Навигация
Структурная организация нервно-мышечной системы
63491
знак
3
таблицы
14
изображений
1.2 Структурная организация нервно-мышечной системы
Нервно-мышечный аппарат представляет собой функционально связанный комплекс мышц и образований нервной системы: мотонейронов и их аксонов[41]. Мотонейрон, его аксон и группа мышечных волокон, которые иннервируется данным аксоном являются структурным элементом нервно-мышечного аппарата - двигательной единицей и используется как основное понятие в миологии.
Мышечные волокна, относящиеся к одной двигательной единице, действуют по принципу «всё или ничего», когда вследствие импульса, превышающего определённый порог чувствительности, провоцирует сокращение всех двигательных волокон входящих в состав двигательной единицы.
Функционально двигательные единицы различаются между собой, что обусловлено морфологическим и гистохимическим полиморфизмом. Все мышечные элементы одной двигательной единицы одинаково окрашиваются гистохимическими реактивами. По гистохимической реакции выделяют два типа волокон: I и II. На основании этого определены два основных типа двигательных единиц.
Двигательные единицы I типа (медленные двигательные единицы) включают в свою структуру «медленный» мотонейрон, «медленный» аксон, «медленные» мышечные волокна. Медленные мотонейроны — малые по величине (альфа-малые мотонейроны) — имеют высокую возбудимость, низкую частоту генерации импульса, высокую выносливость, менее утомляемы. Обмен в них преимущественно аэробный. Аксон этих клеток тонкий, слабо миелинизирован, и скорость проведения возбуждения по нему невелика, но в отличие от толстых аксонов он более возбудим при непрямой электрической стимуляции. Медленные мышечные волокна, состоят из не большого количества миофибрилл и поэтому развивают меньшее усилие, однако они относительно более выносливы и могут длительное время давать стабильную импульсацию. Такие мышечные клетки имеют богатую сеть капилляров, что обеспечивает высокую степень окисления. В целом, медленные двигательные единицы обеспечивают длительное (тоническое) напряжение мышцы.
Двигательные единицы II типа (быстрые двигательные единицы) состоят из "быстрых" элементов. Быстрые мотонейроны - более крупные по величине, но менее возбудимы, могут давать высокую частоту импульсации. В клетках преобладает анаэробный обмен. Они имеют толстый аксон с хорошо развитым слоем миелина, что обеспечивает высокую скорость проведения импульса. Быстрые мышечные волокна более толстые, обладают мощной лактацидной анаэробной системой энергообеспечения. Они способны развивать достаточно большое усилие за короткий период времени, но быстро утомляются. Быстрые двигательные единицы обеспечивают мощное быстрое (фазическое) напряжение.
Среди быстрых двигательных единиц выделяют два подтипа: 2А — медленно утомляемый и 2В — быстро утомляемый. Эти подтипы двигательных единиц различаются порогом возбуждения, частотным диапазоном импульсации, а также особенностями обмена.
Площадь и количество двигательных единиц зависят от размеров мышцы и выполняемой функции. Один мышечный пучок может содержать в себе волокна от разных двигательных единиц.
Мышечные волокна I и II типа располагаются мозаично по всему поперечнику мышцы, рядом может лежать не более 2-3 мышечных волокон одного гистохимического типа. Таким образом, в области одного пучка представлены мышечные волокна разных двигательных единиц.
Мак-Комас А.Дж. (2001) указывает на возможность определения количества двигательных единиц в мышце при помощи регистрации М-ответа.
При достижении мотонейрона волна антидромного возбуждения вызывает активность в самом чувствительном его месте — аксональном холмике. Благодаря особенностям строения мембраны аксональный холмик имеет минимальный порог возбуждения, но он также вариабелен для данного пула мотонейронов. К антидромному возбуждению более склонны крупные (фазические) мотонейроны. Теоретически считалось, что только они способны к антидромному ответу, При проведении опытов с блокированием быстрых аксонов было показано, что антидромный ответ возникает и в малых мотонейронах с медленно проводящими аксонами.
Важным фактом является то, что все мышечные волокна, составляющие двигательную единицу, имеют одинаковый гистохимический тип. Вследствие этого, изучение элементарных структурно-функциональных систем периферического нейромоторного аппарата даёт возможность дать оценку состоянию мышцы, как функционального целого.
В связи с тем что исследование проводилось на верхних конечностях ниже будет представлен нервный аппарат кисти плеча и предплечья (рис. 1).
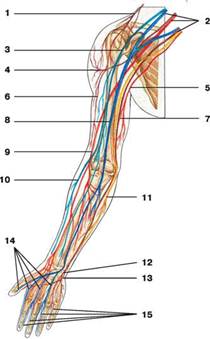
Рис. 1. Схема нервов верхней конечности: 1 - надключичный нерв; 2 - плечевое сплетение; 3 - мышечно-кожный нерв; 4 - подкрыльцовый нерв; 5 - медиальный кожный нерв плеча; 6 - латеральный кожный нерв плеча; 7 - локтевой нерв; 8 - срединный нерв; 9 - лучевой нерв; 10 - латеральный кожный нерв предплечья; 11 - медиальный кожный нерв предплечья; 12 - поверхностная ветвь локтевого нерва; 13 - глубокая ветвь локтевого нерва; 14 общие ладонные пальцевые нервы; 15 - собственные ладонные пальцевые нервы
Количество пар спинно-мозговых нервов и их локализация соответствуют сегментам спинного мозга: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых, 1 копчиковая пара. Все они отходят от спинного мозга задним чувствительным и передним двигательным корешками. Корешки объединяются в один ствол и выходят из позвоночного канала через межпозвоночное отверстие. В области межпозвоночного отверстия находятся спинальные узлы (ganglion spinale), представляющие собой скопление чувствительных клеток и являющиеся частью задних корешков. От клеток спинального узла начинаются чувствительные волокна, а от клеток переднего рога -двигательные. Объединившись, нервы становятся смешанными. После выхода из межпозвоночного отверстия спинно-мозговые нервы разделяются на задние и передние смешанные ветви. Задние направляются к мышцам и коже задних отделов туловища, а передние иннервируют мышцы переднего отдела туловища и конечности. Объединяясь друг с другом в других отделах, нервы образуют шейное, плечевое, поясничное и крестцовое сплетения.
Плечевое сплетение (plexus brachialis) образуется ветвями четырех нижних шейных нервов и передней ветвью 1-го грудного нерва. Ветви сплетения выходят на шею между передней и средней лестничными мышцами и направляются в подмышечную область. Сплетение состоит из надключичного отдела, образованного короткими ветвями, направляющимися к плечевому поясу, груди и спине, и подключичного отдела, в состав которого входят длинные ветви, иннервирующие кожу и мышцы свободной части верхней конечности.
Подключичный отдел находится в подмышечной области и состоит из трех пучков: медиального, латерального и заднего. Стволы этих пучков иннервируют подмышечную артерию и являются началом длинных ветвей.
Медиальный ствол включает:
- медиальный кожный нерв плеча (n. cutaneus brachii medialis) направляющийся к коже медиальной поверхности плеча;
- медиальный кожный нерв предплечья (n. cutaneus antebrachii medialis), иннервирующий кожу медиальной поверхности предплечья;
- локтевой нерв (n. ulnaris), который является смешанным. Его чувствительные волокна направляются к коже медиальных отделов кисти. На ладонной поверхности они иннервируют кожу V пальца и локтевой стороны IV пальца, на тыльной поверхности - кожу IV и V пальцев и локтевой стороны III пальца. Двигательные волокна на предплечье направляются к локтевому сгибателю запястья и медиальному отделу глубокого сгибателя пальцев. На кисти они иннервируют мышцу, приводящую большой палец кисти, мышцы возвышения мизинца, а также 3–4-ю червеобразные мышцы.
Латеральный ствол включает:
- срединный нерв (n. medianus), который также относится к смешанным. Он выходит из латерального и медиального стволов. Чувствительные волокна направляются к коже латерального отдела ладонной поверхности и коже I, II и III пальцев, а также к лучевой стороне IV пальца и отчасти к тыльной поверхности этих пальцев. Двигательные волокна на предплечье иннервируют сгибатели предплечья, за исключением локтевого сгибателя запястья и глубокого сгибателя пальцев, а также направляются к квадратному и круглому пронаторам. На кисти двигательная часть иннервирует мышцы возвышения большого пальца;
- мышечно-кожный нерв (n. musculocutaneus), являющийся смешанным. Его ветви направляются к сгибателям передней поверхности плеча;
- латеральный кожный нерв предплечья (n. cutaneus anterbrachii lateralis) который является концевой ветвью предыдущего нерва и иннервирует область предплечья.
Задний ствол включает:
- лучевой нерв (n. radialis) , являющийся смешанным. Чувствительные волокна направляются к коже латеральных отделов тыльной поверхности кисти и I, II пальцев, а также лучевой стороны III пальца. Двигательные волокна иннервируют разгибатели плеча и предплечья;
- задний кожный нерв плеча (n. cutaneus brachii posterior), который является чувствительной ветвью лучевого нерва и направляется к коже задней поверхности плеча;
- задний кожный нерв предплечья (n. cutaneus anterbrachii posterior), также являющийся чувствительной ветвью лучевого нерва и иннервирующий кожу задней поверхности предплечья.
1.3 Биомеханическая характеристика силовых качествЛюбые движения человека-это результат согласованной деятельности ЦНС и периферических отделов двигательного аппарата, в частности скелетно-мышечной системы. Без проявления мышечной силы никакие физические упражнения выполнять невозможно.
Чрезвычайно важной особенностью мышечной силы, проявляемой в динамическом режиме, является то, что ее проявление может быть мгновенным. Наибольшая величина мгновенной силы будет характеризовать максимальную динамическую силу. Однако, как известно, проявление мышечной силы при выполнении любого движения всегда протекает во времени. В этом случае конечный эффект постоянного проявления силы во времени определяется импульсом силы – F*t.
Исследуя механизмы динамики мышечного сокращения при преодолении на инерционном динамографе, Н.Н. Гончаров при обработке полученных данных ввел понятие средняя сила, которая, по его расчетам, равна 50% максимальной динамической силы. Средняя динамическая сила представляет собой условную величину, удобную для оценки эффекта действия силы по полной амплитуде движения и максимально волевом усилии. При статическом режиме работы мышц сила замеряется как абсолютная и относительная статическая сила.
Говоря о механических свойствах активной мышцы, необходимо помнить о наличии тех многообразий, сочетание которых характеризует механическая динамика мышечного сокращения. В настоящее время с уверенностью можно говорить о четырех зависимостях, каждая из которых дает лишь частичное представление об активной мышце. Наиболее подробно и всесторонне данная проблема изучена В.М. Зациорским, анализ работ которого позволяет в самом кратком изложении представить ее следующим образом:
а) Кривая длин напряжений системы последовательных эластических компонентов. Данная кривая не зависит прямо от контрактильного механизма и может быть выражена уравнением:
P=f (l-1)
Где S (см.) – растяжение;
P – нормализованное (т.е. приведенное к Р =1) напряжение;
f – константы;
б) Кривая длин напряжений активной мышцы. Можно предположить, что данная кривая отражает свойства контрактильного протеина внутримышечных фибрилл и может быть выражена предложенным А. Хиллом (1922) уравнением:
cos
Где P – максимальное напряжение (при r=0);
R – максимальное укорочение (P=0);
P и r – соответствующие мгновенные значения напряжения и укорочения;
в) кривая сила-скорость. Можно полагать, что данная зависимость отражает те же свойства, что и предыдущие. Рассматриваемая кривая может быть выражена так называемым основным уравнением мышечного сокращения, предложенным А. Хиллом:
(P+a) (v+b)=(P +a)b
где P – максимальное напряжение при данной длине мышцы;v – скорость;
P – напряжение;
а и b – константы, которые можно получить как из кривой сила- скорость, так и в результате биометрических измерений;
г) кривая активного состояния является результатом механизма, в котором контрактильный компонент включается и выключается в ответ на изменение потенциалов в клетках мембран. Мышечная сила человека при прочих равных условиях пропорциональна площади физиологического поперечника мышцы. Это еще отметил немецкий физиолог Е. Вебер (1846). Известно, что 1 см. мышцы поднимает от 6 – 10 кг. безотносительно к тому, тренирован или не тренирован ее обладатель.
Зависимость мышечной силы от физиологического поперечника мышцы признают все специалисты в области анатомии и физиологии. В то же время в работах по физиологии отмечается, что важнейшим фактором проявления силы является не периферическое изменение, а регуляция работы мышц со стороны нервных центров.
Важным моментом для понимания механизма мышечного напряжения является то, что по мере роста проявления мышечной силы частота колебания потенциала одной двигательной единицы может возрасти с 5 – 6 до 35 – 40 раз в секунду. Однако поскольку предельная частота колебаний намного меньше частоты, при которой мышца начинает трансформировать ритм поступающих в нее импульсов, можно полностью согласиться с мнением В.М. Зациорского о том, что деятельность мышцы не связана с трансформацией ритма, как это предполагали ранее. Исследования показали, что частота импульсов линейно пропорциональна развиваемой кинетической энергии. Что же касается амплитуды токов действия одного миоцита, то она, как правило, не изменяется.
Только при различии пороговых значений амплитуда токов действия может увеличиться из-за неодновременного включения в работу отдельных волокон. Что касается электроактивности всей напрягаемой мышцы, то она также возрастает по мере роста величины ее напряжения, но до определенного предела.
При мышечных напряжениях, когда они не доходят до предельных величин, регуляция мышечной силы происходит за счет изменения различного количества двигательных единиц.
В основе регуляции двигательных единиц в этом случае лежит механизм асинхронности. По данным русского ученого Р.С. Персон, асинхронизация определяется проприоцептивным влиянием, которое накладывается на синхронную импульсацию центральных и моторных структур. При этом степень напряжения не регулируется потенциалом отдельных импульсов, поскольку первое волокно является проводником импульсов, характеризующихся постоянной величиной потенциала. В результате создаются условия для получения большей надежности при значительной пропускной способности накала и принципиальной простоте, что позволяет обеспечивать передачу возбуждения в широком диапазоне при относительно небольшом применении частоты импульсации.
Величина проявления силы при выполнении физических упражнений во многом зависит от формирования условных рефлексов, которые обеспечивают необходимую концентрацию процессов возбуждения и торможения и вовлечение в однократное максимальное сокращение наибольшего числа двигательных единиц (Д.Е.) при оптимальном возбуждении мышцах-антагонистах.
В напряжении мышцы, как полагает целый ряд исследователей, участвуют не все двигательные единицы. При этом чем сильнее возбуждение, тем большее число Д.Е. принимает участие в сокращении. Наибольшее проявление силы может быть достигнуто (если прочие условия равны) при одновременном сокращении максимально возможного количества всех двигательных единиц в мышце.
До настоящего времени неясным в механизмах регуляции мышечного напряжения является деятельность центрально-нервных механизмов. Исследования, выполненные в последние годы, дают возможность предполагать, что имеется по крайней мере три ведущих механизма. Один из них, в основе которого лежит рефлекс на растяжение (миотатический рефлекс), связан с регуляцией напряжения при сохранении положения тела. Изменение позы тела меняет и растяжение мышечных веретен, тем самым способствуя возбуждения их рецепторного аппарата, что в свою очередь рефлекторно вызывает изменение мышечного напряжения растянутых мышц.
При выполнении движений, не требующих проявления максимальной мышечной силы, для дозирования мышечного напряжения используется другой механизм. В этом случае высшие нервные центры определяют в основном необходимые величины пространственных, временных и скоростных параметров движения. Что касается нужных комбинаций мышечных напряжений, то он осуществляется более низко расположенными нервными отделами. Известно, что эффекторная импульсация поступает сначала не в мышечные волокна, а в мускульный аппарат мышечных веретен, что приводит к изменению натяжений в них и соответствующему возбуждению их рецепторного аппарата. Далее регуляция осуществляется по схеме миостатического рефлекса.
При выполнении движений, требующих предельных величин проявления мышечной силы, эффекторная импульсация поступает от соответствующих отделов головного мозга через мотонейроны прямо в Д.Е.
В экспериментальных исследованиях было показано, что предварительно растянутая до определенной оптимальной степени мышца сокращается сильнее и быстрее.
Следовательно, использование эластичных свойств мышцы также будет способствовать проявлению большой силы. В динамической анатомии такую работу мышц принято называть баллистической. И.М. Сеченов писал: «Груз действует на мышцы одновременно в двух противоположных направлениях – растягивает ее как всякое упругое тело, и усиливает в то же время развитие в ней сократительных осей».
Таким образом, с одной стороны, физиологи установили, что сила человека пропорциональна массе мышц, с другой стороны, биологи доказали, что с увеличением массы у представителей одного и того же класса животных, например млекопитающих, уменьшается относительная сила, то есть отношение абсолютной величины максимальной силы к весу тела.
0 комментариев