует действие выделенных анализом возбуждений в разнообразные комплексы. Это и представляет собой проявление аналитической и синтетической деятельности коры головного мозга. Более того, высший отдел нервной системы - кора большого мозга является (у человека) материальным субстратом высшей нервной деятельности: мышления, сознания, памяти, рассудка, воли, то есть всей психической деятельности, которая представляет собой результат физиологической деятельности коры головного мозга.
Б) Обонятельный мозг-rhinencephalon
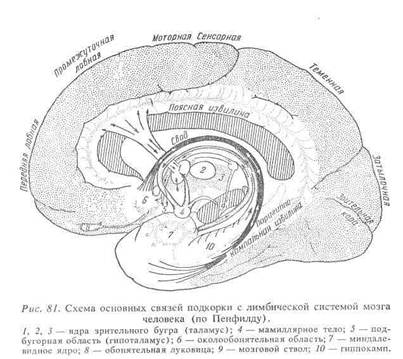
Располагается в вентро - медиальном отделе каждого полушария большого мозга. Отдельные части его видны на базальной и медиальной поверхностях полушарий, а также на дне боковых желудочков мозга. На базальной поверхности полушарий располагаются обонятельные луковицы, обонятельные тракты и извилины, обонятельные треугольники и грушевидные доли. На медиальных поверхностях полушарий видны околообонятельное поле, извилины гиппокампа, поясные извилины и поверхность разреза назальной спайки, а на дне боковых желудочков мозга- хвостатые ядра, гиппокамп и свод. Поясничная извилина, гиппокамп, свод и миндалевидное ядро составляют лимбическую систему-pars limbica rhinencephalon.
1.Обонятельная луковица-bulbus olfactorius- парное образование в виде довольно плоского, вытянутого и загнутого дорсально мозгового отростка, который выдается за передний край полушария мозга в обонятельную ямку решетчатой кости. Дорсомедиальный отдел луковицы построен из серого мозгового вещества, а латеро - вентральный – из белого мозгового вещества. В луковице находится желудочек обонятельной луковицы-ventriculus bulbi olfactorii- он является продолжением бокового желудочка мозга. В обонятельную луковицу входит обонятельный нерв-n. olfactorius. Он содержит многочисленные пучки нервных волокон –fila olfactoria- направляющихся от обонятельных клеток слизистой оболочки носа к нервным клеткам луковицы. Таким образом, обонятельные луковицы являются первичными обонятельными центрами.
2. От нервных клеток обонятельной луковицы начинаются обонятельные проводящие пути. Они образуют белое мозговое вещество самой луковицы и обонятельные тракты - общий, медиальный и латеральный-tractus olfactorius communis, medialis et lateralis. Латеральный обонятельный тракт переходит на грушевидную долю, покрывая на всем своем пути латеральную обонятельную извилину-girus olfactorius lateralis. Медиальный обонятельный тракт достигает медиальной поверхности плаща, формируя околообонятельное поле-area parolfactoria. Обонятельные тракты ограничивают обонятельный треугольник-trigonum olfactorium- из серого вещества. Они проводят импульсы от обонятельной луковицы к клеткам вторичных обонятельных центров в обонятельных извилинах, обонятельных треугольниках, околообонятельных полях, в грушевидных долях, а также в гипоталамус и в средний мозг.
3.Грушевидная доля-lobus periformis- расположена медиально от латерального обонятельного тракта и каудально от обонятельного треугольника; медиально граничит с ножками большого мозга. Каудо-медиальной границей грушевидной доли является медиальная пограничная щель, или щель гиппокампа-fissura hippocampi. В грушевидной доле заключена полость, представляющая собой задний отдел бокового желудочка мозга.
Грушевидная доля каудально переходит без четкой границы в извилину гиппокампа, расположенную на медиальной поверхности полушария, сзади и латерально от щели гиппокампа.
Извилина гиппокампа дорсально продолжается в поясную извилину-girus cinguli. Последняя проходит непосредственно дорсально от мозолистого тела,и огибая его спереди, соединяется с околообонятельным полем и представляет собой часть коры плаща. Грушевидная доля является вторичным обонятельным центром.
4.Гиппокамп, или аммонов рог-hippocampus- своим дорсальным отделом образует дно бокового желудочка мозга, позади хвостатого ядра, от которого отделяются сосудистым сплетением бокового желудочка. Гиппокамп представляет собой складку коры головного мозга в области щели гиппокампа и грушевидной доли. Он серповидно изгибается латер0-каудально и вентрально и теряется в стенке грушевидной доли. Аммоновы рога лежат дорсально на зрительных буграх, будучи отделены от них сосудистым сплетением третьего мозгового желудочка. Являясь высшими ассоциационными подкорковыми обонятельными и вкусовыми центрами, аммоновы рога связаны с различными участками коры больших полушарий и подкорковыми ядрами. Проводящие пути их образуют свод и его производные.
5. Свод -fornix- содержит в своем составе проводящие пути, соединяющие гиппокамп с сосцевидным телом промежуточного мозга. Отдельные участки этого пучка волокон образуют желобоватый листок, кайму аммонов рога, ножки, столбы, тело и переднюю спайку мозга.
Желобоватый листок-alveus hippocampi- покрывает гиппокамп с его поверхности, обращенной в боковой желудочек мозга. Он образован нервными волокнами, происходящими из серого мозгового вещества грушевидной доли и гиппокампа. По дорсолатеральному краю эти волокна создают кайму гиппокампа-fimbria hippocampi- продолжающуюся назально в ножку свода-crus fornicis. Последняя, соединяясь с ножкой с другой стороны, становится коротким телом свода-corpus fornicis. Тело свода служит дорсальной стенкой третьего мозгового желудочка; назально оно разделяется на два столба свода-columnae fornicis. Последние медиально от хвостатых ядер направляются к сосцевидному телу и серому бугру гипоталамуса. Комиссура свода-comissura fornicis- образована поперечными волокнами между ножками свода; она соединяет дорсальные концы аммоновых рогов друг с другом.
6. Между передним концом мозолистого тела и столбами свода находится передняя спайка мозга-comissura nasalis. Она соединяет друг с другом обонятельный мозг обоих полушарий.
В) Полосатое тело-corpus striatum
Расположено впереди зрительных бугров, дорсально от обонятельных треугольников, состоит из четырех ядер: хвостатого, чечевицеобразного, миндалевидного и ограды. Между ядрами находятся внутренняя и наружная спайка капсулы, образованные проекционными проводящими путями.
На хвостатом ядре-nucleus caudatus- различают головку и хвост. Головка хвостатого ядра своей дорсальной поверхностью образует дно бокового желудочка, назо -латерально от гиппокампа.
Чечевицеобразное ядро-nucleus lentiformis- располагается латерально от головки хвостатого ядра и зрительного бугра и отделяется о них внутренней капсулой-capsula interna. Латеральная часть чечевицеобразного ядра называется скорлупой-putamen. Медиальная часть чечевицеобразного ядра более древнего происхождения, называется бледным ядром-globus pallidus. Оно связано с обонятельным мозгом, лежит латерально от зрительного бугра и впереди от ядра латерального коленчатого тела-ganglion geniculatum. Латерально от чечевицеобразного ядра находится наружная капсула-capsula externa- а латерально от капсулы -другое ядро виде узкой полоски -ограда-claustrum. На границе между оградой, скорлупой и аммоновым рогом располагается миндалевидное ядро-nucleus amigdale- которое является обонятельным мозгом.
Полосатые тела соединяются проводящими путями: 1) с корой большого мозга; 2) со зрительными буграми и гипоталамусом; 3)с ядрами сетчатого образования среднего мозга, с ядрами моста и продолговатого мозга; 4) с ядрами черепно-мозговых нервов.
Через полосатые тела замыкаются различные рефлекторные цепи или:
А) периферический рецепторный аппарат - зрительные бугры - полосатые тела - соматические и висцеральные эффекторные аппараты.
Б) кора - полосатое тело - соматические и висцеральные эффекторные аппараты.
У млекопитающих ядра полосатых тел являются важнейшими подкорковыми двигательными центрами:
1) Координированных непроизвольных движений;
2) Регуляции мышечного тонуса в состоянии покоя и движения;
3) Безусловных рефлексов;
4) Высшими подкорковыми вегетативными центрами
Полосатые тела функционируют как единое целое, но отдельные их части действуют противоположно.
Развитие концевого мозга
Из более древних передней и вентральной стенок концевого мозга происходят обонятельные доли и полосатые тела, а из более поздней дорсальной его стенки - плащ.
1. Развитие обонятельных долей мозга обуславливается наличием обонятельного анализатора, который у водных животных имеет исключительное значение при ориентации во внешней среде. Отростки чувствительных обонятельных клеток заканчиваются в обонятельных луковицах, составляющих часть обонятельных долей концевого мозга, проксимальные отделы которых формируют древнейшую обонятельную кору-palaeocortex- из серого мозгового вещества. У млекопитающих она образует обонятельные извилины и обонятельные треугольники-trigonum olfactorium- являющиеся вторичными обонятельными центрами.
2. В вентральной стенке концевого мозга, дорсально от обонятельной коры очень рано появляется крупноклеточный базальный ганглий-ganglion basale. Это высший двигательный центр. Базальный ганглий сохраняется и у млекопитающих животных в виде бледного ядра. Позднее у наземных животных разрастаются дополнительные мелкоклеточные ядра, образующие в целом -neostriatum- который состоит из скорлупы, а у млекопитающих еще хвостатого ядра. Кроме того, обособляются новые ядра - ограда и миндалевидное ядро. С появлением вторичного плаща сложный базальный ганглий пронизывается внутренней и наружной капсулами из проводящих путей, идущих в кору плаща и обратно. В силу этого и возник новый термин -полосатое тело.
3. В эволюции плаща наблюдаются две формации различной функции и строения, не считая мембранообразного примитивного плаща-pallium membranaceum- характерного для водных животных и состоящего из эпендимы. У наземных животных сначала возникает кора более простой, трехслойной, клеточной структуры. Из нее формируются высшие обонятельные центры: грушевидные доли, извилины гиппокампа и их производные. Все эти части объединяются понятием первичный плащ-arhipallium.
Первичный плащ характеризуется расположением серого мозгового вещества под белым мозговым веществом. Архипаллиум, также как и палэопаллиум, лучше всего выражен у макросматических животных-macrosmates- обладающих острым обонянием, в отличие о микросматов-microsmates- у которых орган обоняния менее развит.
У млекопитающих на границе между обонятельной долей и первичным плащом появляется вторичный плащ-neopallium. Вторичный плащ характеризуется поверхностным расположением серого мозгового вещества в виде коры над белым мозговым веществом. Зачатки его появляются еще у рептилий. По мере своего разрастания новый плащ оттесняется от арихипаллуим в медиальную сторону и вентрально. При этом в области щели гиппокампа образуется складка коры в виде гиппокампа, а сам неопаллиум занимает всю дорсолатеральную часть полушария и, распространяясь каудально, покрывает промежуточный и даже средний мозг с их дорсальной и боковых поверхностей.
Разрастание вторичного плаща вызывается внедрением в него новых проекционных нервных волокон из промежуточного мозга, являющихся проводниками различных анализаторов- кожного, зрительного, слухового, мышечного и развитием новых ассоциативных нейронов и центров, включая сетчатое образование.
Кора вторичного плаща в ряду животных чрезвычайно усложняется в своем строении, резко дифференцируются ее функции и структура и увеличиваются размеры. У крупных млекопитающих животных плащ обычно несет извилины и борозды. Такие животные называются-gyrencephali. Извилины в ряду животных располагаются неодинаково. В одних случаях они в основном идут дугами вокруг поперечной сильвиевой борозды; у приматов извилины образуют две системы - лобную и теменную. Обе системы разделены сильвиевой бороздой. У третьей группы животных сильвиева борозда отсутствует, а борозды идут в передней части мозга, продольно, а в задней поперечно. Поэтому гомологизация извилин между животными разных отрядов крайне затруднительна, а в отдельных случаях может быть и невозможна.
У мелких животных извилины вообще отсутствуют. Наибольшее число извилин насчитывается у слонов и китов. В онтогенезе извилины появляются также не сразу, а в определенной последовательности.
Промежуточный мозг-diencephalon
Располагается позади полосатых тел концевого мозга и впереди среднего мозга, дорсально прикрыт сосудистой покрышкой третьего мозгового желудочка и гиппокампом. Промежуточный мозг состоит из трех отделов различного происхождения, строения и функции: эпиталамуса, таламуса и гипоталамуса. Эпиталамус образован сосудистой покрышкой третьего мозгового желудочка, эпифизом и парным узлом уздечки. Таламус состоит из зрительных бугров, между которыми находится кольцевидной формы третий желудочек мозга. Гипоталамус составляют зрительный выступ с концевой пластинкой, серый бугор с воронкой и гипофизом и сосцевидное тело. Все части гипоталамуса видны на базальной поверхности мозга, между ножками большого мозга, позади перекреста зрительных нервов.
Таламус. Зрительные бугры-thalami optici- самая массивная часть промежуточного мозга. Назо-латерально они срастаются с хвостатыми ядрами полосатого тела; зрительные бугры отделятся от последних пограничной полоской-stria terminalis-, от четверохолмия - поперечной бороздой, а друг от друга - ямкой зрительных бугров, прикрытой сосудистой покрышкой третьего желудочка. Бугры составляют многочисленные скопления ядер серого мозгового вещества. Наиболее крупные из них:
1.назальное ядро-nucleus nasalis thalami- лежит в толще назального бугорка-tuberculum nasalis thalami- в назо-медиальном отделе зрительного бугра. Оно наиболее древнее и является центром переключения афферентных путей на рефлекторные пути.
2.каудальное ядро-nucleus caudalis thalami- заключено в толщу каудо-латерального отдела латерального бугорка-tuberculum lateralis thalami- и состоит из промежуточных зрительных и слуховых центров. От перекреста зрительных нервов и слуховых центров. От перекреста зрительных нервов-chiasma opticum- на базальной поверхности мозга начинаются зрительные тракты-tractus opticum. Каждый тракт огибает латерально таламус и переходит в латеральное коленчатое тело-corpus geniculatum lateralis- теряющееся в каудальном ядре зрительных путей, идущих в кору большого мозга. Между латеральным коленчатым телом и четверохолмием выступает медиальное коленчатое тело-corpus geniculatum medialis. Оно соединяет каудальные холмы четверохолмия с каудальным ядром зрительного бугра и является промежуточным слуховым центром на пути в кору. Оба коленчатых тела объединяются в 7.
3. латеральное ядро-nucleus lateralis thalami- является центром переключения проводящих путей кожного анализатора и суставно-мышечного анализатора на проводящие пути, идущие в кору большого мозга и в полосатое тело.
4. медиальное ядро-nucleus medialis thalami- представляет собой промежуточный двигательный центр для проводящих путей из коры в ядра экстрапирамидной системы: в красное ядро, в ядра черепно-мозговых нервов и в спинной мозг.
5. сетчатое образование-formatio reticularis- располагается между ядрами и связано с ними.
Третий желудочек мозга-ventriculus tertius-находится между зрительными буграми, имеет кольцевидную форму, так как в него прорастает промежуточная масса зрительных бугров-massa intermedia thalami. В стенках желудочка находится центральное серое мозговое вещество-substantia grisea centralis- в нем располагаются подкорковые вегетативные центры. Третий желудочек сообщается с мозговым водопроводом среднего мозга, а позади назальной спайки мозга-comissura nasalis- с боковыми желудочками мозга через межжелудочковое отверстие-foramen interventriculare.
Эпиталамус. По краям ямки зрительных бугров видны мозговые полоски зрительных бугров-stria medullaris- а на них – парный узел уздечки-ganglion habenula. Узел уздечки переходит в уздечку или поводок-habenula- на нем укреплен эпифиз-epiphysis- грушевидной формы. Эпифиз-железа внутренней секреции, лежит в ямке между зрительными буграми и четверохолмием. Ганглий уздечки служит промежуточным центром для рефлекторных путей между мозгом, ядрами 5 пары и межножковым ядром.
Сосудистая покрышка третьего мозгового желудочка-tela chorioidea ventriculi tertii- образована складкой эпителиальной пластинки мягкой оболочки мозга и сосудистым сплетением. Эпителиальные пластинки покрышки прикрепляются по краям ямки зрительных бугров и свода. Сосудистая покрышка отделяет зрительные бугры от аммоновых рогов и от свода, она проникает через межжелудочковое отверстие в боковые желудочки мозга в виде сосудистых сплетений боковых желудочков мозга-plexus chorioideus ventriculi lateralis. Сосудистая покрышка формирует впереди эпифиза и непосредственно позади валика мозолистого тела выступ-recessus suprapinealis.
Гипоталамус - подталамическая, более древняя часть-hypothalamus- образует стенку третьего желудочка вентрально от зрительных бугров, является высшим подкорковым вегетативным центром, соединяется со зрительными буграми, с обонятельным мозгом и со средним мозгом. Гипоталамус содержит центры регуляции: дыхания, кроволимфообращения, температуры, обмена веществ, половых функций. Передний отдел его представлен серым бугром, воронкой и гипофизом, а задний- сосцевидным телом и стенками третьего желудочка.
Серый бугор гипоталамуса-tuber cinereum- лежит непосредственно позади перекреста зрительных нервов, между ножками большого мозга. В центра серого бугра имеется бухта воронки-recessus infundibuli. Сама воронка-infundibulum- тонкостенная, к ней прикрепляется гипофиз.
Гипофиз-hypophysis- представляет собой плоскоокруглое тело сложного строения с небольшой центральной полостью. Гипофиз состоит из трех частей: мозговой, промежуточной и железистой. Он является важнейшей железой внутренней секреции, выделяющей разнообразные гормоны и регулирующей все остальные железы внутренней секреции, симпатическую и парасимпатическую нервную системы.
Сосцевидное тело-corpus mamillare- лежит непосредственно позади серого бугра и служит промежуточным рефлекторным обонятельным центром, который через комплекс образований свода соединяется с обонятельным мозгом. Кроме того, сосцевидное тело связано со зрительными буграми и с сетчатым образованием среднего мозга. У собаки сосцевидное тело парное.
Развитие промежуточного мозга
Промежуточный мозг закладывается из небольшого числа клеток в стенке обширного третьего желудочка мозга, лишь у наземных животных. И особенно у млекопитающих, он достигает значительного размера.
1.хороо выраженная эмбриональная пластинка покрышки у всех взрослых млекопитающих формирует эпиталамус. Эпифиз является рудиментом третьего непарного теменного глаза. Лишь у некоторых водных животных и у рептилий имеется глазообразный пузырь под кожным покровом. У млекопитающих эпифиз становится железой внутренней секреции, функция которой еще недостаточно изучена.
2.своеобразно и сильно развитая эмбриональная пластинка дна формирует подталамическую часть промежуточного мозга. Она происходит из эпихордального мозга, то есть связана со средним мозгом.
Гипофиз происходит из трех разных источников. Эктодерма глотки превращается в разветвленную железу. Просвет железы впоследствии исчезает, но остаются тяжи из железистых клеток разнообразного строения, окруженные большим числом сосудов. У наземных животных из стенки воронки возникает нервная часть гипофиза из нервных и глиальных клеток и нервных волокон. Обособляется промежуточная часть гипофиза из эпителиальных клеток. Гипофиз в целом выделяет свыше десятка разнообразных гормонов, которые из железистой части поступают в кровь, а из нервной и промежуточной частей - в спинномозговую жидкость. Гипофиз находится во взаимодействии с вегетативными центрами, заложенными в стенках третьего желудочка мозга.
3.эмбрональная боковая пластинка образует зрительные буры-thalamus - и глазные пузыри. В состав таламуса входят ядра зрительных бугров. Они служат:
а) промежуточными центрами всех проводящих путей, которые направляются в кору полушарий и проводят различные импульсы - обонятельные, кожной и мышечной чувствительности, вкусовые, а у наземных животных, кроме того, зрительные и статоакустические;
б) промежуточным центром всех проводящих путей, идущих из коры плаща в разные отделы мозга.
Этим объясняется, почему зрительные бугры начинают формироваться с появлением обонятельного плаща и достигают максимального развития у млекопитающих в связи с образованием вторичного плаща. Мощное развитие зрительных бугров обусловлено также перемещением зрительных центров из среднего мозга во вторичный плащ, связью с мозжечком. В результате обогащения таламуса промежуточными центрами разрастается промежуточная масса зрительных бугров, которая внедряясь в полость третьего желудочка мозга, превращает его в кольцевидный канал. В сером веществе стенки третьего мозгового желудочка находятся многочисленные высшие подкорковые вегетативные центры.
Средний мозг-mesencephalon
Состоит из: пластинки четверохолмия, покрышки ножек, или чепца, из ножек большого мозга. Полость среднего мозга превратилась в мозговой водопровод-aqueductus cerebri- он соединяет третий и четвертый мозговые желудочки. В стенках водопровода заложено центральное серое мозговое вещество покрышки.
1.пластинка четверохолмия-tectum- представляет дорсальную часть среднего мозга, она лежит каудально от зрительных бугров и назально от мозжечка. Пластинка состоит из парных назальных и каудальных холмов-colliculi nasalis et caudalis. Холмы разделяются поперечной и срединной бороздами. С поверхности пластинка четверохолмия покрыта белым мозговым веществом, под которым находится серое мозговое вещество, в назальных холмах оно является подкорковым центром разнообразных афферентных путей, в то числе и зрительных, а в каудальных- подкорковым слуховым и равновесным центром.
В целом пластинка четверохолмия является координирующим центром ряда импульсов: обонятельных, зрительных, равновесно-слуховых, общей чувствительности и импульсов из коры больших полушарий. Двигательные импульсы передаются в красное ядро, в спинной мозг, а также к главным мышцам, в мозжечок и в мост.
2.в центре среднего мозга между пластинкой четверохолмия и ножками большого мозга находится покрышка ножек или чепец-tegmentum pedunculi. В нем залегают парные ядра серого мозгового вещества, в плоскости передних холмов лежат:
а) красное ядро-nucleus ruder-двигательный центр спинного мозга;
б) ядро глазодвигательного нерва-nucleus n. oculomotorius.
в) парасимпатические ядра Якубовича.
Каудальнее располагаются:
г) ядро блокового нерва-nucleus n. trochlealis;
д) часть ядра пятой пары нервов.
Через весь чепец из продолговатого мозга в промежуточный проходит сетчатое образование, образующее проводящие пути из спинного мозга и мозжечка в четверохолмие, в зрительные бугры, и из них в спинной мозг.
3.ножки большого мозга-pedunculi cerebri- выступают на базальной поверхности мозга в виде двух толстых валиков между зрительными трактами и мозговым мостом. Они разделены межножковой бороздой. Из ножек выход 3 пара черепно-мозговых нервов - глазодвигательный нерв. Ножки образованы эфферентными проводящими путями, которые соединяют кору плаща с ромбовидным и спинным мозгом. Поэтому ножки сильнее развиты у тех животных, у которых кора полушарий более мощная. У человека они развиты наиболее сильно.
Развитие среднего мозга
Средний мозг у низших животных, как и у эмбрионов высших животных достигает очень значительных размеров. Серое мозговое вещество эмбриональных боковых пластинок среднего мозгового пузыря, разрастаясь, формирует свод среднего мозга, из которого у низших животных происходит двухолмие, или зрительные доли. Первоначально двухолмие было высшим координирующим центром, поскольку в нем оканчиваются не только зрительные нервы, но и афферентные проводящие пути из полосатого тела, ромбовидного и спинного мозга. Но уже у рептилий часть волокон, а у млекопитающих почти все волокна зрительного нерва перемещаются через зрительные бугры промежуточного мозга в кору вторичного плаща. Поэтому зрительные доли отстают в росте, а вторичный плащ и соответственно зрительные бугры разрастаются все сильнее и сильнее.
У наземных животных в своде среднего мозга закладывается равновесно-слуховые центры, первоначально в виде микроскопических образований, а позднее и макроскопических. Лишь у млекопитающих вместо двухолмия появляется четверохолмие. У животных с хорошим слухом преобладают задние - слуховые холмы.
Из эмбриональной основной пластинки в вентральной стенке среднего мозгового пузыря формируется чепец. В результате образования свода и чепца полость среднего мозга превращается в мозговой водопровод. Чепец состоит из ядер 3 и 4 пар черепно-мозговых нервов и специальных двигательных ядер чепца. К последним относятся красное ядро, связывающее мозжечок со спинным мозгом, и межножковое ядро, соединяющееся через ганглий уздечки с обонятельным мозгом.
У млекопитающих в связи с развитием вторичного плаща чепец вентрально порывается слоем белого мозгового вещества из проводящих путей, идущих из коры в ромбовидный и спинной мозг. Эти проводящие пути формируют ножки большого мозга, мощность которых соответствует степени развития коры полушарий большого мозга.
Ромбовидный мозг-rhombencephalon
Подразделяют на продолговатый и задний мозг. Задний мозг состоит из мозжечка и мозгового моста. Между мозжечком и продолговатым мостом находится четвертый мозговой желудочек.
А) Продолговатый мозг-medulla oblongata
Каудально продолжается без заметной границы в спинной мозг. На базальной поверхности его хорошо видна вентральная срединная борозда –fissura mediana ventralis. По обе стороны от нее проходят боковые борозды-sulcus paramedianus- каудально они вливаются в срединную вентральную борозду. Между этими тремя бороздами выступают два узких валика- пирамиды-pyramis- в них проходят пирамидные проводящие пучки от коры большого мозга в спинной мозг. Так как проводящие пучки продолжаются в боковые канатики и спинного мозга и при этом перекрещиваются справа налево и наоборот, то образуется перекрест пирамид-decussatio pyramidum. Латерально от пирамиды позади моста выходит 4 пара черепно-мозговых нервов - отводящий нерв. Близ перекреста пирамид и латерально от него отходит 12 пара - подъязычный нерв, латерально же от подъязычного нерва один за другим еще два нерва: 10 пара- блуждающий нерв, более назально- 9 пара- языкоглоточный нерв.
Серое мозговое вещество продолговатого мозга сгруппировано:
а) в отдельные чувствительные и двигательные ядра, из которых выходят 5, 6,7,9,10 и 12 пары черепно-мозговых нервов;
б) в промежуточные ядра пучков Голля и Бурдаха;
в) в ядра 8 пары и связанные с ними назальные и каудальные оливы-7.
Среди ядер залегает сетчатое образование-formatio reticularis- из переплетающихся нервных волокон и нервных клеток между ними, которое из продолговатого мозга переходит в чепец среднего мозга и в промежуточный мозг. Оно выполняет, прежде всего, ассоциационную и координационную функцию между различными ядрами ромбовидного и среднего мозга и является центром дыхания и сердечнососудистой системы.
Белое мозговое вещество продолговатого мозга состоит из большого количества пучков проводящих путей, идущих из спинного мозга в различные отделы головного мозга и обратно.
Б) Мозжечок-cerebellum
Имеет почти шаровидную форму, двумя бороздами он разделяется на среднюю часть- червячок и две боковые доли. Серое мозговое вещество образует кору мозжечка-cortex cerebelli- и отдельные ядра, залегающие в центрально расположено белом веществе мозжечка. Функция- координация движений.
Поверхность коры червячка-vermis- изрезана поперечными бороздами и щелями. Двумя основными щелями червячок разграничен на переднюю, среднюю и заднюю доли-lobus rostralis, caudalis et medialis. Каждая из них связана с ножками мозжечка - передними, средними и задними, состоящими из проводящих путей. Передний и задний концы червяка загнуты вентрально и навстречу друг другу, между ними остается небольшая щель- верхушка шатра-fastigium. Белое мозговое вещество червячка-7- на сагиттальном разрезе напоминает ветку туи, поэтому оно получило название- древо жизни-arbor vitae. В белом мозговом веществе червячка находится шатровое ядро-nucleus fastigii- которое является подкорковым центром равновесного анализатора.
С продолговатым мозгом червячок соединяется через каудальный мозговой парус-velum medullare caudale- а с четверохолмием через назальный мозговой парус-velum medullare nasale.
Полушария мозжечка-hemisphaerum cerebelli- как и червячок состоят из многочисленных долек, из которых одна - клочок-flocculus- в виде небольшого придатка червячка лежит сзади от боковой ножки. В белом мозговом веществе полушарий мозжечка находятся зубчатые ядра-nucleus dentatus- служащие передаточными центрами двигательных импульсов.
Мозжечок соединяется с продолговатым мозгом. С мозговым мостом - боковыми ножками и со средним мозгом - назальными ножками.
Каудальные ножки мозжечка-pedunculus cerebellaris medullum oblongatum- в виде двух валиков выступают на дорсальной поверхности продолговатого мозга. В них проходят проводящие пути:
а) из спинного мозга- пучки Флексига, Голля и Бурдаха с их ядрами;
б) из ядер слухового нерва;
в) из каудальных олив;
г) из ядер 5,10 и 12 пар черепно-мозговых нервов.
Назальные ножки мозжечка-brachium conjum ctivum cerebelli- идут под задними холмами четверохолмия в ножки большого мозга. Назальные ножки лежат на дорсальной поверхности продолговатого мозга. В них проходят проводящие пути:
а) из спинного мозга в червячок- пучок Говерса;
б) из зубчатых ядер полушарий мозжечка в красное ядро;
в) в ядра зрительных бугров.
Собственные проводящие пути мозжечка представлены волокнами между корой мозжечка и его ядрами и ассоциационными волокнами, соединяющими друг с другом отдельные извилины в сагиттальной плоскости.
В) Мозговой мост-pons cerebri
Лежит на переднем конце продолговатого мозга, на границе его со средним мозгом, в виде поперечного валика, который своими концами загибается дорсально к мозжечку, образуя боковые ножки мозжечка-brachium pontis. Мост и ножки состоят из проводящих путей, соединяющих ядра моста-nucleu pontis- с ядрами мозжечка. В ядрах моста заканчиваются проводящие пути из коры большого мозга и начинаются проводящие пути в полушария мозжечка. Через боковые отделы моста выходит 5 пара - тройничный нерв - самый массивный нерв из всех черепно-мозговых нервов. Он имеет два корня: вентрально - назальный двигательный и дорсо - каудальный чувствительный. На последнем находится большой полулунный ганглий. Каудально от моста, также в поперечном направлении, лежит трапецоидное тело-corpus trapezoideum- в виде узкого и низкого валика. Оно сформировано проводящими путями, идущими от ядер слухового нерва. Через боковые части трапецоидного тела выходят 8 пара- равновесно-слуховой нерв и 7 пара -лицевой нерв.
Г) Четвертый мозговой желудочек-ventriculus quartus
Помещается между мозжечком и продолговатым мозгом. Сводом ему служат червячок и мозговые паруса, а дном - продолговатый мозг.
Дно четвертого желудочка называется ромбовидной ямкой-fossa rhomboidea. Срединной и двумя боковыми бороздами на дне ямки выделяется парное срединное возвышение-eminentia medialis- на котором против латеральной ножки мозжечок выступает лицевой холмик-colliculi facialis. В области лицевого холмика залегают ядра отводящего и лицевого нервов. На каудальном конце срединного возвышения находится поле подъязычного нерва-area hypoglossi- с одноименным ядром.
Латерально от подъязычного нерва выступают ядра 9 и 10 пар нервов. Они формируют серое крыло-ala cinerea. Область заднего конца серых крыльев известна под названием писчего пера-calamus scriptorius.
Непосредственно позади боковых ножек мозжечка и медиально от них выступают в виде небольших возвышений вестибулярные поля-area vestibularis. Они содержат вестибулярные и улитковые ядра 8 пары нервов. Улитковые ядра лежат латеральнее.
Развитие ромбовидного мозга
Продолговатый мозг развивается путем разрастания боковых и основных пластинок эмбриональной мозговой трубки. Вследствие раздвигания боковых пластинок формирующимся мозжечком двигательные центры продолговатого мозга оказались оттесненными медиально, а чувствительные латерально.
Эти же обусловливается и медиальное расположение на продолговатом мозге двигательных нервов и латеральное - чувствительных, а также особенности строения дна четвертого желудочка мозга, в толще которого чувствительные ядра лежат латерально от двигательных.
Строение продолговатого мозга в целом у животных макроскопически довольно однообразно, что объясняется и однообразной его функцией - центры жаберного аппарата и его производных, кишечника, центры органов осязания, суставно- мышечного чувства, слуха и равновесия. Ввиду того, что через продолговатый мозг проходят проводящие пути, соединяющие головной мозг со спинным, его серое мозговое вещество разбивается на отдельные ядра в отличие от компактного серого мозгового вещества спинного мозга.
В продолговатом мозге заслуживает особого внимания моторное ядро чепца или сетчатое образование, которое впервые возникает у рыб и является древнейшим ассоциационным и двигательным центром. У наземных животных за счет моторного ядра чепца формируются назальным и каудальные оливы как ассоциационные центры. Назальные оливы появляются только у наземных животных, начиная с амфибий. Они служат промежуточным центром на пути от улиткового нерва к зрительным буграм. Пучок этих волокон образует трапецоидное тело. Каудальные оливы формируются еще позднее у птиц и млекопитающих. Они связаны с ядрами дорсальных канатиков, со зрительными буграми, с мозжечком и спинным мозгом. По своим связям каудальные оливы имеют самое ближайшее отношение к функции сохранения равновесия.
Мозжечок развивается в связи с выполняемой им функцией сохранения равновесия тела и поддержания мышечного тонуса. Поэтому он выражен наиболее сильно у животных, быстро плавающих, бегающих, прыгающих, летающих, и слабее у животных, передвигающихся медленно. В примитивном виде мозжечок представляет непарную пластинку и бело или серого мозгового вещества. Мозжечок закладывается в средней части эмбриональной пластинки покрышки ромбовидного мозга, а из передней и задней частей последней формируются передний и задний мозговые паруса. Пластинка мозжечка, разрастаясь спереди назад, изгибается дугообразно в дорсальном направлении. Благодаря появлению продольных борозд обособляется средняя часть пластинки- тело мозжечка- и парные боковые части- ушки мозжечка. Тело мозжечка и ушки относятся к palaeocerebellum. Тело мозжечка у наземных животных разделяется поперечными бороздами на переднюю, среднюю и заднюю доли, на которых возникают добавочные поперечные борозды. Передняя доля связана с мышцами головы, а средняя и задняя- с мышцами туловища и конечностей.
У млекопитающих преобладает средняя доля. Продольные борозды на ней отделяют среднюю непарную часть- червячок - от парных боковых долей, или полушарий мозжечка. Полушария мозжечка относятся к neocerebellum. В червячке находятся центры координированных, синхронных движений туловища и конечностей. Полушария мозжечка сильнее всего выражены у высших млекопитающих, обладающих в большей или меньшей степени способностью обособленных движений конечностей. Совершенствование этой способности, в свою очередь зависит от мощности коры большого мозга, как высшего центра нервной деятельности, и от возникновения связей мозжечка с корой большого мозга через боковые его ножки и мост.
Полушария мозжечка и мост имеются только у млекопитающих и достигают максимального своего развития у приматов. Однородной функцией мозжечка у животных объясняется довольно однообразное гистологическое строение его коры, в которой различают поверхностный молекулярный слой, глубокий зернистый и средний слой из крупных клеток Пуркине.
Ушки примитивного мозжечка у водных животных имеют отношение к органам равновесия, то есть органам боковой линии, и к мускулатуре хвоста. С редукцией этих органов у наземных животных уменьшаются и ушки. У млекопитающих от них сохраняются клочки, соединяющиеся с задней долей червячка.
Онтогенез червячка также определен его функцией, что особенно хорошо видно на мозге человека. У новорожденного ребенка он еще очень мал и составляет лишь 1/16-1/18 веса большого мозга. Но эти соотношения изменяются по мере роста ребенка и увеличения его подвижности, у взрослого отношения уже иные – 1/8-1/9 веса мозга.
Список используемой литературы
1) А.И. Акаевский «Анатомия Домашних Животных»
2) И.В. Хрусталева «Анатомия домашних животных»
3) Л.В. Антипова, В.С. Слободяник, С.М.Сулейманов «Анатомия и гистология сельскохозяйственных животных»
4) А.Ф. Римов «Анатомия домашних животных»
5) А.Ф. Кузнецова «Справочник по ветеринарной медицине»
Похожие работы
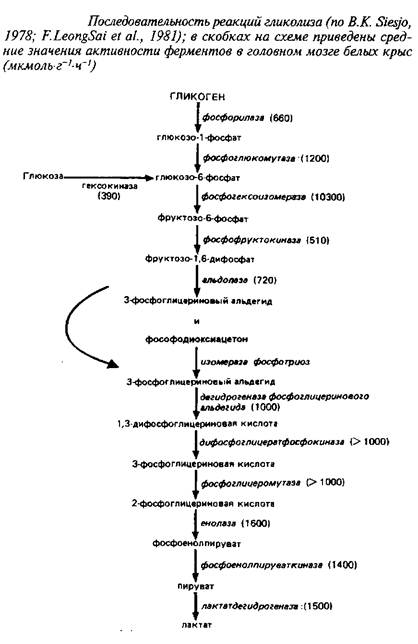
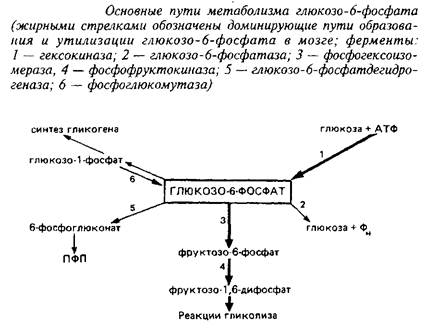
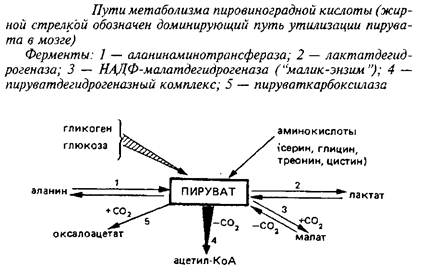
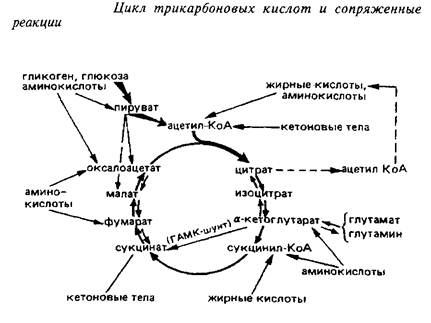
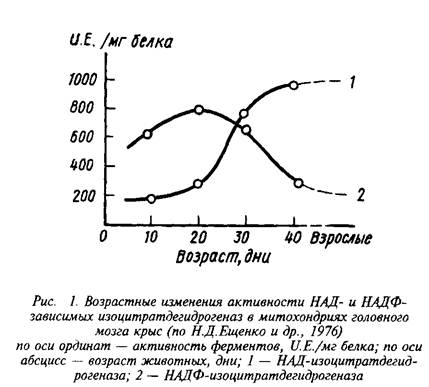
... 4,8±0,6 Аспарагиновая кислота + аспарагин 4,8–5,8 0,4–0,7 6,0±1,5 6,3±0,8 ГАМ К 1,9–2,4 Следы 3,1 + 1,1 4,1±0,4 Из других путей метаболизма аминокислот, которые играют относительно небольшую роль в энергетическом метаболизме головного мозга, можно упомянуть превращение аспартата и аспарагина в оксалоацетат, а также аланина и серина – в пируват. Очень невелико и значение аминокислот как ...
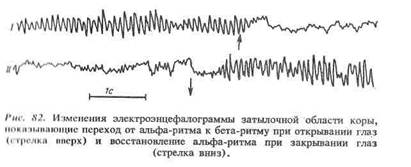
... принимающие участие в образовании временных нервных связей. Установлено, что в формировании рефлекторной дуги условных рефлексов принимают участие нейроны ретикулярной формации, подкорковых ядер, лимбической системы. Следовательно, замыкательный процесс может осуществляться на разных уровнях головного мозга. Однако кора больших полушарий, тесно взаимодействуя с нижележащими отделами мозга, играет ...
... эффект ?-сон индуцирующего пептида при гипокинетическом стрессе // Укр.биохим.-1991.-63.-№1.-С.34-37. 118.Механизмы развития стресса // Сб.статей.- Кишинев: Штиинца.- 1987.-222с. 119.Митюшина Н.В. Влияние энкефалинов на активность ферментов обмена регуляторных пептидов в головном мозге и периферических тканях крыс // дис. на соиск. степени.канд.биол.наук.- Пенза.-1999 120.Наркевич В.Б. ...
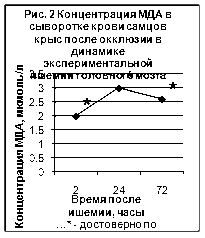

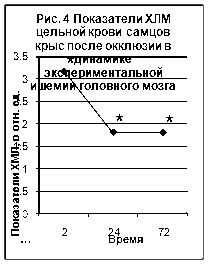
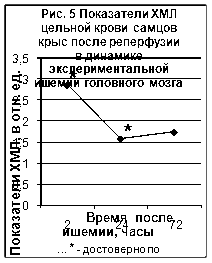
... ТБК-активных продуктов – малонового диальдегида (МДА), показателям люминолзависимой хемилюминесценции. 3.2. Исследование активности хемилюминесценции цельной крови и концентрации МДА в сыворотке крови при экспериментальной ишемии головного мозга Центральное место в изучении ишемического инсульта занимают экспериментальные модели на животных [65]. Именно по результатам эксперимента ...
0 комментариев