Навигация
Поперечносмугаста м’язова тканина
23009
знаков
0
таблиц
1
изображение
2. Поперечносмугаста м’язова тканина
Скелетна (соматична) м'язова тканина — це робоча мускулатура апарату руху, глотки, гортані, верхньої половини стравоходу, язика, жуйних м'язів. Джерелом розвитку цієї тканини є матеріал міотомів сегментованої мезодерми, а поперечносмугаста тканина внутрішніх органів розвивається із спланхнотому. На ранній стадії розвитку міотоми складаються із щільно розміщених м'язових клітин — міобластів, цю стадію гістогенезу називають міобластичною.
Міобласти диференціюються у двох напрямах, одні здатні зливатися і утворювати симпласти — м'язові трубочки, які далі формують дефінітивні структури – міосимпласти. У результаті другої лінії диференціації розвиваються клітинні структури - міосателітоцити.
На міобластичній стадії гістогенезу цитоплазма міобластів має тонковолокнисту будову, що характеризує розвиток скорочувальних білків. Уже на цій стадії міобласти здатні до скорочення. Ядра їх великі, овальні з невеликим вмістом гетерохроматину. Вони здатні до інтенсивнішого поділу, ніж сама клітина, в результаті чого міобласти стають багатоядерними, витягуються у довжину і набувають форму волокна — симпласта. Міосимпласти можуть утворюватися і шляхом злиття міобластів — це друга стадія гістогенезу — м'язових трубочок. Останні здатні розщеплюватися вздовж і формувати м'язові волокна, в яких різко збільшується кількість міофібрил. Численні ядра зміщуються на периферію під плазмолему, волокна набувають посмугованості. Це третя стадія гістогенезу — м'язових волокон. До м'язових волокон підростає сполучна тканина з кровоносними судинами, нервами, які диференціюються в нервові закінчення.
Наведені дані про гістогенез допоможуть зрозуміти будову соматичної м'язової тканини та ті складні зміни, які у ній відбуваються під час тренування, фізичних навантажень та при фізіологічній регенерації і патології.
Як видно з гістогенезу, у диференційованої скелетної м'язової тканини відсутня клітинна будова. Основною структурною і функціональною одиницею її є м'язове волокно. Довжина його часто збігається з довжиною м'яза, до складу якого воно входить. Волокно покрите сарколемою (від грецьк. «саркос» — м'ясо), яка складається із двох мембран. Зовнішня базальна мембрана стикується із ретикулярними та тонкими колагеновими волокнами сполучної тканини, що її оточує. Внутрішня мембрана аналогічна плазмолемі усіх тканинних клітин і бере участь у проведенні імпульсів скорочення волокна. Плазмолема утворює систему вузеньких канальців, які пронизують все волокно і зливаються із сарколемою протилежного боку. Таким чином, вся саркоплазма пронизана системою поперечних трубок, Т-системою. Цитоплазма має спеціальну назву — саркоплазма, структурними компонентами якої є міофібрили (скорочувальний апарат волокна), органели, включення, гіалоплазма. Численні ядра м'язового волокна розміщуються на периферії під саркоплазмою. У гіалоплазмі волокна локалізуються добре розвинені мітохондрії (саркосоми), які скупчуються між мікрофібрилами, навколо численних ядер, безпосередньо, під сарколемою. Це ті зони волокна, які потребують значної кількості АТФ, через це стає зрозумілою висока метаболічна активність скелетних м'язів. Найбільш інтенсивного розвитку досягає агранулярна ендоплазматична сітка (саркоплазматичний ретикулум). Її мембранні елементи розмішуються вподовж саркомерів (частинки, з яких складається волокно й залежить його довжина). Саркоплазматичному ретикулуму притаманна специфічна функція нагромадження іонів кальцію, необхідних при скороченні та розслабленні м'язового волокна. Інші органоїди (гранулярний ендоплазматичний ретикулум, комплекс Гольджі та інш.) розвинені слабше і локалізуються біля ядер, розміщених на периферії під сарколемою. Ядра можуть розміщуватися у вигляді ланцюжка один за одним, що є в результаті амітотичного поділу — показник реактивного стану м'язового волокна. Кількість ядер коливається від десятка до декількох сотень, вони мають овальну форму. Гетерохроматин у вигляді великих гранул знаходиться у порівняно світлій нуклеоплазмі.
Поміж міофібрил локалізується значна кількість гранул глікогену (трофічного) включення — матеріалу для синтезу АТФ. У цитоплазмі м'язового волокна містяться дихальні ферменти, білок, міоглобін — аналог гемоглобіну еритроцитів, який також здатний зв'язувати та віддавати кисень. Міоглобін забарвлює м'язові волокна у червоний колір. Залежно від вмісту саркоплазми (а, отже, і міоглобіну), товщини і ферментного складу м'язові волокна поділяють на червоні та білі. Червоні волокна у саркоплазмі містять велику кількість міоглобіну, численні мітохондрії, багаті на цитохроми, міофібрили у них мають незначну товщину. М'язи, у яких переважають червоні волокна, здатні до тривалішої безперервної активності, оскільки їх саркоплазма добре забезпечує енергетичні потреби.
Білі волокна містять у саркоплазмі менше міоглобіну та мітохондрій, у них більша товщина міофібрил. Білі м'язові волокна здатні скорочуватися швидше, ніж червоні, але вони порівняно швидко втомлюються, тому що не забезпечують структури саркоплазми достатньою кількістю енергії.
Найбільшу питому вагу у саркоплазмі становлять міофібрили. Вони розміщуються вздовж м'язового волокна і їх довжина збігається з довжиною волокна, діаметр міофібрил — 1—2 мкм. Вони мають характерну поперечну смугастість (чергування світлих і темних смуг, дисків), що зумовлено особливістю їхньої структури і, в зв'язку з цим, різними оптичними властивостями. Усі темні й всі світлі диски міофібрил в одному волокні містяться на одному рівні, через що волокно має поперечну посмугованість. Повздовжня орієнтація міофібрил надає м'язовому волокну поздовжню смугастість.
У поляризованому світлі темні смуги (диски) мають подвійне променезало-млення — анізотропію, через що їх називають анізотропними (дисками) А. Світлі диски є однопроменезаломлюючими, через що їх називають ізотропними — І. Всередині кожного І-диска проходить темна зона — лінія Z (телофрагма). У центрі А-диска спостерігається світліша зона-лінія Н, або смужка Гензена, посередині якої розміщується тонка, темна лінія М (мезофрагма). Міофібрили можна одержати, розщепивши м'язове волокно.
Структурною одиницею міофібрил є саркомер. Це ділянка міофібрили між двома телофрагмами. В міофібрилі саркомери розміщуються один за одним і в зоні Т-телофрагми, багатої на глікозаміноглікани, міофібрили можуть при мацерації розпадатися на окремі саркомери «саркос»-м'ясо, «мерос»-частина.
Саркомер — це частина міофібрили, що складається із Т-телофрагми — лінії Z (для двох сусідніх саркомерів), половин І-диска та А-диска половини зони Н, мезофрагми - М, половини зони Н, половин А- і І-дисків, лінії Z- телофрагми.
Саркомери — це елементарні скоротливі одиниці поперечно-смугастих м'язів, які скорочуються завдяки здатності зменшувати свою довжину в два рази. Електронномікроскопічними, гістохімічними, біохімічними дослідженнями вдалося встановити функціональну морфологію саркомера. Були ідентифіковані поздовжні нитки саркомера-міофіламенти або мікрофіламенти двох типів. А-диск складається із товстих міофіламентів (діаметр — 10— 12 нм, довжина — 1,5—2 мкм), І-диск — із тонких (діаметр — 5-7 нм, довжина — 1-1,3 мкм) міофіламентів. Матеріалом, з якого складаються товсті міофіламенти, є білок міозін, а тонкі — актин, тропоміозин-В і тропін. Кількісне відношення міозинових і актинових міофіламентів в одній міофібрилі 1:2, тобто на один міозиновий міофіламент припадає два актинових. Актинові й міозинові міофіламенти контактують один з одним не кінець в кінець, а переміщуються (ковзають) по відношенню один до одного і в А-диску утворюють зону перекриття. Частину А-диска, яка складається лише з міозинових міофіламентів, називають Н-лінією і, порівняно із зоною перекриття, вони світліші. При скороченні саркомера актинові міофіламенти ще далі проникають у проміжки між міозиновими, а при повному скороченні їхні вільні кінці майже збігаються у середині саркомера.
Оскільки довжина таких філаментів залишається незмінною, вони, просочуючись між товстими філаментами, тягнуть за собою телофрагми, до яких прикріплені, тим самим зближуючи кінці усіх саркомерів. У повністю скороченому саркомері Н-зона, а також І-диски майже зникають і весь саркомер перетворюється на зону перекриття. М-лінія (мезофрагма) — це місце з'єднання товстих міозинових міофіламентів в анізотропному А-диску, Z-лінія (телофрагма) проходить через усю товщину саркомера, а зона прикріплення тонких актинових міофіламентів має зигзагоподібний контур. Z-лінія складається із Z-філаментів, до складу яких входять білки — тропоміозин-В та L-актин, Z-філаменти формують решітку, до якої з обох боків прикріплюються тонкі актинові міофіламенти І-дисків двох сусідніх саркомерів. Таким чином, телофрагми (Z-лінії) та мезофрагми (М-лінії) є опорним апаратом саркомерів. Отже, під час скорочення м'язового волокна його скоротливий апарат зазнає таких змін: зменшується довжина саркомера, оскільки актинові міофіламенти І-диску просуваються (ковзають) між міозиновими А-диску, зсуваючись до Н-лінії (мезофрагми) А-диску; збільшуються зони перекриття, формуючи бокові з'єднання (місточки) між актиновими та міозиновими міофіламентами; скорочується Н-лінія (мезофрагма); зближуються Z-лінії (телофрагми). Чим сильніше скорочується міофібрила, тим глибше актинові міофіламенти заходять у проміжки між міозиновими, зони перекриття розширяються за рахунок звуження Н-ліній.
Для розуміння механізму скорочення міофібрили необхідно згадати про наявність спеціалізованого саркоплазматичного ретикулуму саркоплазми волокна та утворення плазмолемою системи поперечних канальців Т-трубочок. Зони контакту системи Т-трубочок і термінальних цистерн саркоплазматичного ретикулуму називають тріадами. Т-трубки проходять на рівні Z-ліній (телофрагми) через все волокно і контактують з цистернами саркоплазматичної сітки з протилежного боку. Таким чином, Т-трубочки локалізуються на межі двох саркомерів і контактують з терміналями саркоплазматичної сітки обох саркомерів, утворюючи тріади. Ці структури відіграють основну роль в деполяризації (поширенні імпульсу) та акумуляції іонів кальцію.
Плазмолема м'язового волокна, так як і неврилема нервових волокон, електрично поляризована. Внутрішня поверхня плазмолеми, розслабленого м'язового волокна, має негативний потенціал, а зовнішня-позитивний. При скороченні м'язових волокон хвиля деполяризації по нервовому волокну через нервове закінчення досягає плазмолеми м'язового волокна і викликає її місцеву деполяризацію. Через систему Т-трубочок, яка зв'язана з плазмолемою та тріадою, хвиля деполяризації впливає на проникність мембран саркоплазматичного ретикулуму, що призводить до звільнення акумульованих іонів кальцію з її поверхні у саркоплазму. В присутності іонів кальцію активізується розщеплення АТФ, що необхідно для утворення актоміозинового комплексу та ковзання актинових міофіламентів по відношенню до міозинових. Це викликає скорочення кожного саркомера, а звідси міофібрил та м'язових волокон у цілому (рис. 1).
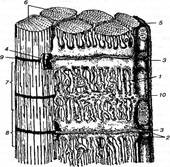
Рис.1. Схема будови відрізка м'язового волокна:
1—саркоплазматичний ретикулум; 2—термінальні цистерни саркоплазматичного ретикулуму; 3 — Т-трубки; 4 — тріади; 5 —сарколема; 6 —міофібрили; 7—А-диск; 8— І-диск; 9 — Z-лінія; 10 — мітохондрії.
Важливе місце у цьому процесі належить молекулам міофіламентів-міозину. Вони складаються із головки та довгого хвостика. При гідролізі АТФ, чому сприяє АТФ-на активність головок молекул міозину, вони утворюють зв'язки (містки) з певними частинами молекул міофіламентів-актину. Актинові міофіламенти зближаються до центру саркомеру, Z-лінії (телофрагми) зближуються, збільшуються зони перекриття, зменшуються Н-зони (мезофрагма) анізотропних дисків міофібрил. Потім з участю АТФ актоміозинові зв'язки руйнуються, а міозинові головки приєднуються до сусідніх ділянок актинових міофіламентів, що сприяє дальшому просуванню міофіламентів один до одного. При зменшенні концентрації іонів кальцію (вони трансформуються на мембрани саркоплазматичного ретикулуму) скорочення м'язового волокна припиняється. Для цього також необхідна енергія АТФ. Отже, при розслабленні так, як і при скороченні м'язового волокна витрачається АТФ, джерелом якої в саркоплазмі є глікоген, глюкоза та жирні кислоти.
Кінці м'язових волокон міцно фіксуються до сухожилків або сухожильних прошарків, що розміщуються між ними. Сарколема утворює пальцеподібні вирости, між якими знаходяться колагенові волокна сполучної тканини, які кріплять м'язові волокна до кісток. Цей зв'язок настільки міцний, що при навантаженні, яке здатне розірвати м'язи або сухожилки, структура залишається цілою. Тонкі прошарки рихлої сполучної тканини між м'язовими волокнами називаються ендомізієм, ретикулярні і колагенові волокна його переплітаються з волокнами сарколеми (зовнішній сполучно-тканинний шар). В ендомізії локалізуються гемокапіляри та структури нервової тканини. Комплекс волокна з оточуючими його елементами є структурною і функціональною одиницею скелетного м'яза. М'язові волокна об'єднуються у пучки, між якими наявні товстіщі прошарки рихлої сполучної тканини, яка носить назву перимізій. Сполучна тканина, що покриває м'яз у цілому, як орган, називається епімізієм.
Поперечносмугаста м'язова тканина здатна до активної регенерації. Репа-ративна регенерація відбувається на фоні відмирання старих структур і утворення нових. Як і при нормальному гістогенезі, регенерація відбувається у три фази: міобластичну; м'язових трубочок; формування м'язового волокна.
3. Серцева м’язова тканина (міокард)
Серцева м'язова тканина складається із м'язових клітин-кардіоміоцитів, які, з'єднуючись своїми кінцями по довгій осі клітини формують структури, подібні до м'язових волокон. Поперечна смугастість має ту ж природу, що і в скелетних м'язах, тобто зумовлена оптичною неоднорідністю міофібрил, що складаються із двох типів міофіламентів. Останні містять анізотропні А-диски та ізотропні І-диски, що займають периферійну частину клітини. Кардіоміо-цити формують м'язові волокна, які анастомозують між собою, утворюючи симпласт. Різниця в будові та функції кардіоміоцитів дає підставу класифікувати їх на два види: скоротливі, або типові серцеві міоцити (становлять більшу частину серцевого м'яза) — це робоча мускулатура та провідні або атипові серцеві міоцити, що формують провідну систему серця.
Типові кардіоміцити — це одно або двоядерні клітини, світлі ядра яких локалізуються у центрі, на відміну від крайового розміщення у скелетних м'язових волокнах. На поверхні міоцити мають відростки, або анастомози, за допомогою яких вони з'єднуються. Саркоплазма містить міофібрили, органели, включення, гіалоплазму. Добре розвинені мітохондрії (саркосоми), гірше-комплекс Гольджі та саркоплазматична сітка, яка не утворює великих термінальних цистерн (як у скелетних м'язах). Із включень найбільше міститься гранул глікогену та пігменту ліпофусцину, кількість якого з віком збільшується.
Місце з'єднання міоцитів - це вставні диски-аналоги Z-пластинок (тело-фрагми). Вставні диски під електронним мікроскопом мають хвилястий вигляд і два види сполучення: це десмосоподібні та щільні контакти. Десмосоподібні контакти забезпечують міцне з'єднання клітин, а щільні-електричний зв'язок сусідніх кардіоміоцитів та проведення імпульсів по м'язовому пласту міокарда. У кардіоміоцитах Т-система (трубочки) заходять всередину клітини на рівні Z-пластинок (телофрагм), тому їх кількість відповідає кількості саркомерів. Т-трубочки набагато ширші, ніж у скелетних м'язах і, крім того, ще вистелені базальною мембраною, що лежить зовні від сарколеми. Характерною особливістю серцевих міоцитів є відсутність тріад, оскільки цистерни саркоплазматичного ретикулуму разом із Т-трубочками не утворюють нових кілець навколо міофібрил. Функція Т-трубочок кардіоміоцитів до проведення імпульсів у клітину і забезпечення одночасного скорочення усіх міофібрил. Оскільки у типових серцевих міоцитів відсутні тріади, для їх скорочення необхідний приплив іонів кальцію з інших тканин. Ось чому виведення в кров солей кальцію підсилює роботу серцевого м'яза. Кардіоміоцити, що розміщуються у передсерді, мають здатність синтезувати передсердний натрійуретичний фактор (ПНФ), який діє діуретично (підсилює виведення з організму води і солей та знижує артеріальний тиск). Передсердний натрійуретичний фактор — це поліпептидний гормон, що є модулятором або антагоністом системи ренин-ангіотензин-альдостерон (гормони нирок і надниркових залоз).
Атипові, провідні кардіоміоцити формують провідну систему серця. Серед провідних, атипових серцевих міоцитів за морфологічними та функціональними особливостями виділяють три типи клітин.
Пейсмейкерні клітини (Р-клітини) — це водії ритму серця. Вони мають нестабільний біопотенціал спокою і здатні у спокої деполяризуватися з частотою 70 разів за 1 хв, тобто ці клітини генерують імпульси до скорочення.
Перехідні клітини — передають збудження від Р-клітин до наступних клітин пучка і скоротливих елементів міокарда. Клітини провідного пучка та його ніжок (волокна Пуркіньє). Вони передають збудження від перехідних клітин до скоротливих серцевих міоцитів шлуночків.
Ця різновидність серцевої м'язової тканини формує систему, яка забезпечує проведення збудження серцевого ритму.
Спеціалізовані скоротливі тканини складаються із клітин, що входять до складу стінки потових, слинних та молочних залоз і розвиваються із епітеліальних клітин. Клітини, які звужують зіницю і розміщуються в райдужній оболонці, також здатні до скорочення і мають нейрогліальне походження.
Література
1. Гистология, цитология и эмбриология / Под ред. О.В.Волкова, Ю.К. Елецкого. - М: Медицина, 1996.
2. Новак В.П., Пилипенко М.Ю., Бичков Ю.П. Цитологія, гістологія, ембріологія.: Підручник. – К.: ВІРА-Р, 2001. – 288 с.
Похожие работы
... суглоба, прискорюється регенерація кісткової тканини, розсмоктування внутрішньосуглобового випоту і патологічних відкладень у періартрикулярних тканинах. Безпосередньо на жирову тканину масаж майже не діє. Лише при тривалому його застосуванні спостерігається незначне зменшення жирових відкладень, що можна пояснити загальним впливом масажу на обмін речовин. Підвищуючи його в організмі, посилюючи ...

... віком вона продовжує формуватися і набуває індивідуальні особливості. Ці особливості визначаються багатьма факторами: ростом, вагою, пропорціями тулуба і кінцівок, наявністю вроджених порушень опорно-рухового апарату, особливостями обміну речовин. На осанку негативно сприяє і неповноцінне харчування, і загальний незадовільний стан здоров’я, і забруднення навколишнього середовища, і гострі та хрон ...
... беременности у крыс / М. А. Волошин, О. Г. Кущ // Актуальные вопросы эволюционной, возрастной и экологической морфологии. – Белгород, 2006. – С. 34. (Самостійно проведено дослідження, обробка результатів, написання тез). АНОТАЦІЯ Кущ О.Г. Закономірності будови плаценти і лімфоїдної тканини, асоційованої з нею, протягом третього періоду вагітності (анатомо-експериментальне дослідження). – Рук
... ів, тканин внутрішнього середовища, які виконують захисну функцію. Це клітини з двома, трьома відростками, які мають вторинні розгалуження, ядра їх багаті на гетерохроматин. При подразненні нервової тканини (запалення, рана) клітини мікроглії збільшуються в об'ємі, стають рухомими, наповнюються фагоцитованим матеріалом. Мікрогліоцити здатні до синтезу білків-імуноглобулінів, вони характерні для ...
0 комментариев