Навигация
3.3 Мутагенна дія
Під впливом зовнішнього й внутрішнього опромінення радіонуклідами, викинутими при аварії, у рослинах реєстрували мутації як у соматичних, так і в статевих клітинах.
Виникнення мутацій зв'язується, як правило, із прямим фізичним руйнуванням або функціональною інактивацією під впливом продуктів радіолізу ділянок хромосом і їхніх унікальних структур. Про ушкодження унікальних структур може свідчити збільшення розривів у молекулах ДНК. Використовуючи прийом додаткового гамма-опромінення рослин, що вже одержали певну дозу радіації, за допомогою методу електрофорезу однониткової ДНК і лужного розплітання, виявили при обстежені в 1987р. популяціях трав'янистих рослин, які ростуть на околицях ЧАЕС, зокрема, у радіочуттєвої бобової рослини горщика мишачого (Vicia cracca L.) підвищений вихід однониткових ДНК [20, 41-46].
Формування генетичних змін і їхня фіксація в наступних поколіннях багато в чому залежать від функціонування систем репарації ДНК. При цьому в рослин дані системи грають особливо важливу роль у пилку, що обумовлено її особливою функцією й гаплоїдністю ядер. Ефективність функціонування систем репарації ДНК повинна забезпечити стабільність генома, що може легко ушкоджуватися внаслідок слабкої захищеності дозрілого пилкового зерна. Тому ослаблення функціонування систем репарації є свідченням посилення генетичної дії випромінювання. При випромінюванні спонтанного відновлення ДНК і індукованого гострим гамма-опроміненням позапланового синтезу ДНК, що відбиває її репарацію, було встановлено, що формування пилка берези бородавчастої (Betula pendula L.) при високих концентраціях радіонуклідів у ґрунті приpзводить до придушення системи темнової репарації ДНК у зрілому пилку, що зберігається, [20, 73]. При збереженні в такого пилка здатності до запліднення можна чекати в рослин нагромадження генетичних ушкоджень.
Багатьма дослідниками відзначено збільшення аберацій хромосом в утворювальних тканинах рослин, які ростуть в зоні аварії ЧАЕС. Так, виявлено [16, 28-35] збільшення в кілька разів числа клітин з абераціями хромосом у кореневих мерисистемах проростків ослинника дворічного (Oenothera biennis L.), вирощених у лабораторних умовах з насіння рослин, що виростають при рівнях гамма-фоону в 1986р. від 0,05 до 60мР/ч. При цьому показана характерна прямо пропорційна залежність виходу мутацій від потужності дози й виявлені множинні аберації хромосом, які можуть бути наслідком дії щільно-іонізуючого альфа-випромінювання.
При проведенні досліджень [16, 38-41] на різних за рівнем забруднення радіоактивними випаданнями ділянках (від 120 до 800 МБк/м2),спостерігали лінійну або близьку до неї залежність між поглиненою протягом 26.04 - 5.06 (збір насіння) 1986р. сумарної бета- і гамма-дозою рослинами озимого посівного жита (Secale cereale L.) і виходом клітин з абераціями хромосом у кореневих меристемах проростків М 2-поколіннями (мал. 4.).
Цей же тест був використаний [13, 592] для оцінки впливу радіоактивного забруднення на насіння 7 видів рослин, зібраних у зоні, що прилягає до ЧАЕС. Потужність дози склала 2 - 3 мр/ч. При цьому в деяких видах, які відрізняються за радіочуттєвістю було виявлено від 1 до 8% аберацій хромосом, причому спостерігалося підвищення частоти аберацій зі збільшенням потужності дози. Але лінійної залежності не встановлено, що, очевидно, обумовлено нерівномірним розподілом радіонуклідів по поверхні ґрунту або впливом різних забруднювачів хімічної природи. Цікаво, що на другий рік після аварії частота аберацій хромосом у деяких видів зросла в 1,5 - 3 рази незважаючи на 2 - 3-кратне зниження радіаційного фону. Цей факт має винятково важливе значення з погляду генетичної небезпеки не тільки зовнішнього, але й внутрішнього опромінення за рахунок радіонуклідів, які попадають у рослини із ґрунту.
При вирощуванні високо-радіочуттєвих кінських бобів (Vicia faba L.) сорту Російські чорні [12, 149-152] в 30-кілометровій зоні при рівнях радіоактивного забруднення орного обрію ґрунту по 90Srблизько2·1012 Бк/км2 і по 137Cs близько 4·1012 Бк/км2 (поглинена доза для кореневої системи за 3 місяці вирощування рослин складала 0,15 - 0,4 Гр) реєстрували збільшення в 2 - 3 рази кількість клітин з абераціями хромосом.
Крім того, було встановлено [14, 46-48] збільшення кількості клітин з абераціями хромосом і хроматид у кореневій меристемі проростків скерди покрівельної (Crepis tectorum L.), вирощених з насіння рослин, зібраних на території зони ЧАЕС при умовах гамма-фону 5-10 мР/год у перший і 0,02 - 20 мР/год у другий роки після аварії. У проростках першого року максимальна частота клітин з абераціями досягала 10,2 - 15,3%, причому нерідко спостерігалися клітини із множинними абераціями. У другий рік на ділянках з максимальним рівнем фона було виявлено 1,4 - 2,2% клітин з абераціями хромосом. При мінімальних потужностях опромінення частота клітин з абераціями склала всього 0,3 - 0,5%; ці значення відповідають спонтанному рівню. Крім того, відзначали також появу рослин зі зміненим каріотипом, що свідчить про активні мікроеволюційні процеси в популяціях, які опромінюються хронічно.
Є дані [8, 3-9], що свідчать про підвищення кількості стерильного пилка в рослинах, що ростуть на територіях, забруднених радіонуклідами. Так, досліджуючи дві популяції фіалки ранкової (Viola matutina klok), що виростає на двох ділянках 30-кілометрової зони ЧАЕС, які різняться на 3 порядки по величинах поглинених доз, виявили в 1987 і 1988 р. збільшення в 1,5 - 2 рази кількості стерильних пилкових зерен. При рівні потужності дози на висоті 2 м 4 - 5 Гр/рік спостерігали часткову жіночу стерильність сосни (Pinus sylvestris L.), що виражалася в зниженні гамето-фитної виживаності сім’ябруньок, обпилених в 1986 р. і зменшенні ембріональної виживаності сім’ябруньок, обпилених в 1985 р.
Безумовно, стерильність — нездатність організму утворювати гамети або достатня їхня кількість, не приводить до генетичного ушкодження, а лише знижує число нащадків. Але при дії іонізуючої радіації стерильність, як правило, є наслідком генних або хромосомних мутацій, що обумовлюють порушення мейозу — основної ланки гаметогенезу.
Прямим свідченням генетичної дії іонізуючих випромінювань на пилок є безпосередня реєстрація в них мутацій. У рослин, вирощуваних на ґрунтах 30-кілометрової зони ЧАЕС із загальною бета-активністю 2,6·103- 6,3·104Бк/кг, в пилку ячменя (Hordeum vulgare L.) відзначали збільшення частоти waxy-змін в 2 - 3 рази [6, 147-156].
Таблиця 3
Кількість хлорофільних мутацій типу альбіна в житі (Secale cereale L.) і ячмені (Hardeum vulgare L.), %
| Культура | Контроль | 1986 р. | 1987 р. | 1988 р. | 1989 р. |
| Жито Київське 80 | 0,01 | 0,14 | 0,40 | 0,91 | 0,71 |
| Жито Харківське 03 | 0,02 | 0,80 | 0,99 | 1,20 | 1,14 |
| Ячмінь | 0,40 | 0,90 | 0,74 | 0,80 | 0,91 |
Примітка. Насіння отримані в 30-кілометровій зоні ЧАЕС.
У перші місяці після аварії при вирощуванні рослин жита (Secale cereale L.) і ячменя (Hardeum sativum jessen) в умовах закритого ґрунту на ґрунтах, привезених з 30-кілометрової зони, було виявлене збільшення частоти зустрічальності різних типів хлорофільних мутацій. Переважали нежиттєздатні мутації типу альбіна. У наступні роки насіння, зібране в зоні, висівали знову. У жита і ячменя, вирощуваних в 30-кілометровій зоні на ґрунті із сумарною гамма-активністю 1·105Бк/кг, реєстрували багаторазове перевищення спонтанного рівня виходу хлорофільних мутацій (табл. 3). При цьому було відзначено, що в кожному наступному поколінні мутації не елімінувалися, оскільки висіяні насіння піддавалися наступному пресингу радіонуклідів. Вихід морфологічних аномалій в 4 сортів озимої пшениці (Triticum vulgare L.) при їхньому вирощуванні протягом 3 років на ґрунті із сумарною активністю гамма-випромінювання 5,2·104- 1,8·105Бк/кг в 1986 - 1987 рр. перевищив 40 %, але в 1988 р. зменшився.
Спектр виявлених аномалій досить широкий: у перший рік після аварії найбільш частим (до 49 %) було утворення в колоссях стерильних зон — череззерниця; в 1987 р. кількість аномалій цього типу досягалася 30 %, а в наступному поколінні знижувалося до 1,9 %. Часто зустрічалися колосся з додатковими колосками ("грижі") і вкорочені колосся. До розповсюджених змін структури колосся варто також віднести аномалії типу скверхед.
Зустрічалися також рослини з підвищеною остистістю, нерівномірною остистістю, колосся "ялинка", зміни фарбування стебла й інші. У різних сортів вихід морфологічних аномалій проявляється по-різному, що може бути зв'язано, з одного боку, з різною їх радіо чуттєвістю, а з іншого боку — з неоднаковою здатністю до нагромадження радіонуклідів.
Безумовно, відмічувані морфологічні аномалії, відхилення від норми, як правило, мають місце лише в поколіннях організмів, що опромінюються, і зазвичай не закріплюються в потомстві. Проте їхній стійкий прояв в усі наступні роки, аж до 1994 р., дозволяє припускати, що більшість з цих змін носять генетичний характер і свідчать про те, що під впливом іонізуючих випромінювань радіонуклідів, викинутих при аварії на ЧАЕС, у популяціях рослин іде мутаційний процес.
Похожие работы
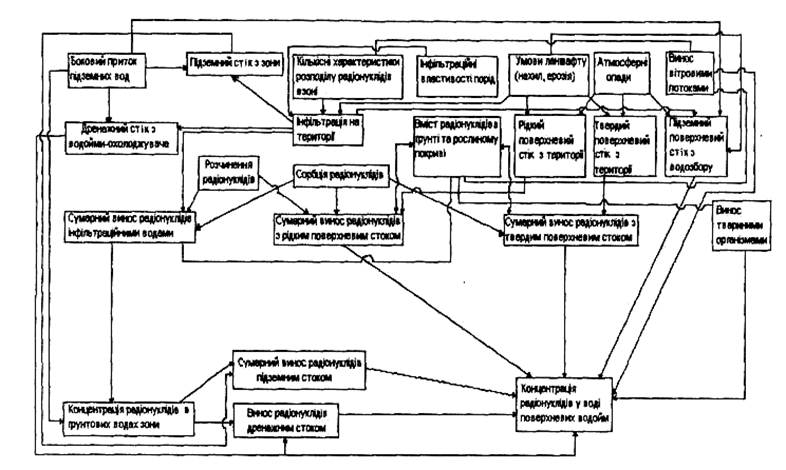
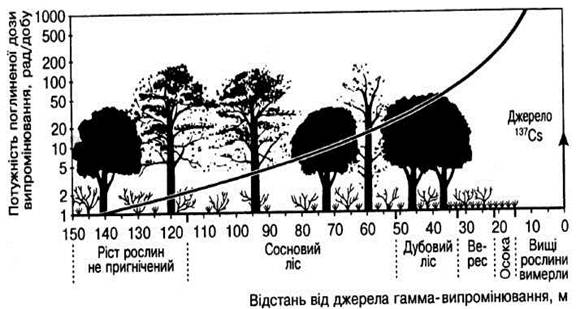
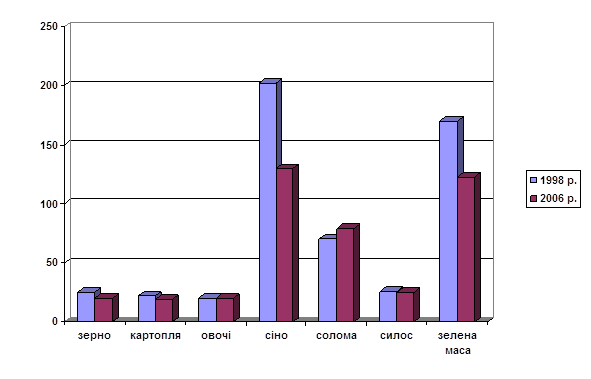
... селищ і сіл за шляхом вітрового перенесення аерозолів і попелу. Багато важливих проблем радіоекології горілого лісу ще потребують детального дослідження. РОЗДІЛ 5 АНАЛІЗ РАДІОАКТИВНОГО ЗАБРУДНЕННЯ ҐРУНТІВ ТА РОСЛИННОЇ ПРОДУКЦІЇ ЧЕРНІГІВСЬКОЇ ОБЛАСТІ Спостереження за щільністю забруднення ґрунту і рівнями забруднення рослинницької продукції радіонуклідами проводяться Чернігівським ...
оній - 240). Крім того, у викидах були присутні високорадіоактивні частки палива (гарячі частиці). Результати аеродозиметричного контролю радіаційного становища і відбора та аналіза навколишнього чередовища, які почали проводитися невдовзі після аварії, показали, що шайбільш забрудненим виявився район навколо реактора, який згодом став забороненою зоною. В інших районах зміни напрямку вітру і ...
... ії суші, хоча це й не така велика проблема, якщо врахувати, що 15% території Землі – пустелі. Геологічні проблеми на Україні Декілька слів скажемо про основні екологічні проблеми пов’язані з геологічною діяльністю на Україні. Територія України характеризується складними і різноманітними природними і інженерно - геологічними умовами. Багато районів відносяться до категорії техногенно навантажених ...
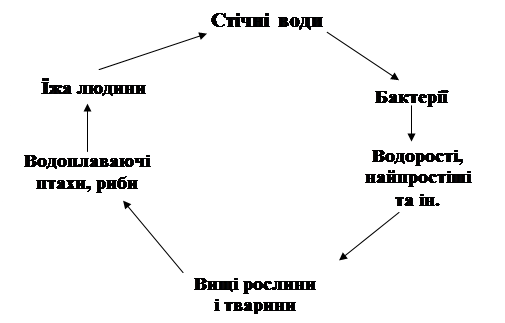
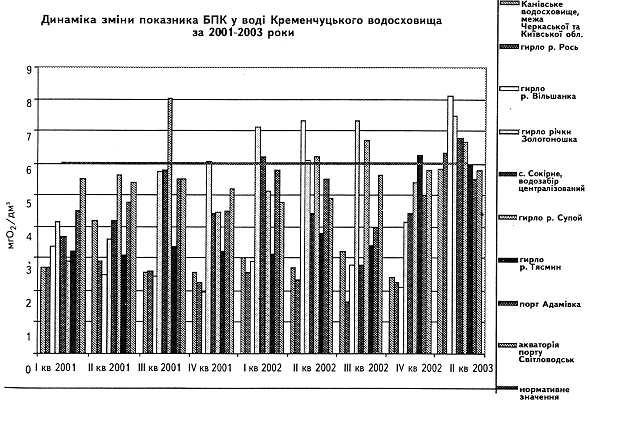
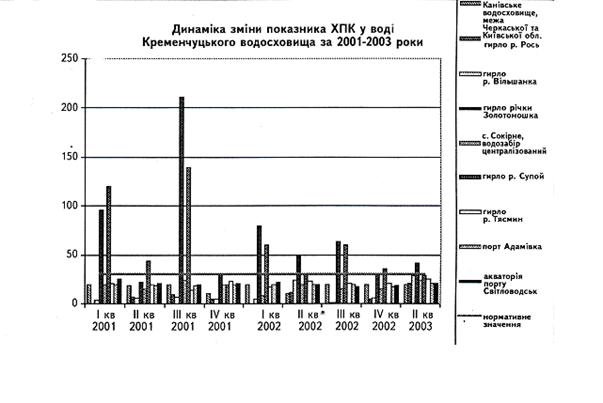
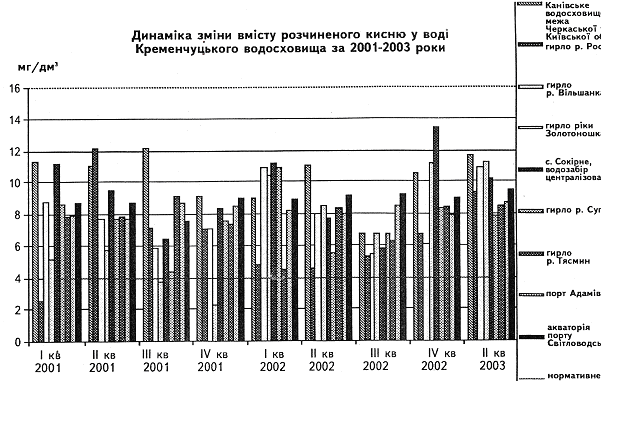
... у кістяку наземних видів тварин. Нині людство майже не вживає природну питну воду і перейшло на техногенну — її одержують на водопровідних станціях шляхом фізико-хімічної обробки [1]. 2 СТАН ГІДРОБІОЦЕНОЗУ КРЕМЕНЧУЦЬКОГО ВОДОСХОВИЩА 2.1 Гігієнічний моніторинг Кременчуцього водосховища 2.1.1 Створення моніторингу Кременчуцького водосховища в межах Черкаського регіону ...
0 комментариев